







研究论文
● 期刊:Nature (IF:50.5)
● DOI:https://doi.org/10.1038/s41586-024-08440-7
● 原文链接:
https://www.nature.com/articles/s41586-024-08440-7
● 第一作者:Emmanuel Stephen-Victor
● 通讯作者:Seth Rakoff-Nahoum(seth.rakoff-nahoum@childrens.harvard.edu),Talal A. Chatila (talal.chatila@childrens.harvard.edu)
● 发表日期:2025-1-22
● 主要单位:
美国马萨诸塞州波士顿儿童医院、美国马萨诸塞州波士顿哈佛医学院、美国马萨诸塞州哈佛儿童医院、美国加利福尼亚州斯坦福大学、美国马萨诸塞州波森堡哈佛大学陈曾熙公共卫生学院环境卫生系、美国马萨诸塞州剑桥市哈佛干细胞研究所、美国佛罗里达州迈阿密市杰克逊卫生系统迈阿密大学米勒医学院和霍尔茨儿童医院儿科新生儿科、美国马萨诸塞州坎布里奇市麻省理工学院和哈佛大学博德研究所
摘要Abstract
对膳食抗原的免疫耐受是避免引发食物过敏(FA)和过敏反应的有害2型免疫应答的关键。然而,导致食物抗原耐受性维持与失效的机制尚不明确。本研究发现,杯状细胞来源(goblet-cell-derived)的抗性素样分子β(resistin-like molecule β, RELMβ)是口服耐受的核心调控因子。RELMβ在FA患者和FA小鼠模型的血清中均显著富集。基因敲除RELMβ可保护小鼠免受FA及食物抗原特异性IgE和过敏反应的发生。RELMβ通过调控肠道微生物组,耗竭产吲哚代谢物的乳杆菌属(Lactobacilli)和另枝菌属(Alistipes)菌株,破坏机体对食物的耐受性。这种耐受性由肠道局部产生的吲哚衍生物维持,这些代谢物通过激活芳香烃受体(aryl hydrocarbon receptor),驱动具有FA保护作用的RORγt⁺调节性T(Treg)细胞。在断奶关键期进行RELMβ拮抗干预可恢复口服耐受性,并对遗传易感后代在生命后期发生FA具有保护作用。本研究揭示了RELMβ通过调控"肠道免疫-上皮轴"来调节食物抗原耐受性的新机制,即通过微生物组编辑实现适应性免疫的先天性调控新模式,并确定了该通路中可用于FA预防和治疗的潜在靶点。
正文main
2型免疫在介导针对寄生虫和毒素的保护性组织反应中具有关键作用。其失调会导致病理性反应,包括食物过敏(FA)和过敏反应。值得注意的是,以IL-4和IL-13为代表的2型免疫细胞因子可诱导包括RELMβ在内的多种肠道上皮源性蛋白表达。通过使用因IL-4受体功能获得性突变(Il4raF709)导致的2型免疫异常小鼠模型,我们发现相较于野生型(WT)小鼠,Il4raF709小鼠肠道中Retnlb(编码RELMβ)转录本表达显著升高(图1a)。通过口服致敏食物抗原卵清蛋白(OVA)与黏膜佐剂金黄色葡萄球菌肠毒素B(SEB)构建的OVA-SEB模型显示,Il4raF709小鼠可产生强烈的食物过敏和过敏反应。在OVA-SEB诱导的FA模型中,与基线状态或野生型小鼠相比,RELMβ的肠道表达水平(图1a)和全身蛋白水平(图1b)均显著增加,而其同源蛋白RELMα在野生型或Il4raF709小鼠中,无论是基线状态还是FA诱导后均未出现显著差异(图1a,b)。
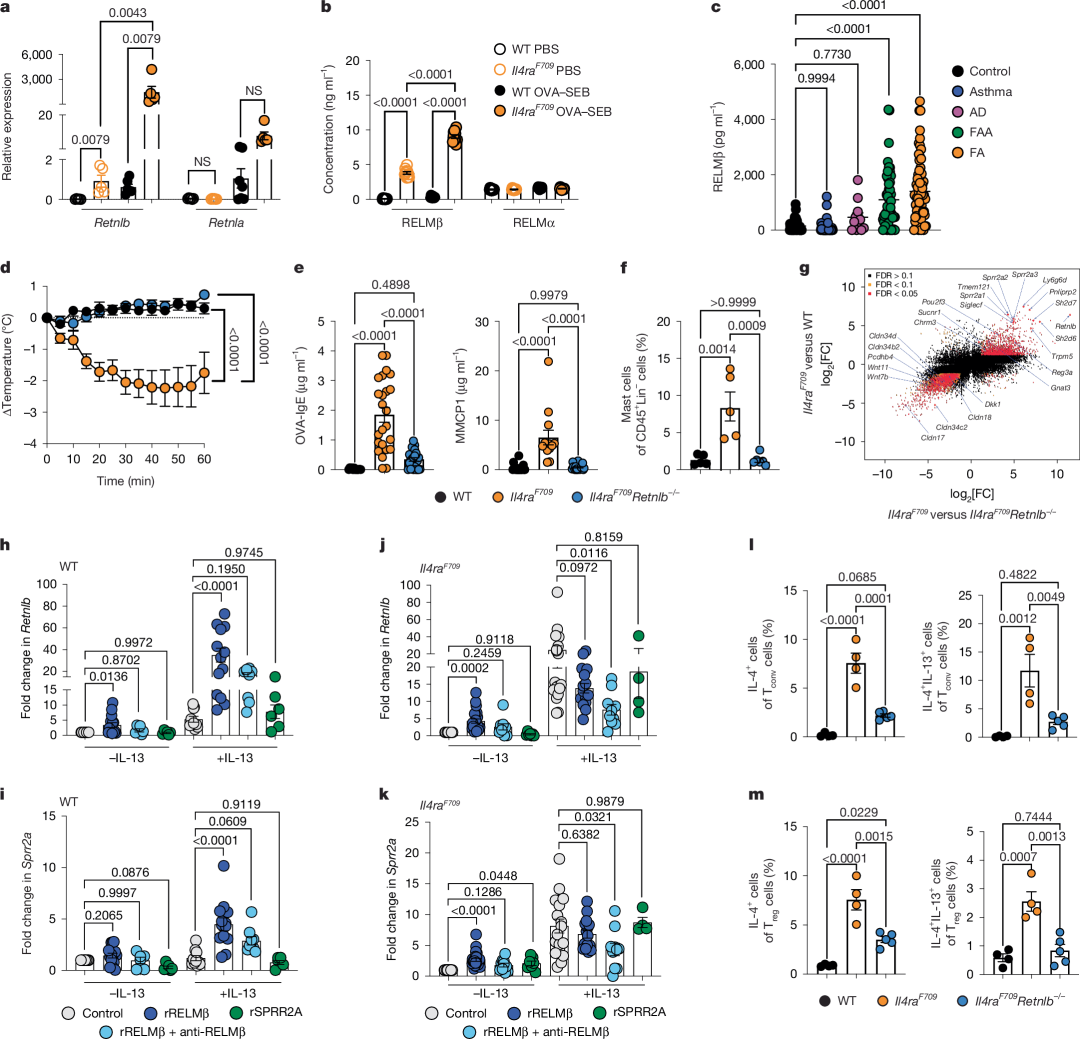
图1 | RELMβ在FA诱导中的关键作用
a, 空肠组织Retnlb和Retnla转录本的定量PCR(qPCR)分析(相对于基线的相对表达量),样本包括假致敏(PBS)的WT和FA易感Il4raF709小鼠(从左到右n = 5、5、6、6)以及OVA–SEB致敏后经OVA激发的小鼠(n = 6、5、7、5)。
b, 通过酶联免疫吸附试验(ELISA)测定PBS处理或OVA–SEB致敏的WT(n = 10)和Il4raF709(n = 10)小鼠血清中RELMβ水平。
c, 健康对照个体(n = 44)、哮喘患者(n = 34)、特应性皮炎(AD)患者(n = 12)、FA合并哮喘(FAA)患者(n = 52)及单纯FA患者(n = 67)血清中RELMβ水平。
d, OVA–SEB致敏并经OVA激发的WT、Il4raF709及Il4raF709Retnlb−/−小鼠的核心体温变化(n = 7、12、11)。
e, 激发后血清MMCP1浓度(n = 19、13、19)。
f, 激发后空肠肥大细胞频率(n = 5、5、6)。
g, 空肠上皮细胞RNA测序(RNA-seq)分析的log–log图,比较Il4raF709与Il4raF709Retnlb−/−(x轴)及Il4raF709与WT小鼠(y轴)。每组n = 3。
