





 文章介绍了神经退行性疾病如帕金森病、阿尔茨海默病等中脑网络分析的进展,包括使用PET和fMRI识别疾病特异性网络,以及这些网络如何与病理过程和治疗响应相关。多变量分析和图论方法在识别异常功能连接和预测疾病进展方面显示出潜力,可用于提高诊断准确性并监测治疗效果。
文章介绍了神经退行性疾病如帕金森病、阿尔茨海默病等中脑网络分析的进展,包括使用PET和fMRI识别疾病特异性网络,以及这些网络如何与病理过程和治疗响应相关。多变量分析和图论方法在识别异常功能连接和预测疾病进展方面显示出潜力,可用于提高诊断准确性并监测治疗效果。
网络分析工具正越来越多地应用于静息代谢活动成像(PET)或血液氧合依赖信号成像(功能MRI),以表征脑疾病的异常神经回路。这种方法对于神经退行性疾病的研究特别有价值,这些疾病的特征是沿着离散的神经通路扩散的。识别和验证疾病特异性脑网络有助于定量评估神经回路随时间和治疗过程的变化。网络异常通常可以在症状出现前就被识别出来,甚至在临床前阶段也可以用于跟踪疾病的进展。同样地,网络活动可以通过治疗来调节,因此可以在临床试验中被用作治疗疗效的标记。最后,早期的鉴别诊断可以通过同时测量单个患者的多个疾病网络的活动水平来实现。虽然这些技术最初是为PET开发的,但在过去的几年里,类似的方法已经被引入用于功能MRI,这是一种更容易获得的无创成像方式。这一进展有望扩大网络工具的应用范围,以适应大规模和多样化的患者群体。本文发表在Nature Reviews Neurology杂志。
一、介绍
神经退行性脑疾病是全球范围内老年人群(年龄>75岁)残疾的主要原因之一。2019年,总共约有5700万人患有痴呆症,还有850万人患有帕金森氏症。这些疾病的流行率在过去的三十年中显著上升。治疗方案目前仅依靠症状来选择,但疾病修饰疗法是以减缓或停止潜在的病理过程为药物开发的一个重点。准确的诊断对于新疗法试验的成功是必要的。但是仅凭临床依据诊断常见的神经退行性疾病只有中等准确度(70-80%)。虽然脑组织的组织病理学检查仍然是诊断的金标准,但详细了解涉及的分子和解剖途径,并使用体内生物标志物来支持可靠的,早期的诊断也越来越重要。
在过去的十年里,神经成像研究已经从映射局部效应过渡到开发神经活动的预测模型,该模型整合了分布在健康和疾病中的多个大脑系统中的信息。基于主成分分析(PCA)的大脑活动多变量模式分析(MVPA)是数据驱动的,可以设计在没有预先指定的兴趣区域,来区分不同疾病状态(如疾病和健康的状态),评估个体行为差异或表现差异(包括健康个体或特定人群的认知功能),并评估疾病和/或治疗随着时间的效应(例如,在随机临床试验中,比较接受药物治疗的患者与接受安慰剂治疗的患者)。各种成像已被用于识别和评估功能脑网络模式:[18F]氟脱氧葡萄糖(FDG)PET代谢成像;基于灌注成像的H2 15O PET、单光子发射CT(SPECT)或18F标记放射性示踪的早期动态PET;非侵入性MRI技术,如静息状态(rs)功能MRI(fMRI),它记录局部血氧水平依赖(BOLD)信号的波动,以及动脉自旋标记(ASL)MRI来绘制脑区内脑血流。由于神经血管耦合,大脑糖代谢的功能脑成像和大脑血流相关的功能脑成像通常提供互补信息。也就是说,这两种措施可以在某些疾病状态下分离,如缺血性中风、创伤性脑损伤和胶质瘤以及在特定的治疗条件下。例如,在PD患者中,左旋多巴给药诱导多巴胺反应型PD相关运动网络区域的血流和糖代谢分离,导致大量的神经血管解耦。尽管它们在方法上存在差异,但这些成像模式都被用于疾病网络定量评估的前瞻性研究。事实上,脑成像和网络量化的多变量方法的临床效用来自于这种方法,它被称为Topographic Profile Rating (TPR)(Box 1)。
Box 1
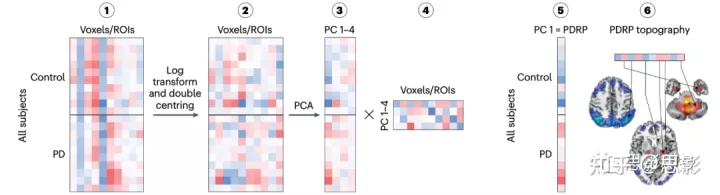
代谢性脑网络模式识别 主成分(PC)分析(PCA)是一种统计方法,通过创建新的不相关变量来降维,这些变量可以解释数据中的最大差异。尺度子配置模型(scaled subprofile model, SSM)是一种基于PCA的方法,它能够识别与疾病相关的空间协方差拓扑结构。来自一组患者(在本例中,帕金森病(PD))和健康对照个体的脑扫描的SSM-PCA,不受临床类别的影响,导致一个或多个线性独立(即正交)空间协方差模式,称为PCs。PCs解释了成像数据中受试者-脑区(体素)的相当一部分差异,并在患者和对照组之间显示了表达水平的最大差异。简而言之,为了确定一种稳定的疾病相关模式,我们分析了10-20个来自患者的扫描和来自年龄匹配和性别匹配的健康对照个体的类似数量的扫描。