





本文介绍了一款新的软件包,其中包含了一系列标准化的神经纤维束追踪协议,这些协议旨在稳健地自动提取人类和猕猴大脑中的白质纤维束。利用人类连接组项目(HCP)和英国生物银行的数据,以及猕猴大脑数据集的体外数据,我们获得了两个物种的白质图谱,以及白质和灰质边界上的纤维束终点图谱。我们展示了我们的协议对数据质量具有鲁棒性,能够跨两个物种通用,并且反映了已知的解剖结构。我们进一步证明了这些协议能够捕捉到个体间的变异性,通过保留人类纤维束的侧化特征和HCP队列中双胞胎间纤维束的相似性。我们的结果表明,这个工具箱将有助于在大型队列中生成成像衍生特征,并促进比较神经解剖学研究。该软件、神经纤维束追踪协议和图谱通过FSL公开发布,允许用户以标准化的方式定义自己的神经纤维束追踪协议,进一步促进开放科学。本文发表在NeuroImage杂志。(可添加微信号19962074063或18983979082获取原文及补充材料,另思影提供免费文献下载服务,如需要也可添加此微信号入群,另思影脑影像分析业务,如感兴趣也可咨询)
研究亮点:
• 开发了一款新的软件包,用于标准化和自动化跨物种的神经纤维束追踪技术。
• 在人类和猕猴大脑中,发现了相似的白质纤维束。
• 利用大型数据集(1000名受试者)生成了人类白质纤维束图谱。
• 神经纤维束追踪协议已标准化,但仍保留了个体间的变异性。
• 使用人类连接组项目(HCP)和英国生物银行数据,展示了该技术在不同数据集中的普适性。
关键词:扩散MRI、白质纤维束、白质图谱、非人灵长类动物、人类连接组项目、比较解剖学
1.引言
扩散纤维束追踪是一种独特的工具,用于非侵入性地、在体内提取白质(WM)通路。主要WM纤维束的虚拟解剖使得研究大脑组织成为可能(Catani等,2013;Jbabdi等,2015),并为大脑发育(Huppi和Dubois,2006)和WM病理(Ciccarelli等,2008;Griffa等,2013)提供了探针。它还允许探索个体差异(Assaf等,2017)和跨物种的解剖和连接性差异(Mars等,2018b)。这一信息具有功能相关性,因为每个功能性大脑亚单位到大脑其余部分的外在白质连接模式是独特的。(Mars等,2018a;Passingham等,2002)。