h-k, 来自WT(h,i)和Il4raF709(j,k)小鼠的肠道类器官中Retnlb(h,j)和Sprr2a(i,k)转录本的qPCR分析。类器官分别接受假处理(培养基)、重组RELMβ(rRELMβ)、重组SPRR2A(rSPRR2A)或rRELMβ加抗RELMβ单克隆抗体处理(未加IL-13:WT组n = 12、15、6、6;Il4raF709组n = 15、23、9、10、7;加IL-13:WT组n = 10、13、9、6;Il4raF709组n = 18、15、10、4)。
l, m, 激发后血清MMCP1浓度(n = 19、13、19)。流式细胞术分析WT(n = 4)、Il4raF709(n = 4)及Il4raF709Retnlb−/−(n = 5)小鼠空肠固有层淋巴细胞(SI-LPLs)中IL-4⁺和IL-13⁺ CD4⁺ Tconv(l)或Treg(m)细胞群的频率。
(a–m数据均以均值±标准误表示。统计分析采用单因素方差分析(ANOVA;a–c、e、f、h–m)配合Šídák(a、b)、Tukey(c、e、f、l、m)或Dunnett(h–k)事后检验,或重复测量双因素方差分析配合Tukey事后检验(d)。)
在确定RELMβ在小鼠FA模型中存在局部和系统性异常表达后,我们分析了FA患儿血清中的RELMβ浓度,比较对象包括未并发哮喘的FA患儿、并发哮喘的FA患儿、单纯哮喘患儿、单纯特应性皮炎(AD)患儿以及无过敏或特应性体质的健康儿童。与单纯哮喘、AD或非过敏对照组相比,无论是否并发哮喘,多数FA受试者的RELMβ水平均显著升高(图1c和补充表1)
为探究RELMβ在FA发生中的机制作用,我们在Il4raF709小鼠中敲除Retnlb基因。RELMβ缺失完全消除了口服激发后的过敏反应(图1d),抑制了血清OVA特异性IgE和黏膜肥大细胞蛋白酶1(MMCP1)水平(图1e),并阻断了小肠组织肥大细胞扩增(图1f)。此外,RELMβ调控小肠上皮细胞中更广泛的2型免疫相关程序,包括依赖RELMβ的杯状细胞相关抗菌基因转录本(如Sprr2a1/2/3和Reg3a11)增加、簇细胞相关基因转录本(Trpm5、Pou2f3、Sucnr1和Gnat3)上调,以及多个紧密连接相关claudin蛋白编码转录本表达下调(图1g和扩展数据图1a,b)。体内给予重组RELMβ处理可诱导初始Il4raF709小鼠表达Sprr2a、Reg3a和Reg3b(扩展数据图1c)。为明确RELMβ对肠上皮2型免疫程序的直接作用,我们用重组RELMβ处理WT或Il4raF709来源的小鼠肠上皮类器官。RELMβ通过与IL-13和IL-4受体功能获得性信号(Il4raF709)协同作用,调控肠上皮细胞Retnlb和Sprr2a表达(图1h-k和扩展数据图1d,e)及RELMβ蛋白水平(扩展数据图1f-i)。该协同效应依赖内源性RELMβ,因为抗RELMβ抗体处理可抑制该效应,而Sprr2a的诱导在Retnlb敲除后消失(图1h-k和扩展数据图1d-i)。相比之下,RELMβ调控的SPRR2A处理既不诱导Retnlb也不诱导Sprr2a表达(图1h-k和扩展数据图1d,e)。在人肠道类器官模型中,RELMβ单独或联合IL-13均可诱导RETNLB和SPRR2A转录本(扩展数据图1j,k)。这些发现表明RELMβ通过自我调节和IL-13依赖性反馈环路成为2型上皮程序的主调控因子。
为探究RELMβ是否近端调控2型肠道T细胞免疫应答,我们检测了小肠固有层淋巴细胞(SI-LPLs)中表达IL-4和IL-13的CD4⁺ T细胞亚群频率。结果显示,与Il4raF709小鼠相比,Il4raF709Retnlb−/−小鼠LPL中常规T细胞(Tconv)和调节性T细胞(Treg)的IL-4⁺及IL-13⁺细胞群均减少(图1l,m和扩展数据图1l,m)。RELMβ缺失还抑制了肠系膜淋巴结(MLNs)中IL-4⁺细胞、促过敏反应的IL-4/IL-13共表达滤泡辅助性T细胞13(TFH13)频率,以及高亲和力促过敏抗OVA IgE抗体产生(扩展数据图2a-f)。这些结果证实RELMβ协调病理性局部和系统性2型效应免疫应答。
在明确RELMβ对Il4raF709小鼠FA发生的关键作用后,我们进一步采用模拟湿疹相关FA的WT BALB/c小鼠经皮致敏模型,以基因型和佐剂非依赖的方式评估RELMβ在FA中的普遍作用。在该模型中,与WT小鼠相比,RELMβ缺陷小鼠经皮OVA致敏后口服激发引发的过敏反应被消除(扩展数据图3a-d)。这些发现证明RELMβ在不同遗传背景和致敏途径的小鼠FA模型中具有普适性作用。
RELMβ抑制FA保护性RORγt⁺ Treg细胞
RORγt⁺ Treg细胞在免疫调节(包括维持食物过敏原耐受)中起核心作用。与WT小鼠相比,Il4raF709小鼠SI-LPLs中RORγt⁺ Treg细胞减少,这是其口服耐受破坏的基础[23]。Retnlb敲除可逆转RORγt⁺ Treg细胞耗竭(图2a,b)。重要的是,在Il4raF709Retnlb−/−小鼠Treg细胞中敲除编码RORγt的Rorc基因,可消除RELMβ缺失对局部和系统性FA的保护作用(图2c-f)。这些结果表明RELMβ通过抑制耐受性和保护性RORγt⁺ Treg细胞,成为FA中2型介质和效应应答的核心调控因子。