在预处理过程中,每个扫描都被空间归一化和平滑,然后(步骤1)转换为一个被试的体素数据矩阵(为了方便起见,这个步骤使用感兴趣区域(roi)而不是体素来描述)。灰质的提取使用阈值或通过应用一个mask。其余的体素进行对数变换(第2步),通过减去行和列均值对矩阵进行双中心化,以产生被试残差轮廓(subject residual profile,SRP)。 将PCA应用于受试者-体素协方差矩阵,获得受试者得分向量(步骤3)和相关的特征值和PCs(步骤4)。在最前面的PCs被选择并进入一系列逻辑回归模型,用于根据它们的表达水平(受试者得分)对扫描进行分类。选择Akaike信息最低的模型(步骤5)作为疾病相关网络(步骤6)。 还需要进一步的步骤来确认疾病相关网络的有效性和生物学意义。体素权重的稳定性可以通过bootstrapping或留一交叉验证(leave-one-out cross-validation)方法来测试。与疾病相关的网络表达在独立的患者队列中进行前瞻性的验证。这种计算过程被称为topographic profile rating (TPR)。所得到的值量化了个体病例的模式表达,并已广泛用于评估疾病进展和鉴别诊断。TPR对轻度认知障碍患者转化为阿尔茨海默病,以及快速眼动睡眠行为障碍患者转化为PD或路易体痴呆的预后价值已被证明。然而,目前尚无研究探讨TPR在预测轻度认知障碍患者转化为帕金森病或路易体痴呆中的价值。PDRP,pd相关模式。 |
在本文中,我们总结了神经退行性疾病患者脑成像网络分析的进展,包括帕金森病、阿尔茨海默病、路易体痴呆(DLB)和额颞叶痴呆(FTD)。我们专注于识别和验证疾病特异性网络的技术,并考虑这些网络与潜在的病理过程和疾病相关基因型之间的关系。此外,我们还讨论了使用图论分析来描述这些疾病中可能发生的内部网络变化的研究结果。最后,我们讨论了脑网络分析的潜在临床应用:提高诊断准确性,预测功能衰退,跟踪疾病进展和监测治疗效果。
二、功能脑网络
多变量算法
单变量算法已被广泛用于定位脑功能的区域异常,但这些技术侧重于体素-体素比较,而没有明确考虑空间分布的脑区团块之间的关系。相比之下,多变量分析特别关注这些关系,可用于识别成像数据中疾病相关变异的来源。多变量分析已用于基于氟脱氧葡萄糖(FDG)PET的代谢连通性研究,并使用多种方法从患者扫描数据中提取脑网络信息。重要的是,PCA等方法能够表征与疾病状态密切相关的协方差拓扑,同时通过降维促进成像的生物学解释。用于研究大脑疾病的最流行的基于PCA的算法之一是尺度子配置模型(scaled subprofile model, SSM)(Box 1)。一种基于监督PCA的相关方法,称为有序趋势分析(OrT,一种典型变量分析(CVA)的一种形式)随后被开发来处理个体在不同试验条件下的网络改变,如发生在疾病进展和治疗试验中,在任何一种方法中,衍生的网络都必须通过前瞻性试验来验证。
代谢PET与rs-fMRI
整合来自脑成像研究的多模态数据可以提供有关网络功能的相关信息,特别是在神经退行性疾病患者中。通过FDG PET评估,大脑中的局部葡萄糖代谢与局部突触活动耦合,尽管一些证据表明,在病理条件下,邻近的小胶质细胞的炎症反应可能有助于葡萄糖利用信号的增加。FDG PET产生几分钟的大脑代谢活动的放射自显影图像,rs-fMRI以BOLD信号的形式记录血氧合的瞬时血流动力学波动,该信号与神经元活动耦合。尽管如此,由于rs-fMRI对发生在比FDG PET更短的时间窗口内的事件很敏感,rs-fMRI和FDG PET可能提取不同的区域关系。
在FDG PET中,协方差拓扑通过经典的矩阵分解算法进行识别,通过该算法,将成像的大型有序行和列,降维为更小的统计显著成分集。在rs-fMRI中,网络的提取依赖于关键脑区记录的BOLD信号时间序列之间的关系。为此,基于种子的方法评估了在预先指定的“种子”脑区测量的时间序列与在其他脑区记录的活动之间的相关性。协方差方法,如PCA,并不是最优的应用于时空数据,如那些由BOLD信号提供的数据。也就是说,盲源分离的方法,如独立成分分析(ICA),提供了一种数据驱动的算法来消除rs-fMRI数据中的独立源信号的歧义。与PCA一样,ICA寻求从扫描数据中提取离散的多脑区成分。然而,与PCA和其他基于相关性的方法不同,ICA将信号相关联,同时降低了高阶统计相关性。因此,ICA识别最大独立的信号,而PCA识别一个或多个线性独立的模式(正交特征向量),这些模式可以解释数据的最大差异。ICA已被用于研究群体和个体患者的功能连接。事实上,已经开发了一种类似于SSM-PCA的策略,其中ICA用于识别和验证PD患者的rs-fMRI数据中可重复的疾病网络(Box 2)。事实上,使用来自神经退行性疾病患者的FDG PET或rs-fMRI数据,已经确定了类似的网络特征。