为了可靠地研究WM通路中的个体变异,纤维束追踪方法通常使用协议来提取预定义的一组WM纤维束。这些协议通常包括感兴趣区域(ROI)及其使用规则(例如作为种子、排除和终止掩模)。它们反映了用于指导和约束曲线传播的先前解剖学知识,减少了假阳性的可能性(Catani等,2002;Wakana等,2004)。纤维束追踪协议必须是鲁棒和可重复的,允许在尊重潜在的解剖变异和个体差异的同时,以一致的方式在受试者之间重建WM纤维束。一种可能使用的方法是定义特定于受试者的纤维束追踪协议(Conturo等,1996),考虑到个体解剖的特定变化。然而,基于受试者的掩模定义既耗时又主观(Jones,2008;Nucifora等,2012),而对于大型队列来说,这些限制变得难以克服。这种手动方法的替代方案是在模板空间中定义一组标准化的掩模,然后将其配准到个体几何结构中,并以一致和自动化的方式用于每个受试者。
这些基于自动化感兴趣区(ROI)的纤维束追踪方法在提取多种纤维束方面已被证明是强大的(Catani等,2012;Eichert等,2019b;Hau等,2016;Hecht等,2015;Howells等,2018;Maffei等,2019;Makris等,2013, 2009;Menjot de Champfleur等,2013;Nowell等,2016;Takemura等,2017;Thiebaut de Schotten等,2011a;Zhao等,2016)。它们进一步允许应用于大型队列,以发展特定纤维束的图谱(Archer等,2018;Chenot等,2019)和提取特定纤维束的特征(Miller等,2016)。
在本文中,我们介绍了一款新的软件包,其中包含了一系列标准化的纤维束追踪协议,这些协议旨在自动提取人类和非人灵长类动物大脑中的白质纤维束。我们将人类和猕猴大脑的先前解剖学知识转化为一组等效的纤维束追踪规则,以获得多种白质束的同源重建。这种等效性为比较神经解剖学和在不同物种间识别功能上等效的皮层区域提供了独特的可能性(Mars等,2018b)。
这项工作建立在以前努力的基础上,并扩展了在人类大脑中开发纤维束追踪协议库的工作(Catani和Thiebaut de Schotten,2008;de Groot等,2013;Hua等,2008;Thiebaut de Schotten等,2011b;Wakana等,2007;Wassermann等,2016;Yendiki等,2011;Zhang等,2008)(参见补充表1以获取摘要)。