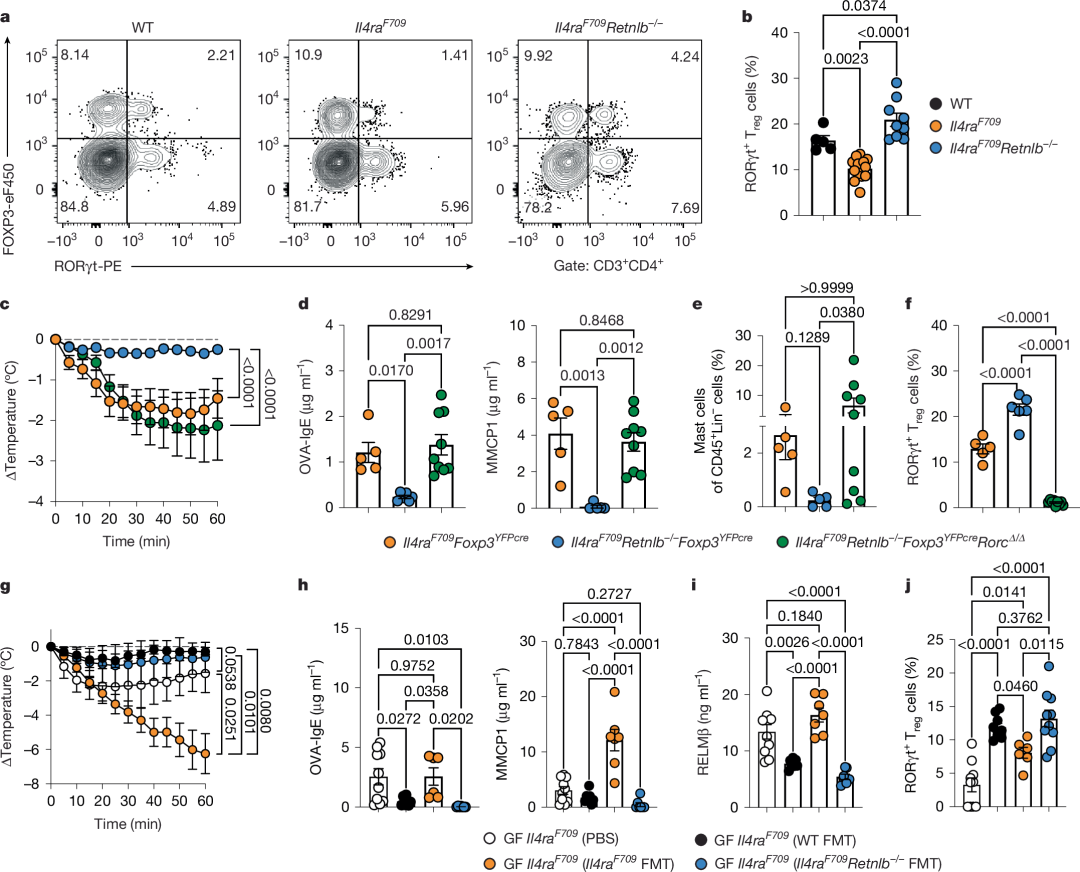
图2 | RELMβ抑制FA保护性RORγt⁺ Treg细胞
a,b, OVA–SEB致敏的WT、Il4raF709及Il4raF709Retnlb−/−小鼠小肠固有层淋巴细胞(SI-LPLs)中RORγt⁺ Treg细胞的流式细胞分析(a),以及各品系小鼠SI-LPL CD4⁺FOXP3⁺RORγt⁺ Treg细胞频率(n = 5、14、9)(b)。
c, OVA–SEB致敏并经OVA激发的Il4raF709Foxp3YFPcre(n = 18)、Il4raF709Retnlb−/−Foxp3YFPcre(n = 17)及Il4raF709Retnlb−/−Foxp3YFPcreRorc∆/∆(n = 13)小鼠的核心体温变化。
d, OVA特异性血清IgE浓度(n = 5、6、9)及血清MMCP1浓度(n = 5、5、9)。
e, 空肠肥大细胞频率(n = 5、5、9)。
f, SI-LPL RORγt⁺ Treg细胞频率(n = 5、6、9)。
g, 无菌(GF)Il4raF709小鼠接受假处理(PBS)(n = 10)或WT小鼠(n = 4)、Il4raF709小鼠(n = 7)、Il4raF709Retnlb−/−小鼠(n = 7)粪便微生物移植(FMT)后,经OVA–SEB致敏及OVA激发的核心体温变化。
h,i, 血清OVA特异性IgE浓度(n = 10、7、6、7)及血清MMCP1浓度(n = 10、7、7、7)(h),以及血清RELMβ浓度(n = 10、7、7、7)(i)。
j, 各品系小鼠SI-LPL RORγt⁺ Treg细胞频率(n = 10、9、6、10)。
(b–j数据均以均值±标准误表示。统计分析采用单因素方差分析(ANOVA)配合Tukey事后检验(b、d–f、h–j),或重复测量双因素方差分析配合Tukey事后检验(c、g)。)
RELMβ可在多个器官和组织的多种细胞类型中表达,包括肠道上皮(尤其是杯状细胞)。为明确肠道上皮源性RELMβ在驱动FA中的作用,我们将携带条件性Retnlb等位基因(Retnlbfl)的小鼠与在Il4raF709背景上表达villin启动子驱动Cre重组酶的小鼠交配,构建肠道上皮特异性RELMβ缺陷小鼠。经肠道上皮Retnlb转录本缺失证实,该模型在基线和致敏状态下均能抑制Il4raF709小鼠升高的血清RELMβ水平(扩展数据图4a,b)。肠道上皮单等位(Il4raF709VilcreRetnlb∆/+)或双等位(Il4raF709VilcreRetnlb∆/∆)Retnlb敲除均可保护小鼠免于过敏反应(扩展数据图4c),同时降低血清OVA特异性IgE和MMCP1水平,减少肠道上皮Sprr2a1和Reg3a转录本表达(扩展数据图4d,e)。重要的是,肠道上皮Retnlb敲除逆转了耐受性Helios⁻RORγt⁺ Treg细胞的抑制,并降低SI-LPLs和MLN中致病性Helios⁺GATA3⁺ Treg细胞频率(扩展数据图4f,g),表明肠道上皮细胞源性RELMβ是破坏食物抗原耐受所必需的。
我们进一步通过将Il4raF709背景的Atoh1(亦称Math1)启动子驱动Cre重组酶小鼠与Retnlbfl/fl小鼠交配,实现肠道分泌细胞特异性RELMβ基因敲除。肠道分泌谱系RELMβ缺陷小鼠受到保护免于过敏反应(扩展数据图4h),血清OVA特异性IgE和MMCP1水平降低(扩展数据图4i)。SI-LPL中耐受性Helios⁻RORγt⁺ Treg细胞频率增加,而致病性Helios⁺GATA3⁺ Treg细胞减少(扩展数据图4j,k)。OVA325–335特异性I-Ab四聚体染色显示,与Il4raF709Atoh1cre小鼠相比,致敏Il4raF709Atoh1creRetnlb∆/∆小鼠MLN和SI-LP中RORγt⁺ Treg细胞的OVA特异性显著富集。这些结果表明RELMβ缺失介导的保护作用由食物过敏原特异性RORγt⁺ Treg细胞赋予(扩展数据图5a-f)。综上证实肠道分泌细胞源性RELMβ是破坏食物抗原耐受的必要条件。
RELMβ缺失构建耐受性肠道微生物组