Box 2
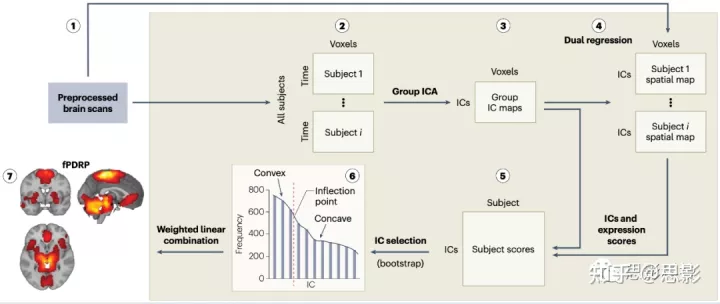
基于功能MRI的网络模式识别 经过预处理(步骤1)后,将健康对照个体和疾病(帕金森病(PD))患者)的静息态(rs)功能(f) MRI扫描转化为单一体素时间序列矩阵(步骤2),然后进行空间组独立成分分析(ICA),生成组水平独立成分(IC)图(步骤3)。为了方便起见,只显示受试者1和i,但在实践中,将分析许多患者和对照组的扫描结果。采用自举法(Bootstrap)重采样来评估组水平的独立成分的可靠性。为了获得成像数据的ICA空间图,采用双回归方法估计每个个体的时间序列和空间图(步骤4)。在网络识别组中每个个体的rs-fMRI数据中的每个独立成分的每个受试者得分将会被计算。这些值代表了个体成像中特定独立成分的表达,它们被计算为相应的个体和组空间图的点积(步骤5)。然后将所有独立成分输入一个logistic回归模型,通过自举(bootstrap)重抽样,以识别最佳分离患者和对照个体的独立成分子集(步骤6)。按下降频率顺序排列的直方图显示了每个独立成分对logistic模型的相对贡献。离散曲线内的拐点用于识别最相关的独立成分用于模式生成。将这些独立成分的相应受试者得分输入第二组logistic模型,并再次使用自举法重新采样来估计每个选定独立成分的相应回归权重(系数)(步骤7)。 基于fmri的pd相关模式(fPDRP)的表达水平被计算为每个选定独立成分的受试者得分的线性组合,通过相应的回归系数加权。fPDRP可应用于来自前瞻性识别的个体患者的rs-fMRI数据。使用双重回归,估计每个患者的空间图。fPDRP的受试者得分是通过所选独立成分的表达水平的线性组合来计算的。 |
基于图论的网络评价
多变量方法,如PCA和ICA,提供了关于特定脑区(或“节点”)的功能信息和对整体网络活动的相对贡献,也就是说,定义一个网络的内部结构的脑区到脑区的连接(“边”)也同样重要。图论是数学的一个分支,通过描述形成疾病网络的特定脑区和连接,提供了一种有价值的方法来描述功能结构的变化。节点可以通过多种方式进行定义,从单个体素或脑区团块到基于标准化的结构图谱的解剖定义的感兴趣区域。节点之间的连接通常由距离度量来定义,其中相关性和部分相关性最常用。基于图论的计算算法也被用于在整个网络空间中隔离小网络(称为模块或子图)。可以通过严格计算相关图度量,评估不同个体、群体、不同时间点和不同处理条件下的连接模式。
图中的连接可以在局部或全局水平上进行探索。在局部水平上,最简单的连通性度量是节点度中心性,它是由单个节点与其他节点的连接数量决定的。相比之下,全局度量可以用来描述网络作为一个整体的连接模式,这反过来又会通过图来影响信息流。在这方面,网络属性如特征路径长度、聚类系数、小世界性、模块性和协调性可以用来识别与疾病状态和治疗条件相关的全网络连接模式。事实上,在模块层面上,这些措施可能有助于区分疾病网络中的适应性和病理性连接变化。
三、运动相关神经变性
帕金森病及相关疾病
PD的病理特征是路易体形成和沉积α突触核蛋白聚集在大脑,开始在下脑干和嗅觉系统,扩散到中脑参与黑质致密部和最终导致新皮质区神经退行性变。PD的临床特征倾向于与这些病理改变的进展和功能网络活动的改变相关。例如,黑质中多巴胺能神经元的丢失会导致一组皮质-基底神经节-丘脑-皮质运动回路的失调,从而导致该疾病的主要症状——运动迟缓、僵硬和震颤。虽然帕金森病的临床进展通常与上述病理改变相似,但也有例外,如在运动症状出现前出现细微的认知变化。涉及单胺能和胆碱能的脑干和基底前脑神经递质通路功能障碍可能发生在疾病早期,在α突触核蛋白沉积在大脑皮层之前。无论如何,大多数帕金森病患者在发病过程中都会出现可测量的认知缺陷。事实上,帕金森病的运动和认知临床表现都与特征性脑网络有关,通过这些网络,神经退行性变的局灶性脑区以疾病特异性的方式影响下游脑区的功能。
一些病理上不同的神经退行性疾病也有帕金森症状,但通常预后比特发性帕金森病更差,并且对用于治疗帕金森的药物和手术干预没有反应。非典型帕金森病综合征(APSs),包括多系统萎缩(MSA)、进行性核上麻痹(PSP)和皮质-基底变性(CBD),可能很难与PD区分,特别是在这些疾病的早期临床阶段。尽管它们的临床特征相似,但非典型帕金森病综合征的病理改变的序列与PD不同,与这些疾病相关的网络模式也不同。涉及功能性脑成像数据的多变量方法能够识别疾病特异性网络,可用于评估和区分这些疾病。
(1)帕金森病运动网络