Wakana等(2007)开发了一组标准空间掩模,用于提取20种纤维束。他们报告了高度的评估者间和评估者内的重复性,并提出一些纤维束可能显示左右不对称。Hua等(2008)扩展了这项工作,生成了22种纤维束(11左/右)的概率性图谱,并通过研究多发性硬化症中的特定纤维束异常来示范其用途。Zhang等(2008)将这些标准化掩模应用于10名受试者,并报告了自动化和手动纤维束分割方法之间的高度一致性。同样,Catani和Thiebaut de Schotten(2008)定义了用于重建19种纤维束(7左/右,5交叉)的标准空间掩模,并评估了他们协议的重复性。他们的工作通过Thiebaut de Schotten等(2011b)进一步扩展,将纤维束追踪协议扩展到31种纤维束(14左/右,3交叉),并报告了他们的自动化纤维束追踪技术与组织学图谱之间的良好对应关系。Yendiki等(2011)引入了一种全局纤维束追踪方法,基于图谱导出的先验来重建18种纤维束(8左/右,2交叉)。对于每种纤维束,他们在训练队列中进行了手动纤维束标记,这为自动化纤维束追踪提供了先验信息,以用于贝叶斯概率框架中。de Groot等(2013)使用标准化的纤维束追踪协议在两个不同质量的数据集中重建了27种纤维束(12左/右,3交叉)。最近,Wassermann等人(2016)提出了一个描述白质解剖和纤维束的框架,该框架使用受试者特定的解剖分割、聚类和查询语言,从整个大脑的纤维束追踪和灰质(GM)分区中提取57个纤维束(25个左/右,7个交叉)。他们的方法将纤维束的定义简化为一组逻辑规则,这些规则参考了纤维束的位置和终止、纤维束的路径及其与给定大脑区域的空间关系。
尽管之前的发展具有巨大潜力,但没有一个针对多物种,这正是我们研究的目标。我们的方法也是对非ROI(感兴趣区域)基础的白质纤维束重建方法的补充,例如无监督的基于聚类的方法(例如Garyfallidis等,2012;O’Donnell和Westin,2007;Siless等,2018)或监督方法(Wasserthal等,2018)。前者是“数据驱动”的,而我们则强加先前的解剖学知识以减少纤维束追踪中的假阳性。后者在训练期间可以从新方法中受益,例如这里介绍的旨在以一致和可重复的方式重建已知解剖学纤维束的方法。
总之,本研究设计了一套扩展的基于ROI的纤维束追踪协议,适用于人类和猕猴大脑,并将这些协议捆绑在一个自动化纤维束追踪工具箱中。本文工作的贡献如下:
1)我们为42种纤维束设计了纤维束追踪协议,并利用来自人类连接组项目(HCP)(Sotiropoulos等,2013)的高分辨率数据和来自英国生物银行(Miller等,2016)的更典型数据来展示它们对数据质量的鲁棒性;