RELMβ以其抗菌功能著称。鉴于RELMβ由杯状细胞分泌至肠腔,且我们已证实肠道上皮分泌细胞源性RELMβ在驱动FA中的作用(扩展数据图4和5),我们推测RELMβ通过重塑肠道微生物组破坏食物抗原耐受。为验证此假说,我们将未致敏成年WT、Il4raF709及Il4raF709Retnlb−/−供体粪便微生物组移植至无菌Il4raF709小鼠。Il4raF709供体FMT未能保护无菌Il4raF709小鼠免于FA,而Il4raF709Retnlb−/−供体FMT则显示出保护作用(图2g)。与常规Il4raF709供体FMT相比,Il4raF709Retnlb−/−供体FMT可抑制OVA特异性IgE应答和肥大细胞脱颗粒(血清MMCP1),同时纠正RELMβ异常水平并增加RORγt⁺ SI-LPL Treg细胞(图2h-j)。这些结果表明RELMβ缺失提供的FA保护源于肠道微生物组在FA发生过程中的组成与功能差异。
我们进一步分析OVA致敏Il4raF709和Il4raF709Retnlb−/−小鼠(均为BALB/c背景)的肠道微生物组。与Il4raF709Retnlb−/−小鼠相比,Il4raF709小鼠Lactobacilli属和Alistipes属丰度降低(图3a),这与既往研究报道RELMβ在Muc2缺陷性结肠炎相关菌群失调中的作用一致。与RELMβ正常且FA易感的Il4raF709Retnlbfl/fl同窝或同笼小鼠相比,肠道上皮特异性RELMβ敲除(C57BL/6背景)可保护Il4raF709VilcreRetnlb∆/∆小鼠免于FA并恢复肠道Lactobacillus水平(图3b)。这种Lactobacilluss恢复现象在OVA变应原经皮致敏模型中同样被观察到(扩展数据图3e)。综上提示RELMβ通过重塑肠道微生物组构成促发食物过敏状态。
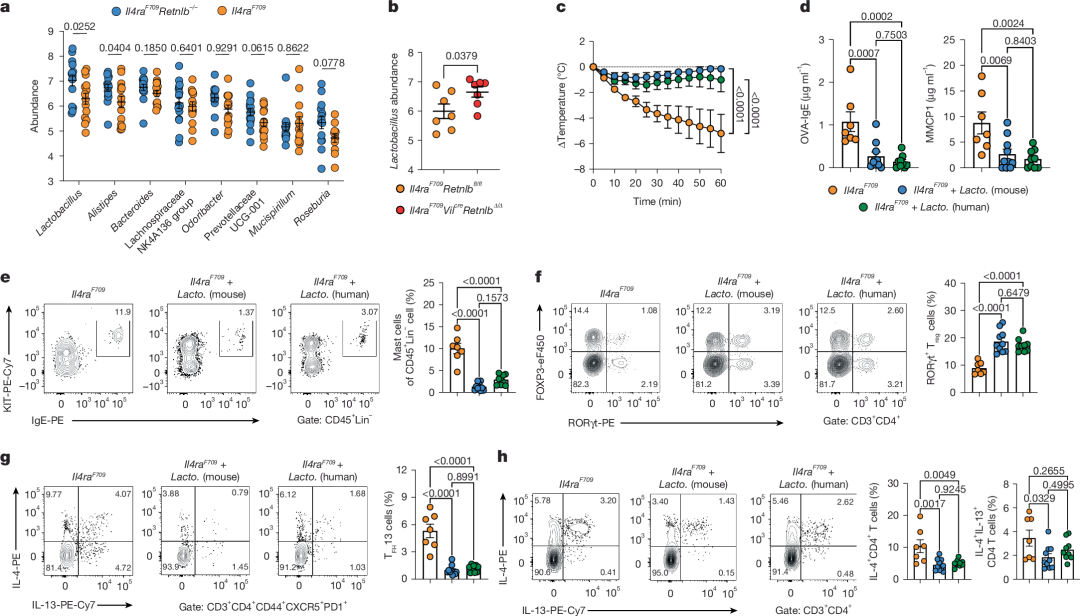
图3 | RELMβ缺失塑造耐受性肠道微生物组
a, OVA–SEB致敏的Il4raF709Retnlb−/−(n = 15)与Il4raF709(n = 14)小鼠(BALB/c背景)肠道中富集度最高的8个细菌属。
b, OVA–SEB致敏并激发的Il4raF709Retnlbfl/fl(n = 7)与Il4raF709VilcreRetnlb∆/∆(n = 7)小鼠(C57BL/6背景)中Lactobacillus属的丰度。
c, 通过给予Il4raF709Retnlb−/−小鼠富集的鼠源乳杆菌(L. reuteri和L. murinus)或人源乳杆菌(L. gasseri、L. reuteri和L. rhamnosus)进行菌群疗法,保护Il4raF709小鼠免于FA。小鼠每周接受OVA–SEB致敏(联合或不联合菌群疗法),随后经OVA口服激发(n = 7、10、9)。
d, 菌群疗法或假处理小鼠的OVA特异性血清IgE浓度及血清MMCP1浓度(n = 7、10、9)。Lacto.:Lactobacillus。
e,f, 流式细胞分析及小鼠(同c)空肠KIT⁺IgE⁺肥大细胞频率(n = 7、10、9)(e)和RORγt⁺ Treg细胞频率(n = 7、10、9)(f)。
g,h, 流式细胞分析及小鼠(同c)空肠TFH13细胞(g)和IL-4⁺及IL-4⁺IL-13⁺ CD4⁺ T细胞(h)频率(n = 7、10、9)。
(a–h数据均以均值±标准误表示。统计分析采用非配对双尾t检验(a、b)、单因素方差分析配合Tukey事后检验(d–h)及重复测量双因素方差分析配合Tukey事后检验(c)。)
我们进一步探究RELMβ在体内耗竭Lactobacillus属的机制。体外实验显示RELMβ对乳杆菌无直接抗菌活性(扩展数据图5g)。鉴于我们已证实RELMβ作为Sprr2a的上游主调控因子(图1h-k及扩展数据图1d,e),且SPRR2A对革兰氏阳性菌具有抗菌功能,我们检测RELMβ调控的SPRR2A是否抑制Lactobacillus。结果显示SPRR2A对乳杆菌具有强效抗菌作用(扩展数据图5g)。随后我们构建了SPRR2A缺陷背景的Il4raF709小鼠。与Il4raF709小鼠相比,Il4raF709Sprr2a−/−小鼠表现出FA保护作用:血清OVA特异性IgE和MMCP1水平降低,Lactobacillus丰度增加,Helios⁻RORγt⁺ Treg细胞频率升高(扩展数据图5h-k)。
吲哚衍生物维持食物抗原耐受
鉴于肠道Lactobacillus属耗竭与RELMβ破坏食物耐受相关,我们深入探究肠道Lactobacillus在维持食物耐受中的作用。首先从Il4raF709Retnlb−/−小鼠粪便中分离Lactobacillus(包括L. murinus和L. reuteri)。随后评估这些Lactobacillus对Il4raF709小鼠FA的保护作用。饲喂Lactobacillus混合物的Il4raF709小鼠受到保护免于过敏反应,并显示肥大细胞脱颗粒抑制和OVA特异性IgE应答减弱(图3c,d)。与内源性小鼠肠道乳杆菌功能类似,三种人源Lactobacillus菌株(L. gasseri、L. reuteri和L. rhamnosus)混合物可抑制FA发生,同时增加SI-LPLs中RORγt⁺ Treg细胞频率并降低TFH13细胞频率(图3e-h)。这些数据表明肠道Lactobacillus通过增加RORγt⁺ Treg细胞发挥FA保护作用。