使用FDG PET和SSM-PCA确定的帕金森相关模式(PDRP)已在全球十几个患者和健康个体队列中得到广泛验证。PDRP最初在北美人群中发现,随后在欧洲、亚洲和美国的PD患者独立队列中重复。PDRP的特征是壳核、苍白球、丘脑、脑桥和小脑的糖代谢增加,而运动前和后顶叶区域的糖代谢相对减少。这种模式在PD患者中的表达水平与运动迟缓和僵硬的严重程度相关,并反映了皮质纹状体-苍白球-丘脑-皮质运动回路的变化。其他回路涉及帕金森性震颤,与小脑-丘脑-皮层运动回路的一个单独网络的表达增加有关。在不同的研究中心使用不同断层扫描的独立人群中,尽管仪器和扫描程序不同,但是PDRP(帕金森相关模式)非常相似,而且它们各自的表达水平在个体患者水平上密切相关。虽然PDRP最初是在轻度至中度PD患者中发现的,这些患者之前已经开始每日使用多巴胺能药物治疗,但在未用药的患者中也发现了同样的模式。
值得注意的是,当按半球测量时,单侧肢体和双侧肢体受累患者的PDRP表达水平与同侧和对侧的升高程度相似。PDRP(帕金森相关模式)表达的这种对称性与特发性帕金森病的临床特征和黑质纹状体多巴胺能功能障碍的不对称性形成对比,表明这些测量方法捕获了潜在疾病过程的不同方面。然而,在缺乏(罕见的)MSA或PSP不对称患者表现的数据的情况下,尚不清楚是否这些疾病也会发生对称的脑网络异常。
虽然一种使用rs-fMRI来描述这些疾病网络的算法仅在5年前才开发出来,但由此产生的基于fMRI的PD相关模式(fPDRP)已经被独立验证。在两个独立的患者群体中发现的fPDRP网络的模式结构相似,而对患者的两种成像分析中,基于FDG PET的PDRP和fPDRP表达水平密切相关。在使用H215O PET或ASL MRI获得的PD患者的灌注扫描中,观察到FDG PET PDRP的表达水平异常升高。SSM-PCA也被用于识别PD患者和健康个体的结构MRI和ASL MRI中的PDRP。但这两种成像方式区分PD患者与健康个体的能力都不如FDG PET准确。
基于图论的新分析技术为健康和疾病中的大脑网络组织提供了见解。我们的研究表明,PDRP由两个不同的子网络组成:一个核心区域,由一组位于壳核、苍白球和丘脑的代谢活跃和紧密相连的节点组成;和一个外周区,由代谢较不活跃和相对松散连接的皮质节点组成。在随后的研究中,我们使用传统的检测方法证实了PDRP的核心-外围结构。在这方面,图分析显示在PD患者中,这两个PDRP模块有不同的连接模式和异常的信息流。
(2)帕金森病认知网络
PD患者的认知障碍与正常静息态网络(特别是默认模式网络(DMN))的中断有关和与记忆受损和执行功能障碍相关的异常PD相关认知网络的表达增加相关。帕金森病患者的认知功能障碍被认为涉及多种机制,包括多巴胺能和胆碱能通路的变性以及边缘和新皮质区中路易体的进展。这些变化如何影响正常和与疾病相关的大脑网络的功能是一个正在进行的研究课题。
PD认知相关模式(PDCP)最初是在无痴呆患者的FDG PET数据中发现的,这些PD无痴呆患者有一系列神经心理表现得分,随后在4个独立队列患者中得到验证。基于rs-fMRI的PD认知相关模式(fPDCP)随后使用改进的ICA算法进行了识别和验证。对153名PD患者进行神经心理评估的FDG PET扫描分析,显示PDCP表达水平的增加与正常DMN活动的丧失之间存在很强的负相关。虽然PDCP和DMN共享约50%的脑区差异,但这两个网络是不可互换的。事实上,我们的研究小组发现PD患者的fPDCP反映了DMN腹侧成分的丢失(对应于后扣带区和楔前叶)的丢失,而这些个体的DMN的前和后成分保持完整。除了腹侧DMN的缺失外,fPDCP还包括来自其他大脑区域的贡献,如内侧颞叶和外侧额顶叶区域。值得注意的是,PDRP(帕金森相关模式)网络和PDCP( PD认知相关模式)网络在模式结构上是无关的。
DMN主要涉及到记忆和抽象推理相关的认知功能。尽管在PD或阿尔茨海默病患者中已观察到DMN结构和功能的破坏,该网络对神经退行性疾病的贡献以及DMN变化与神经心理测试性能的相关性超出了本综述的范围。
(3)在疾病进展和治疗过程中的网络变化
特发性快速眼动(REM)睡眠行为障碍(RBD)是一种前驱综合征,可演变为临床α-突触核蛋白特发性相关综合征,包括特发性PD、DLB和MSA。事实上,PDRP的表达水平在几个独立的特发性RBD个体组中升高,其水平在健康个体和早期特发性PD患者之间。此外,两项临床随访研究表明,随后发展为PD或DLB的特发性RBD患者在基线时PDRP表达水平明显升高。有限的数据表明,特发性RBD和PDRP低于正常表达的个体更容易发生MSA,而不是特发性PD或DLB,尽管这一建议仍有待证实。另一组研究人员在发病前患有RBD的新发PD患者中发现,一种称为PDRBD-RP的代谢协方差模式比标准PDRP更好地预测PD表型转化。值得注意的是,PDRBD-RP和PDRP主要在运动前皮层有所不同,运动前皮层对PDRBD-RP的贡献大于对PDRP的贡献。