2)我们展示了纤维束追踪协议对猕猴大脑的普适性;
3)我们使用这些协议为人类大脑(1000名HCP受试者)和猕猴大脑(6只动物的高分辨率体外数据集)派生出高质量的纤维束图谱;
4)我们通过评估提取的纤维束的侧化(在人类中)进行间接验证;
5)我们展示了,尽管是模板驱动的,重建的纤维束仍保留了个体变异性,这是通过双胞胎分析评估的;
6)我们提供了一个开源灵活的框架,用于在FSL(Jenkinson等人,2012)内公开交换纤维束追踪协议。可以定义新的标准空间白质纤维束追踪协议,并“插入”到工具箱中,允许进一步扩展和纤维束交换,有助于开放科学和结果的可重复性。
2.纤维束追踪协议定义
我们为42种白质(WM)纤维束(19种双侧和4种交叉)设计了纤维束追踪协议,这些协议以一种普适的方式制定,允许等效的掩模定义适用于人类和猕猴大脑。目前支持的纤维束完整列表在表1中呈现。我们进一步将它们打包进一个新的跨物种纤维束追踪(XTRACT)工具箱中,这是一个包装器,能够读取标准空间纤维束追踪协议,并使用FSL的probtrackx2(Behrens等,2007)执行概率性纤维束追踪,同时提供GPU加速选项(Hernandez-Fernandez等,2019)。
表1. 重建的WM纤维束列表、它们对应的纤维束类型以及使用的播种策略。
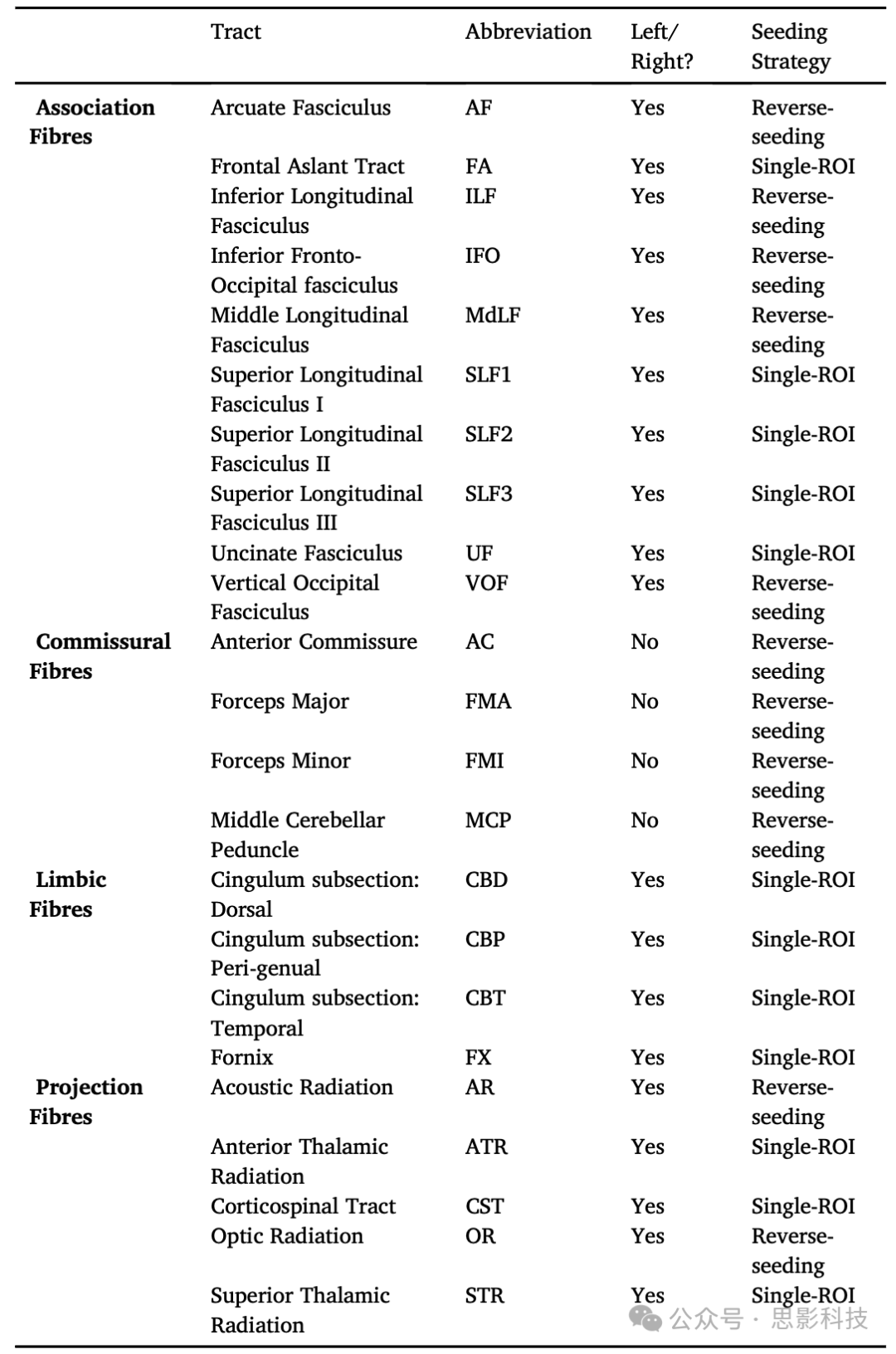
图1展示了单个纤维束追踪协议的主要阶段。每个纤维束使用在标准空间(人类为MNI152,猕猴为F99)定义的独特掩模组合进行重建。这些协议由多位专家(RM、KB、GD、SW、SNS)共同开发和认可的手绘和基于图谱的掩模组成。这些掩模包括种子(纤维束追踪流线的起始点)、目标/途径点(流线必须通过的区域才被视为有效)、排除区(用于排除任何穿过它们的流线)和停止/终止掩模(用于停止任何穿过它们的流线)。播种策略支持:a) 标准的单ROI种子区和b) “反向播种”方法,其中一对种子-目标的mask互换角色,最终将通路相加求和。该协议的masks通过非线性配准形变场从标准空间转换到受试者的个体空间。纤维束追踪在个体空间下进行,结果直接重采样到标准空间,允许受试者间的几何对应,这在某些情况下是必要的(例如,制作图谱)。
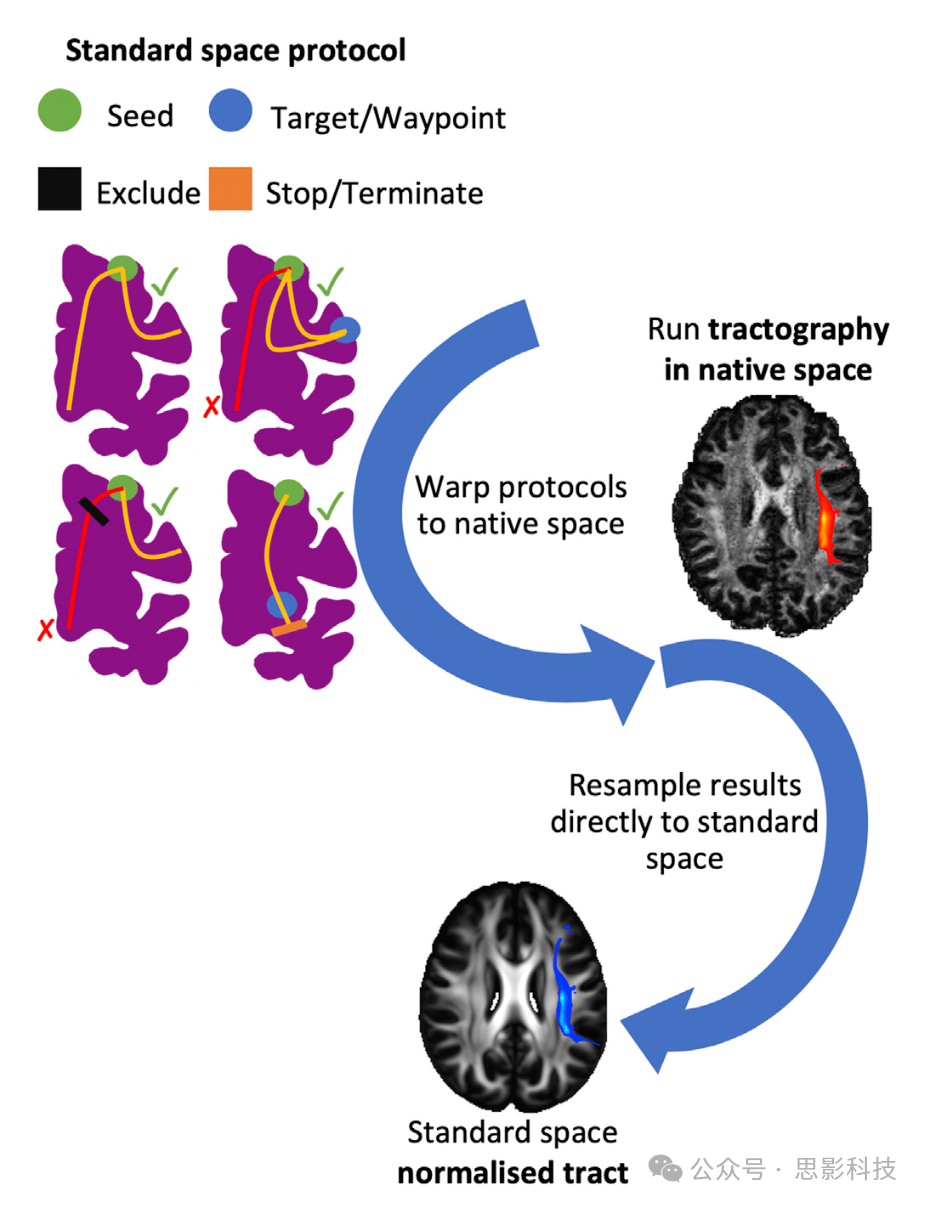