我们进一步探索Lactobacillus抵抗RELMβ驱动FA的机制基础。已知Lactobacillus等肠道菌可将色氨酸代谢为多种具有免疫调节功能的吲哚衍生物(扩展数据图6a)。靶向质谱(MS)检测显示,OVA-SEB致敏的FA易感Il4raF709小鼠粪便中多个关键色氨酸源性吲哚代谢物(包括吲哚、吲哚-3-甲醛I3A、吲哚-3-乙酸IAA、吲哚-3-乙醇I3E、吲哚乳酸ILA及色胺)水平降低,而在FA受保护的Il4raF709Retnlb−/−小鼠中得以恢复(图4a及扩展数据图6b)。OVA经皮致敏模型中,吲哚衍生物同样受RELMβ调控(扩展数据图3f)。在儿童队列中,FA患者粪便吲哚衍生物水平较健康对照组显著降低(图4b及扩展数据图6c)。通过限制Il4raF709Retnlb−/−小鼠饮食中色氨酸(细菌吲哚代谢物的营养源),可消除RELMβ缺失的FA保护作用,表现为肥大细胞脱颗粒增加、OVA特异性IgE应答增强以及SI-LPL RORγt⁺ Treg细胞频率降低(图4c-f)。相反,给予IAA(而非对照溶剂)可保护Il4raF709小鼠免于FA,并伴随RORγt⁺ Treg细胞水平升高(扩展数据图6d-f)。
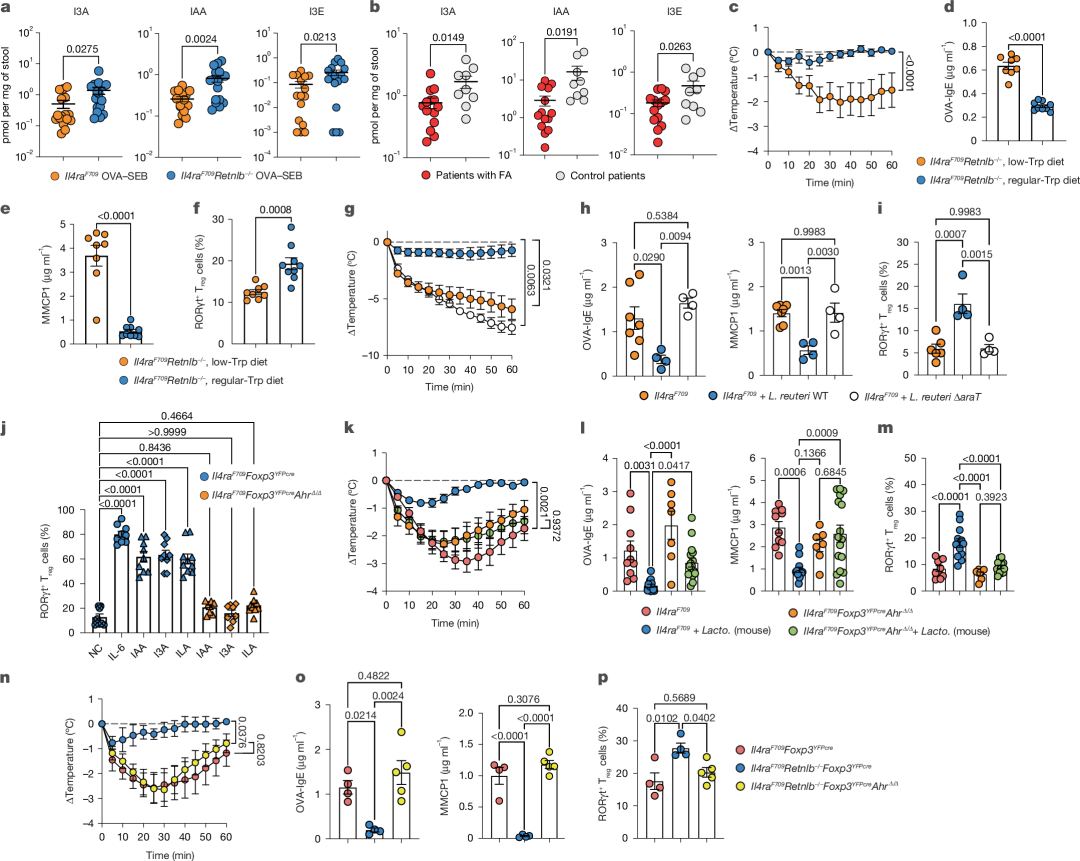
图4 | 肠道微生物组衍生的吲哚衍生物通过AhR诱导FA保护性Treg细胞
a, OVA–SEB致敏的Il4raF709(n = 16、15、16)与Il4raF709Retnlb−/−(n = 17、17、17)小鼠粪便中I3A、IAA及I3E的定量分析。
b, 活动期FA患者(n = 14)与健康对照(n = 9)粪便中I3A、IAA及I3E的定量分析。
c, 饲喂常规色氨酸(n = 9)或低色氨酸饮食(n = 8)的Il4raF709Retnlb−/−小鼠经OVA–SEB致敏及OVA激发后的体温变化。
d-f, 血清OVA特异性IgE(n = 9、8)(d)、MMCP1浓度(n = 9、8)(e)及SI-LPL RORγt⁺ Treg细胞频率(n = 9、8)(f)。
g, 假处理(n = 7)、WT L. reuteri 100-23处理(n = 9)或L. reuteri 100-23-ΔaraT处理(n = 4)的Il4raF709Retnlb−/−小鼠体温变化。
h,i, 血清OVA特异性IgE(h)及MMCP1浓度(h)(n = 7、4、4),以及SI-LPL RORγt⁺ Treg细胞频率(i)(n = 6、4、4)。
j, 初始Il4raF709Foxp3YFPcre或Il4raF709Foxp3YFPcreAhRΔ/Δ T细胞在抗CD3、抗CD28、IL-2和TGF-β单独处理(阴性对照,NC,n = 8)、联合IL-6(n = 9)或联合IAA(n = 9、8)、I3A(n = 9、10)、ILA(n = 10、10)诱导分化的RORγt⁺ Treg细胞频率。
k, OVA–SEB致敏及OVA激发的假处理(n = 10)或鼠源Lactobacillus处理(n = 13)Il4raF70小鼠,与假处理(n = 7)或鼠源Lactobacillus处理(n = 16)Il4raF709Foxp3YFPcreAhrΔ/Δ小鼠的体温变化。
l,m, 血清OVA特异性IgE(n = 10、13、7、16)及MMCP1浓度(n = 10、13、7、16)(l),以及SI-LPL RORγt⁺ Treg细胞频率(n = 10、13、7、15)(m)。
n, OVA–SEB致敏及OVA激发的Il4raF709Foxp3YFPcre(n = 4)、Il4raF709Foxp3YFPcreRetnlb−/−(n = 4)及Il4raF709Foxp3YFPcreAhrΔ/ΔRetnlb−/−(n = 5)小鼠的体温变化。
o,p, 血清OVA特异性IgE(n = 4、4、5)及MMCP1浓度(n = 4、4、5)(o),以及SI-LPL RORγt⁺ Treg细胞频率(n = 4、4、5)(p)。
(a–p数据均以均值±标准误表示。统计分析采用非配对双尾t检验(a,b、d–f)、单因素方差分析(h–j、l、m、o、p)配合Šídák(j)或Tukey(h、i、l、m、o、p)事后检验,或重复测量双因素方差分析配合Tukey事后检验(c、g、k、n)。)BSE对空腹血糖的改善效果在不同病理生理亚组之间存在差异(见补充表6)。具有轻度年龄相关糖尿病(MARD)特征的参与者在接受BSE治疗后,空腹血糖改善更为显著,与安慰剂相比,平均差异为0.4 mmol/L(95%置信区间为-0.6到-0.1;接受BSE治疗的参与者n=24,接受安慰剂的参与者n=20;P=0.008;见扩展数据图3和4)。此外,这些参与者的胰岛素分泌也有所改善,通过HOMA-B(胰岛β细胞功能的HOMA评估)测量,改善范围为3.3–26.0,P=0.02。相比之下,在具有严重胰岛素抵抗型糖尿病(SIRD)或轻度肥胖相关糖尿病(MOD)特征的参与者中,BSE与安慰剂之间没有显著差异。当将治疗(BSE或安慰剂)和聚类分组作为交互项进行评估时,观察到它们对空腹血糖变化具有统计学显著的交互作用(使用线性模型,P=0.008;n=74;见补充表6)。这表明BSE对血糖的改善效果因聚类分组而异。