多项研究表明,PDRP的表达随着疾病的进展呈线性进展。一项来自多个成像中心的数据的分析显示,在同一患者中,PDRP的表达水平始终高于PDCP。这种PDRP优势的倾向已经在多个独立PD队列中观察到,这表明该网络在病程中发展得更早,并且比PDCP进展得更快。此外,PDRP的优势与神经病变的Braak序列一致(图1)。在这种情况下,PDRP与Braak III-IV期病理功能等效,而PDCP与Braak V期的病理功能等效(图1)。然而,考虑到PDRP和PDCP空间中多巴胺能、5-羟色胺能和胆碱能通路的复杂相互作用,这种简化的疾病进展模型应该被认为是假设的,有待进一步研究。有趣的是,PDRP在MSA和PSP等APSs中的优势并不明显。
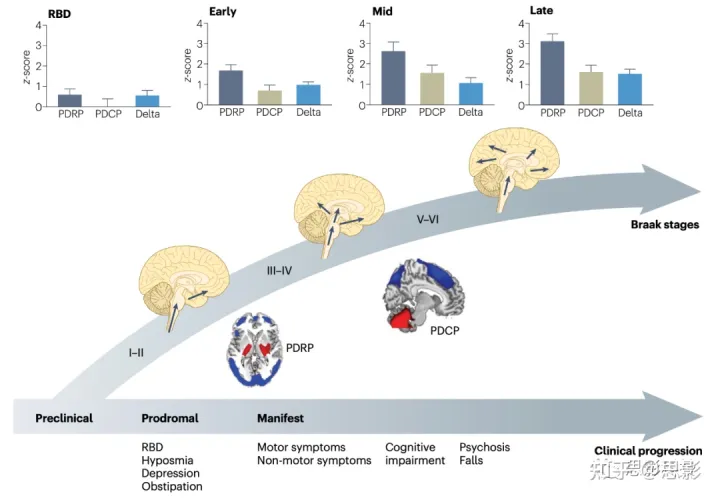
图1|与帕金森病相关的功能脑网络改变。
帕金森病(PD)相关模式(PDRP)的特征是涉及脑干、基底节和边缘区的网络水平的功能改变,这与Braak III-IV期病理相对应。相比之下,PD认知相关模式(PDCP)的特征是功能网络的改变,包括腹侧默认模式网络和其他与Braak V期病理相关的新皮层区域。图中显示,随着疾病进展,PDRP和PDCP表达水平逐步增加,PDRP早于PDCP。有趣的是,在特发性PD患者的个体中,PDRP的表达水平通常大于相应的PDCP值,这导致了一个正向的delta(定义为PDRP和PDCP表达水平之间的差异)。Delta也随着时间的推移而增加。平均PDRP和PDCP表达水平和平均delta按以下顺序显示(快速眼动(REM)睡眠行为障碍(RBD),早期(<4年),中间(4-8年)和晚期(>8年)阶段的16个RBD患者和96名PD患者。
帕金森病的对症治疗与PDRP表达的降低相关,这表明PDRP与临床结果相关。这种效应已在急性左旋多巴给药,丘脑底核(STN)深部脑刺激(DBS)和治疗性丘脑下切开术后中被注意到。考虑到PDRP表达水平与接受DBS手术的PD患者术中记录的自发STN放电率相关,可以预期在该手术后PDRP活性会下降(尽管不是PDCP活性)。左旋多巴治疗对PDCP和PDRP有复杂的影响,因为多巴胺在多个水平上影响基底神经节回路的活动。左旋多巴介导的神经血管效应也可能导致在脑区和网络水平上的血流和代谢的解耦。事实上,这种现象在经历过药物性运动障碍(一种慢性左旋多巴治疗的常见不良反应)的PD患者中尤为显著。
功能成像数据的网络分析已开始在PD的临床试验中发挥重要作用。在这方面,可以在治疗或安慰剂前后评估PDRP和PDCP等网络表达的变化。事实上,类似的方法可以用于剂量发现和衡量治疗对网络进展率的影响。另外,使用监督PCA技术的探索性分析,如OrT-CVA,可以用于盲法试验,以确定特定的治疗相关网络,绘制新的干预措施对大脑通路的影响。例如,该方法已被用于绘制STN腺相关病毒2(AAV2)-GAD(编码谷氨酸脱羧酶1)基因治疗对晚期PD患者引起的连通性变化,所有患者在治疗随机分组前、盲期、双侧基因治疗6个月(n = 16)或假手术(n=21)后6个月和12个月进行FDG PET扫描。有趣的是,在两组患者的试验期间,PDRP的表达均有所增加,这表明该治疗对网络进展没有实质性的影响。事实上,所得到的网络(图2a),研究人员称之为aav2-gad相关模式(GADRP),显示出了一致的结果,即基因治疗后表达的单调增加(图2b)。这些个体中GADRP表达水平的变化与他们在标准化运动测试中随时间的改善相关。图分析显示,在基因治疗后的12个月内,关键GADRP节点的连通性显著增加(图2c)。STN AAV2-GAD基因治疗与小世界性和协调性等异常图指标的改善相关,而假手术后没有发生。这些变化与基因治疗导致的通过GADRP网络空间的更有效的信息流相一致。值得注意的是,在STNAAV2-GAD基因治疗后,这些指标也出现了类似的变化,这表明即使没有显著的模式表达调节,治疗诱导的网络重构也可以发生。
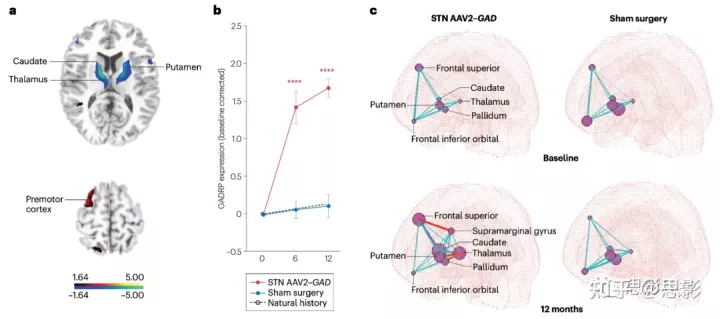
图2|丘脑基因治疗帕金森病的新型功能脑网络。
a,在晚期帕金森病(PD)患者中,通过腺相关病毒2(AAV2)介导的GAD(编码谷氨酸脱羧酶1)传递到丘脑底核(STN)的基因治疗诱导的代谢协方差网络。这种AAV2-gad相关模式(GADRP)的特征是治疗介导的尾状核、壳核、丘脑和额下回的活动减少(蓝色),而顶叶和运动前皮质的活动相对增加(红色)。