图1. 自动化纤维束追踪的阶段示意图,如“XTRACT”工具箱中实现的,以及人类大脑左侧弓状束(AF)的示例。
1)纤维束追踪协议掩模在标准空间中定义,包括种子(绿色)、排除(黑色)、途径点(蓝色)和终止(橙色)掩模(有关定义的完整细节,请参见“协议”部分)。
2)协议掩模使用受试者特定的非线性形变场变形到受试者的个体空间。
3)在受试者的个体空间中使用交叉纤维模型的扩散数据进行概率性纤维束追踪。请注意,纤维束追踪是在个体扩散空间的连续坐标中进行的。插值仅在保存结果为空间直方图(即纤维束访问图)时发生。对于这一步骤,使用最近邻插值,空间分辨率由用户指定。
4)结果存储在标准空间的纤维束,叠加在FSL_HCP1065 FA图谱上。
下面的部分详细描述了在人类纤维束追踪中考虑的每个纤维束的协议,以及对猕猴大脑的任何调整。除了脑干和交叉纤维束之外,所有协议都包括中线矢状平面作为排除掩模,以限制纤维束到同侧半球。
2.1. 联合纤维
上纵束(SLF)1/2/3:上纵束的三个分支是使用(Thiebaut de Schotten等,2011a)采用的方法的扩展来重建的。在每种情况下,额叶/顶叶皮层中央沟区域的冠状面和两个目标mask都被用作种子。在额部,SLF的第一、第二和第三分支的目标mask分别是通过额上、中和下回的冠状切面,放置在胼胝体膝后端的水平上。在后部,一个位于顶叶上部紧靠扣带回边缘后方的较大冠状目标mask用于SLF1。对于SLF2和SLF3,第二个目标mask分别放置在角回和缘上回(SMG)。在每种情况下,种子的位置反映了第二个目标的位置,同时向前移动到中央沟区域。对于每个协议,一个轴向排除mask放置在顶叶皮层下方,一个阻止皮层下区域的mask用于防止追踪到腹侧走向的纤维束中。最后一个通过胼胝体膝尾端后方的皮层下区域的冠状排除mask防止追踪到腹侧纵向纤维束中。
弓状束(AF):弓状束是背侧纵向纤维系统的一部分,但在人脑中以其后部曲线向腹侧延伸至颞皮层而有所区别。重建人类 AF 时,种子位于额旁回(SMG),颞叶目标mask位于包围颞上回(STG)和颞中回(MTG)的白质中,前方目标区域位于腹侧运动前皮层水平、额下回(IFG)后部和前中央沟前部。根据对猕猴的观察,这条纤维束沿着环状岛沟的底部运行(Petrides 等人,2012 年),我们在中央沟后部放置了一个种子掩码。在Sylvian 裂尾端后方的顶-颞-白质中放置了一个轴向目标掩模。在IFG中放置了一个额外的轴向平面。这个协议由(Eichert等,2019b)验证。