为明确肠道微生物组源性吲哚衍生物在FA保护中的作用,我们构建了缺失araT基因(负责从色氨酸生成IAA和I3A)的肠道L. reuteri菌株(扩展数据图7a,b)。该基因缺失不影响L. reuteri的体内适应性(扩展数据图7c,d)。与定植WT L. reuteri后的FA保护相比,定植ΔaraT L. reuteri的Il4raF709小鼠尽管菌株定植水平相当,但粪便IAA水平降低(扩展数据图7d,e),且未表现出FA保护作用,食物抗原激发后仍发生显著过敏反应(图4g)。ΔaraT L. reuteri处理小鼠的血清OVA-IgE和MMCP1水平也升高(图4g,h)。这些数据证实RELMβ通过抑制色氨酸源性肠道微生物组吲哚代谢物促进FA发生。
鉴于FA患者肠道中具有保护作用的免疫调节性吲哚代谢物水平较健康对照降低(图4b及扩展数据图6d),我们探究吲哚代谢物差异是否与肠道微生物组组成相关。通过分析低血清RELMβ健康人群与高RELMβ活动性FA患者的肠道微生物组,发现后者拟杆菌目(Bacteroidales,人肠道主要革兰氏阴性菌)成员Alistipes和Parabacteroides丰度降低(扩展数据图8a)。值得注意的是,与FA受保护的RELMβ缺陷Il4raF709小鼠类似,FA易感Il4raF709小鼠中Alistipes也显著减少(图3a)。这表明人肠道Alistipes与小鼠乳杆菌和Alistipes类似,受RELMβ调控(扩展数据图8b)。我们发现人源Alistipes shahii可被SPRR2A杀灭,且与Lactobacillus属不同,该菌还直接受RELMβ抑制(尽管程度弱于SPRR2A)(扩展数据图8c,d)。
在证实RELMβ调控FA小鼠模型中的Alistipes、Alistipes受RELMβ调控的SPRR2A直接抑制,且人FA中Alistipes减少与RELMβ升高及吲哚代谢物降低相关后,我们探究Alistipes是否与Lactobacillus属类似,通过杯状细胞-RELMβ-肠道微生物组-吲哚轴参与FA。分析166个人肠道Alistipes基因组发现,72%的基因组含有色氨酸生成吲哚必需基因tnaA同源序列。在Alistipes finegoldii、Alistipes putredinis、Alistipes onderdonkii和A. shahii等与人类健康及疾病相关的菌种中,tnaA存在率达94%-100%(补充表3),与既往报道的人肠道拟杆菌目色氨酸酶丰度特征一致。我们发现A. shahii通过色氨酸分解代谢显著产吲哚(扩展数据图8c)。由于Alistipes难以进行基因操作,我们选取人肠道拟杆菌目产吲哚菌株Bacteroides ovatus(含tnaA且产吲哚)进行研究(扩展数据图8c)。为验证拟杆菌目源性吲哚是否介导FA保护,我们在B. ovatus中构建非极性tnaA缺失突变株(ΔtnaA),证实其产吲哚能力缺陷(扩展数据图8c)。用WT或ΔtnaA B. ovatus定植Il4raF709小鼠后发现,尽管定植水平相似,但ΔtnaA菌株无法挽救FA(扩展数据图8e-k)。值得注意的是,B. ovatus与Alistipes和Lactobacillus类似,受RELMβ依赖性SPRR2A抑制(扩展数据图8d)。综上,除Lactobacillus外,其他肠道微生物组成员(包括拟杆菌目菌种)也受肠道上皮RELMβ调控,其丰度与人类FA保护相关,并能通过产吲哚或其衍生物在小鼠FA模型中介导食物抗原口服耐受。
肠道微生物组源性吲哚衍生物诱导FA保护性Treg细胞
与WT L. reuteri相比,定植ΔaraT L. reuteri的Il4raF709小鼠无法驱动耐受性RORγt⁺ Treg细胞在FA中的扩增(图4i)。这表明微生物组源性FA保护性吲哚代谢物可能直接影响RORγt⁺ Treg生成。首先,我们检测微生物组源性吲哚衍生物(如IAA、I3A和ILA)诱导Il4raF709小鼠初始T细胞分化为RORγt⁺ Treg的能力。特定吲哚衍生物可替代IL-6作用,显著诱导RORγt⁺ Treg细胞分化(图4j及扩展数据图7f)。我们进一步探究微生物组源性吲哚衍生物驱动RORγt⁺ Treg分化的机制。化学感应芳香烃受体(AhR)在Treg细胞中表达,且AhR激活剂可驱动Treg细胞扩增。已知IAA、I3A和ILA是AhR激动剂,提示肠道微生物将膳食色氨酸代谢为吲哚衍生物可能通过Treg分化生理性驱动口服耐受。