b,在一项II期盲法随机临床试验中测量并比较了AAV2-GAD基因治疗和假手术的基线校正的GADRP表达。与假手术组(蓝线)相比,基因治疗组(红线)的网络活动水平增加。自然进展的PD的GADRP表达如黑色虚线。统一PD评分量表(UPDRS)值的变化与GADRP的表达水平密切相关(P < 0.005)。
c,在接受STN AAV2-GAD治疗的患者(左)和接受假手术的患者(右)的基线(上行)和干预后12个月(下行)计算GADRP节点之间的连通性。两组的节点-节点功能相互作用在基线时相似,而治疗组仅在基因治疗后12个月(底部一行)出现了一组新的连接。GADRP节点用半径与相应的节点连通性(度中心性)成正比的球体表示。连接节点对的连接用青色线表示:值r≤0.5,细线;r=0.5-0.6,中线;r≥0.6,粗线。统计上显著的正相关,粗体红线;统计上显著的负相关,蓝色粗体线。为简单起见,只显示左半球。
一种类似的方法被用来探索一种潜在的疾病改善疗法--烟酰胺核糖苷,这是一种每日口服补充剂,可改善线粒体呼吸功能。FDG PET数据是在一项针对早期PD患者的30天的盲法随机安慰剂对照试验中获得的。分析显示,在接受补充剂的个体中存在一种特定的烟酰胺核糖体相关模式(NRRP),但在接受安慰剂的个体中没有。NRRP和PDRP在多个脑区相交。治疗介导的NRRP是否是治疗疗效的可靠标志仍有待未来的研究确定。
(4)基因型效应
帕金森病是一种临床异质性疾病。大多数特发性帕金森病患者都有散发性疾病,但也有少数患者携带与该疾病相关的基因变异。一些成像研究关注于功能回路异常与最常见的PD相关突变:在外显子41 LRRK2的6055位置上出现G>A改变,导致G2019S氨基酸替代亮氨酸-富含重复丝氨酸/苏氨酸蛋白激酶2(LRRK2),和基因编码糖基神经酰胺酶(GBA1)的变异。携带LRRK2 G2019S突变(PD-LRRK2)的PD患者通常比散发性PD患者有更低的进展率,并且认知功能被保留。相比之下,携带GBA1变异(PD-GBA)的PD患者往往比散发性PD患者有更具侵袭性疾病,认知能力下降更快。事实上,PD-GBA的PDRP和PDCP网络表达水平高于散发性PD患者,尽管运动症状的持续时间和严重程度相似。PD-LRRK2患者中这两种模式的表达水平均低于PD-GBA患者。值得注意的是,与散发性PD患者相比,GBA1突变携带者在PDRP和PDCP表达水平之间表现出更大的差异。
图分析显示PD患者在PDRP和PDCP子网络的网络结构和信息流方面存在显著的基因型差异。在LRRK2 G2019S突变携带者中,代谢活性PDRP核心内的功能连通性增加,而在GBA1突变携带者中,在外周也观察到类似的变化。
(5)帕金森病的鉴别诊断
网络方法也可以帮助鉴别诊断帕金森运动障碍。帕金森病可与各种神经退行性病理相关,这些疾病在临床上在病程早期难以区分。在过去的12年里,多变量分析方法已经与疾病网络和脑成像扫描一起使用,以提高在研究和临床环境中的诊断准确性。例如,一种基于PDRP表达水平和与MSA和PSP(最常见的两种APSs)相关的疾病模式的两步逻辑回归算法已经被开发出来,用于个体患者预测诊断。MSA相关模式(MSARP)的特征是主要涉及壳核和小脑的代谢活动减少,而PSP相关模式(PSPRP)的特征是尾状核、前运动层和前额叶皮质、内侧丘脑和上脑干的代谢活动减少。MSARP和PSARP已在不同人群中得到广泛验证。虽然MSA和PSP可以分为临床刻板的亚类,但在这些亚类中,相应疾病模式的表达水平往往相似,尽管表型上存在显著差异。与皮质基底变性(CDB)相关的协方差模式(CBDRP)的特征是在额叶和顶叶区域、尾状核和丘脑的代谢活动的不对称的减少。与特发性帕金森病一样,APS相关的代谢模式在人群和临床表型中具有高度的可重复性。
由于疾病网络模式之间的部分重叠,可能需要几个网络来准确地分类单个患者。根据原始的三个网络诊断算法(图3),基于PDRP、MSARP和PSPRP表达水平的疾病分类依次进行:第一步鉴别特发性PD和APS,第二步鉴别MSA和PSP。该方法已在几组接受FDG PET治疗的帕金森病患者中得到验证,以解决最初不确定的临床诊断。一项荟萃分析显示,两步算法的合并敏感性为84%,其合并特异性为96%,这支持了其作为诊断测试的潜在用途。
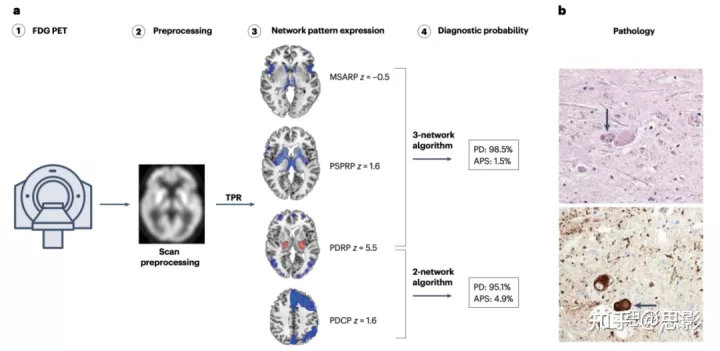
图3|帕金森病鉴别诊断:基于模式的分类算法。
a,52岁患者,对称性帕金森病,明显轴向僵直、低头受限和左旋多巴反应差,临床诊断为非典型帕金森综合征(APS)。患者在发病6年后接受了诊断性氟脱氧葡萄糖(FDG)PET治疗(步骤1)。扫描预处理后(第2步),使用TPR算法计算多系统萎缩相关模式(MSARP)、进行性核上麻痹相关模式(PSPRP)、帕金森病相关模式(PDRP)和帕金森病认知相关模式(PDCP)网络的表达水平(步骤3)。三个网络算法根据PDRP、MSARP和PSPRP表达对患者的扫描进行分类,表明特发性PD的概率高于APS(步骤4)。一个基于PDRP和PDCP表达的更简单的双网络算法对该个体产生了类似的分类结果(步骤4)。FDG PET后几个月的尸检显示特发性帕金森病的特征性表现:
b,上图,黑质色素神经元严重丢失,伴反应性胶质细胞增生,罕见保留神经元路易小体(箭头); 下图,免疫组化染色α-突触核蛋白阳性神经元路易小体(箭头)
PDRP和PDCP可能是已知疾病网络中验证最彻底的。尽管MSARP和PSPRP也得到了验证,但关于它们跨平台的稳定性较小。鉴于这一局限性,我们研究了一种简单的方法,即PDRP和PDCP一起使用于逻辑回归模型,以区分APS患者和特发性PD患者。如上所述,特发性PD患者始终表现出PDRP优势(即正向delta值),而这一特征通常在APS患者中不存在。事实上,一个简单的双网络模型基于PDRP和delta(数学上类似于PDRP和PDCP)大大提高了初始临床判断的准确性:AUC为0.85,尽管分类最准确的是使用原始三个网络模型(AUC为0.95)。尽管如此,考虑到fPDRP和fPDCP在重测评估中的重现性,双网络方法的良好准确性使它可以替代三个网络模型。
单基因神经退行性疾病
影响运动系统的单基因神经退行性疾病是罕见的。这些基因定义的疾病,包括亨廷顿病和脊髓小脑共济失调,其典型特征是具有相当一致的表型。临床前纵向神经成像研究揭示了从临床前或前驱阶段到明显疾病的表型转换的大脑变化。事实上,对无症状亨廷顿病突变携带者的纵向FDG PET扫描的OrT-CVA显示了一种称为亨廷顿疾病进展模式(HDPP)的代谢脑网络,该网络的表达水平在4年内持续增加。
脊髓小脑共济失调3型(SCA3)是一种罕见的单基因神经退行性疾病,与协方差模式相关,其特征是小脑、尾状核和后顶叶皮层的代谢活动减少,而躯体感觉皮层和边缘系统的代谢活动相对增加。
四、认知相关疾病
许多神经退行性疾病表现为认知能力下降,而不是运动症状。其中,最常见的是阿尔茨海默病、DLB和FTD。这些疾病都与特征性的病理改变有关,尽管它们在涉及的脑区,临床体征和症状有相当大的重叠。因此,就像帕金森病一样,对这些疾病的准确鉴别诊断是具有挑战性的,特别是对于处于认知障碍早期阶段的个体。事实上,25-30%的神经退行性认知障碍患者甚至在专科诊所也会被误诊。 研究疾病潜在的脑网络异常对于理解疾病机制和设计这些疾病的新治疗方法至关重要。正常认知相关网络及其随着年龄增长的变化,以及疾病相关的脑区连接改变,已被用来跟踪潜在的神经退行性过程的进展。
阿尔茨海默病网络
阿尔茨海默病的特征是淀粉样蛋白-β(Aβ)的异常积累和内侧颞叶tau蛋白的积累。Aβ和tau蛋白积累的阳性,目前可以通过脑脊液(CSF)分析或PET成像来证实(尽管血液生物标志物的发展正在迅速发展),这对于阿尔茨海默病的生物学诊断是必要的。潜在的病理学特征通常在第一个临床症状出现前数年就开始了,从主观认知能力下降到轻度认知障碍(MCI)到痴呆。阿尔茨海默病的病理对结构和功能脑连接有深远的影响。DMN功能最初出现变化,这些改变可能伴随着突显网络和边缘网络功能障碍或/和连接重构。
阿尔茨海默病相关模式(ADRP)最初是在H215O PET中发现的,并在随后的研究中证实了ADRP表达与认知能力下降之间的关系。从那时起,在不同的中心使用FDG PET和动态淀粉样PET扫描进行的研究,并发现了几种类似的ADRP模式。ADRP的特征是楔前叶和颞顶叶区域的代谢活动减少,这与小脑、桥脑和初级感觉运动皮层的代谢活动相对增加有关(图4a,上面板)。ADRP的表达水平升高在Aβ+MCI患者中和随后进展为痴呆的Aβ+ MCI患者中升高。事实上,这些值与整体认知功能的测量相关(图4a,底部面板),以及与注意力、记忆、执行功能和视觉空间能力测试的表现相关。MCI患者随后出现阿尔茨海默病(“转换”)与MCI患者没有发展阿尔茨海默病(“非转换”)相比产生的阿尔茨海默病转换相关模式(ADCRP)与之前描述的ADRP相似,而ADCRP的表达水平也与认知功能障碍的测量结果相关。也就是说,与临床和遗传生物标志物、脑脊液Aβ42与Aβ40比值、FDG PET的区域葡萄糖摄取测量和淀粉样PET成像结果相比,ADCRP表达水平更能预测随后的痴呆。作为未来认知能力下降的预测因子,ADCRP的表达优于脑脊液总tau蛋白和血浆神经丝轻链水平。有趣的是,ADCRP的表达与神经原纤维tau蛋白的Braak期之间存在中度相关性。