为验证此机制,我们将携带条件性Ahr等位基因的小鼠与Il4raF709Foxp3YFPcre小鼠交配。Treg细胞中Ahr敲除可消除微生物组源性吲哚衍生物诱导RORγt⁺ Treg分化的能力(图4j)。进一步检测微生物组源性吲哚衍生物对食物抗原耐受的作用发现,在FA易感Il4raF709背景小鼠中,Treg细胞AhR缺失可消除乳杆菌介导的FA保护作用,表现为显著过敏反应、OVA-IgE和血清MMCP1水平升高,且无法扩增耐受性RORγt⁺ Treg细胞(图4k-m)。此外,Treg细胞特异性Ahr敲除消除了Il4raF709Retnlb−/−小鼠中RELMβ缺失的FA保护作用,致敏并激发后的Il4raF709Retnlb−/−Foxp3YFPcreAhr∆/∆小鼠出现强烈过敏反应且RORγt⁺ Treg细胞减少(图4n-p)。综上表明,杯状细胞分泌的RELMβ通过限制产吲哚代谢物肠道菌的定植来抑制食物抗原耐受,而这些代谢物通过化学感应AhR驱动耐受性Treg细胞介导口服耐受并预防FA。
生命早期RELMβ失调促进成年期FA发生
FA在婴儿期和幼儿期尤为常见,有证据表明遗传因素及生命早期微生物暴露均参与FA易感性的形成。为明确RELMβ在生命早期口服耐受中的作用,我们首先检测RELMβ缺失的FA保护效应是否可垂直传递。通过Il4raF709Retnlb−/+小鼠互交发现,Retnlb正常的Il4raF709子代小鼠与同窝Il4raF709Retnlb−/+小鼠表现出同等程度的FA保护作用,且伴随肠道Lactobacillus属富集,提示FA结局可能受RELMβ调控并以垂直传播方式影响(扩展数据图9a-f)。
我们进一步探究断奶期(生命早期关键窗口期,与FA终生易感性形成及RORγt⁺ Treg细胞分化加速相关)RELMβ对口服耐受获得的影响。值得注意的是,Retnlb转录本在断奶前后时期(出生后第3-4周)表达升高并随后下降,而Il4raF709小鼠该阶段的表达增幅显著高于WT小鼠(图5a)。与WT小鼠相比,Il4raF709小鼠断奶期SI-LPLs中RORγt⁺ Treg细胞频率降低,与Retnlb异常表达相呼应(图5b)。为直接验证RELMβ表达抑制RORγt⁺ Treg细胞的作用,我们对WT小鼠从出生后第2周至第4周隔日给予重组RELMα、RELMβ或对照溶剂处理。结果显示,RELMβ(而非RELMα或对照溶剂)可抑制第4周时SI-LPLs RORγt⁺ Treg细胞分化(图5c)。此外,生命早期接受RELMβ注射的WT小鼠在成年期表现出FA易感性(扩展数据图10a-c)。反之,在生命早期给予未致敏Il4raF709小鼠抗RELMβ单克隆抗体处理可诱导RORγt⁺ Treg细胞分化(图5d),并伴随Lactobacillus丰度增加(图5e)。这些数据表明生命早期RELMβ表达调控RORγt⁺ Treg细胞分化。
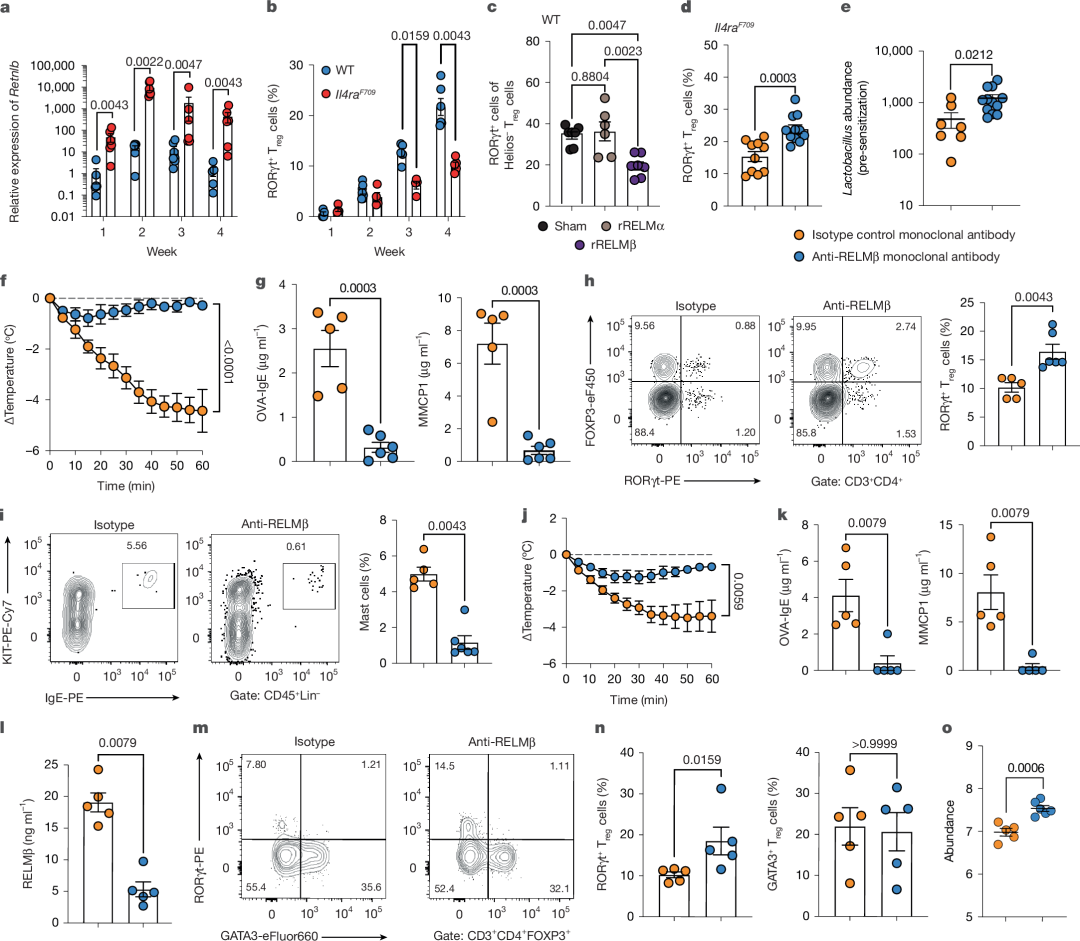
图 5 |生命早期抑制RELMβ预防未来FA发生
a,生命前4周WT与FA易感Il4raF709小鼠小肠组织Retnlb转录本的qPCR分析(WT:n = 6、6、7、5;Il4raF709:n = 6)。
b,生命前4周WT与FA易感Il4raF709小鼠SI-LPL RORγt⁺ Treg细胞频率(WT:n = 3、7、5、5;Il4raF709:n = 4或5)。
c,生命早期(第2-4周)接受RELMβ(n = 7)、RELMα(n = 6)或载体(n = 6)处理的WT小鼠SI-LPL RORγt⁺ Treg细胞频率。
d,生命第2-4周隔日接受抗RELMβ单抗(n = 12)或同型对照单抗(n = 10)处理的Il4raF709小鼠SI-LPL RORγt⁺ Treg细胞频率。
e,生命第4周时,接受抗RELMβ单抗(n = 7)或同型对照单抗(n = 12)处理的Il4raF709小鼠Lactobacillus属丰度。
f,生命第2-4周接受抗RELMβ单抗(n = 6)或同型对照单抗(n = 3)处理的Il4raF709小鼠经OVA–SEB致敏及OVA激发后的核心体温变化。
g,生命早期单抗处理小鼠(n = 5、6)的总血清IgE、OVA特异性IgE及MMCP1浓度。
h,i, 生命早期单抗处理小鼠(n = 5、6)的SI-LPL RORγt⁺ Treg细胞(h)及肥大细胞(i)频率。
j, 生命早期接受抗RELMβ单抗(n = 10)或同型对照单抗(n = 12)处理并持续8周OVA–SEB致敏的Il4raF709小鼠经OVA激发后的体温变化。
k, 来自j组的小鼠(n = 5、5)的OVA特异性IgE及MMCP1浓度。
l, 来自j组的小鼠(n = 5、5)的血清RELMβ浓度。
m,n, 来自j组小鼠(n = 5、5)SI-LPL RORγt⁺ Treg细胞及GATA3⁺ Treg细胞的流式分析(m)与频率(n)。