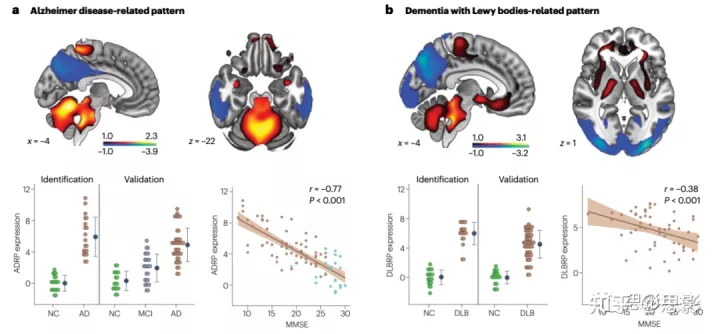
图4|与阿尔茨海默病和路易小体痴呆相关的异常代谢协方差模式。
a,顶部面板:阿尔茨海默病(AD)相关模式(ADRP)的特征是楔前叶和颞顶叶区域的代谢活动减少(蓝色),与小脑、脑桥和初级感觉运动皮层的相对增加(红色)相关。
底部面板:与健康对照个体(NC)相比,在淀粉样蛋白β阳性轻度认知障碍(MCI)患者和痴呆患者ADRP表达水平升高。
b,顶部面板:路易体痴呆(DLB)相关代谢协方差模式(DLBRP)的特点是在楔前叶和枕叶、顶叶下和颞下区域的活动减少(蓝色),壳核、杏仁核、海马、海马旁回和小脑蚓部的活动相对增加(红色)。
底部面板:DLB患者的DLBRP表达水平高于健康对照组患者。ADRP和DLBRP的表达均与迷你精神状态检查(MMSE)评分相关
其他痴呆网络与路易体痴呆
路易体痴呆(DLB)
DLB是继阿尔茨海默病之后的第二种最常见的神经退行性认知障碍。DLB与皮质和皮层下脑区α-突触核蛋白异常积累相关,临床表现为痴呆伴有帕金森病、视觉幻觉、认知波动和/或RBD。DLB相关代谢协方差模式(DLBRP)的特征是枕叶、顶叶下、颞下区和楔前叶的活动减少,与壳核、杏仁核、海马、海马旁回和小脑蚓部的活动相对增加有关(图4b,上面板)。在几个独立患者队列中,已经发现了与相似DLBRP的网络模式。这些研究中的DLBRP表达水平与个体在成像时的认知表现相关(图4b,底部面板)或与生存时间相关。
值得注意的是,多项研究报道了非阿尔茨海默病痴呆患者中ADRP表达增加。事实上,仅根据ADRP就不能将DLB患者与阿尔茨海默病患者区分开来。也就是说,对于单一的疾病相关模式,不一定能实现特定个体的准确疾病分类。一些研究已经调查了交叉模式在鉴别诊断中的应用。这些模式旨在实现两种临床相似的疾病状态之间的最大区分。交叉模式和其他分析策略在区分DLB患者和阿尔茨海默病患者方面取得了令人鼓舞的结果。
额颞叶痴呆(FTD)
神经退行性痴呆的第三个最常见的原因是FTD。这种疾病又分为三种临床亚型:行为变异性FTD(bvFTD)、非流利变异性原发性进行性失语症和语义变异性原发性进行性失语症。这些变异中最常见的是bvFTD,它与人格和行为的变化有关。一种bvFTD相关模式(bvFTDRP)已在三个独立的患者群体和健康对照个体中被确认,并在一个神经病理学研究中证实。这种代谢协方差网络(图5a)的特征是额叶和颞区、前扣带回和中扣带回、脑岛叶和丘脑的活动减少,这些与枕叶皮层和小脑的相对活动增加有关。这种模式的表达水平(图5b)在bvFTD患者中的表达水平始终高于健康对照组个体。
有趣的是,我们发现bvFTD与正常的DMN重叠,在对bvFTD患者的多个研究中,DMN模式的表达持续减少。这一情况表明,DMN是bvFTD的病理过程的一个重要靶点。事实上,由三个独立的患者样本构建的bvFTDRP图的可视化显示了DMN模块的远程连接的变化(图5c)。因此,DMN的碎片化可能是bvFTD患者认知功能障碍的一个重要机制。
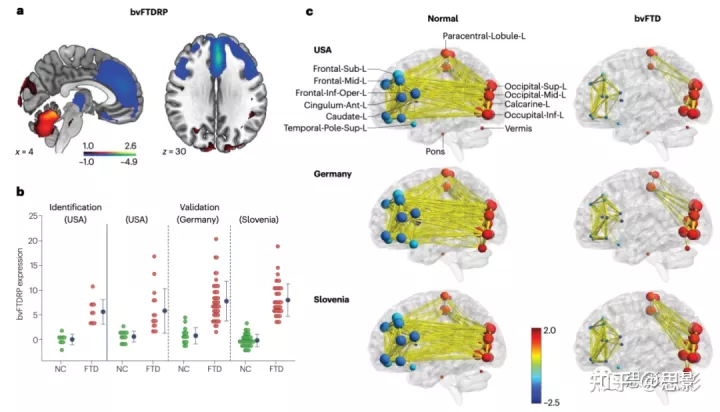
图5|与额颞叶痴呆相关的异常代谢协方差模式。
a,行为变异性额颞叶痴呆(bvFTD)相关模式(bvFTDRP)的特征是额颞叶区域、前扣带回和中扣带回、脑岛叶和丘脑的活动减少(蓝色),枕叶和小脑相对增加(红色)。
b,在用于模式识别的衍生样本(美国)以及用于验证的独立测试样本(美国、德国和斯洛文尼亚)中,bvFTD患者的表达水平升高。
c,网络空间的功能连接,对美国、德国和斯洛文尼亚的bvFTD患者和健康对照个体的扫描样本。为清晰起见,只标记了左侧的节点;关于所有相关节点的详细信息在其他地方提供。图形显示了三个位置相似的连接模式,在所有患者群体中,前后皮质模块断开。边的粗细对应于相关性的大小。给出了所有r>为0.60的边。bvFTDRP节点根据其相应区域权重的符号进行颜色编码;直径与相应的中心性度量成正比。
Ant,前;frontal-inf-oper,额下回盖部;L,左半球;inf,下;sup,上。
总结
在独立的临床中心和成像平台上,对功能性脑网络模式的广泛验证及其在功能磁共振成像中的新兴应用,为这种方法在研究和临床决策中的广泛应用奠定了基础。数据共享的增加和开放获取的数据库数量的增加可能会加速这一趋势,但将需要制定和实施严格的数据协调程序。功能性脑网络成像已经可以促进早期诊断,并帮助监测疾病进展和检测治疗对个别患者的效果。

















 893
893

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