o 激发后接受抗RELMβ单抗或同型对照单抗处理的Il4raF709小鼠Lactobacillus属丰度(n = 5、6)。
(a–o数据均以均值±标准误表示。统计分析采用混合效应双因素方差分析配合Šídák事后检验(b)、非配对双尾t检验(a、d、e、g–i、k、l、n、o)、单因素方差分析配合Tukey事后检验(c)及重复测量双因素方差分析配合Šídák事后检验(f、j)。)
靶向RELMβ预防和治疗FA
为明确生命早期抑制RELMβ能否预防FA易感小鼠成年期发生FA,我们在Il4raF709小鼠断奶关键期(第2-4周)隔日给予抗RELMβ单克隆抗体处理,并于10周龄进行口服致敏及后续食物抗原激发。生命早期抗RELMβ治疗可保护小鼠免于FA,抑制过敏反应(图5f),减少肥大细胞扩增,并增加SI-LPL RORγt⁺ Treg细胞频率(图5g-i)。这些结果表明生命早期RELMβ活性可影响FA终生易感性,靶向RELMβ可预防成年期FA发生。
我们进一步评估抗RELMβ单克隆抗体治疗成年FA模型Il4raF709小鼠能否重置免疫失调并重塑耐受性微生物组。给予成年Il4raF709小鼠致敏期抗RELMβ单克隆抗体(而非同型对照抗体)治疗,可消除食物抗原口服激发后的过敏反应,抑制FA相关Th2细胞应答,并伴随乳杆菌丰度增加(图5j-o及扩展数据图10d-f)。这表明在FA临床前模型中,生命早期异常的免疫耐受印记可通过靶向RELMβ在成年期重置。
本研究阐明了一个调控2型肠道免疫与FA的"免疫-上皮-微生物组"回路。肠道对食物抗原的2型免疫应答可能通过两个并行回路运作:第一为传入性"微生物组-上皮-免疫"回路,即化学感应肠道上皮细胞感知微生物代谢物后驱动2型免疫介质(IL-4、IL-13);第二为传出性"免疫-上皮-微生物组"回路,即2型介质诱导分泌性上皮细胞产生RELMβ,后者通过微生物组编辑调控RORγt⁺ Treg细胞对食物抗原特异性免疫应答的控制。该回路中鉴定的细胞模块(肠道分泌上皮、RORγt⁺ Treg细胞、Lactobacilli)与分子模块(RELMβ、吲哚衍生物、AhR)均为FA防治提供了干预靶点。生命早期RELMβ失调可能通过破坏断奶反应期(伴随固体食物引入而微生物组扩增的关键免疫耐受形成窗口)的RORγt⁺ Treg细胞应答,导致FA终生易感性。因此,靶向RELMβ不仅可抑制FA发生,更能通过重塑微生物组建立长期口服耐受。
作者简介
Seth Rakoff-Nahoum(通讯作者)

Rakoff-Nahoum 实验室的目标是全面理解宿主相关微生物群在各个生物学层次上的作用:从基因到分子、从生物体到生态系统,并且重点在于确定因果关系。为此,我们将实证方法与生态和进化理论相结合。我们的研究工具包括:利用经典肠道厌氧菌的细菌遗传学技术,对哺乳动物微生物群(如拟杆菌、厚壁菌门、放线菌)的单个成员进行培养、随机和定向诱变;使用体内和体外实验系统(例如 TnSeq)研究肠道细菌对环境的适应;应用小鼠无菌模型、计算方法探索微生物群生态学;构建高通量体外管线,对微生物群成员之间以及它们与宿主相互作用的培养、遗传、化学和表型进行分析。
Rakoff-Nahoum 实验室目前的研究重点集中在以下几个(非互斥的)方面:肠道及女性生殖道微生物群中合作与竞争的遗传和分子机制;微生物代谢产物在肠道微生物生态及其与宿主相互作用中的作用;宿主-微生物群相互作用的糖生物学;人群中微生物群的生态学,特别关注儿童健康与疾病;微生物群对膳食和微生物活性分子的代谢。
Talal A. Chatila(通讯作者)

Chatila 的研究主要致力于阐明免疫耐受破坏的机制,涵盖单基因免疫疾病及包括哮喘和食物过敏在内的常见过敏性炎症疾病,尤其关注调节性 T 细胞在疾病发生中的作用。这项研究已鉴定出多个调控人体免疫耐受的基因网络,包括 FOXP3、DOCK8、LRBA 和 IL4R 相关基因。
近年来,Chatila 团队进一步揭示了共生微生物群和 Notch 信号通路的炎症信号如何调控调节性 T 细胞的功能,并确立了致病性调节性 T 细胞重编程在常见过敏性疾病中导致耐受破坏的作用。这些研究的长期目标是通过调控调节性 T 细胞特异性的分子和代谢机制,开发恢复免疫耐受的新型治疗方法。Chatila 的研究主要聚焦于人类免疫疾病的遗传与调控机制。
翻译:皮天宇,中国农科院沼科所,硕士在读;
审核:朱志豪,广东医科大学,基因组所联合博士后;
终审:刘永鑫,中国农科院基因组所,研究员/博导;
排版:荀佳妮,中国农科院基因组所,生物信息学硕士在读
宏基因组推荐
本公众号现全面开放投稿,希望文章作者讲出自己的科研故事,分享论文的精华与亮点。投稿请联系小编(微信号:yongxinliu 或 meta-genomics)
猜你喜欢
iMeta高引文章 fastp 复杂热图 ggtree 绘图imageGP 网络iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