中/下纵束(MdLF,ILF):重建了沿着颞叶分布的三条纤维束(MdLF,ILF,IFO)。中和下纵束保持在外侧后皮层内。MdLF在上额叶回(SFG)的前部播种(Makris等,2009);ILF在颞中回和颞下回播种,以考虑人类大脑相对于猕猴颞叶皮层的扩张(Latini等,2017;Roumazeilles等,提交)。对于MdLF,基于(Makris等,2013)的解剖描述,使用覆盖颞-顶-枕交界处白质的大轴向和冠状平面作为目标区域。对于ILF,使用颞中回和颞下回的冠状平面作为目标。对于这两个协议,排除mask的轴向穿过脑干,冠向穿过穹窿,轴向穿过胼胝体后方的纵束,并穿过整个额叶皮层。此外,MdLF 的种子mask可用作 ILF 的排除mask,反之亦然,ILF 的目标mask可用作 MdLF 的排除mask。此外,对于 ILF,在半卵圆中心放置了冠状排除mask,并使用了覆盖 SMG白质的轴向排除mask。
额下枕束(IFO):与MdLF和ILF不同, 额下枕束,也称为最外囊纤维复合体(Mars等,2016),更靠近中线,并通过最外囊进入额叶皮层。扩展(Wakana等,2007)的方法,种子是通过枕叶皮层前部的冠状面,目标是通过胼胝体膝部前侧的额叶皮层的冠状面。位于前交叉后方的排除mask排除了所有纤维,但穿过最外囊的纤维除外。
钩状束(UF):最外囊的底部包含属于钩状束的纤维,这些纤维从下额叶皮层弯曲到前颞叶皮层。该纤维束通过在STG放置种子(在颞叶和额叶皮层首次分离的位置),通过最外囊的腹侧部分放置目标,以及在种子和目标之间放置排除mask层以强制曲线形成来重建。另一个冠状排除mask防止意外泄漏追踪到沿颞叶纵向穿过的纤维中。
额斜束(FA):额斜束是位于额叶下回和额上回后部之间的一条短束(Catani等,2012)。种子放置在IFG的白质中的矢状位,目标放置在SFG的白质中的轴向位。一个后部冠状排除mask防止追踪到纵向纤维中。
垂直枕束(VOF):垂直枕纤维束(VOF)主要以背-腹方向在枕叶内运行。我们使用了(Takemura等,2017)描述的方法的改编版本。一个轴向种子mask放置在枕叶腹侧白质的外侧部分,位于前枕沟后方(Petrides等,2012)。一个更大的轴向目标mask放置在外侧枕沟水平的背侧。一个位于胼胝体后方的冠状平面作为排除mask,以防止追踪到前-后纤维束中。
2.2. 交叉纤维
中小脑脚(MCP):中小脑脚(MCP)在小脑白质中播种,目标位于对侧半球(及其相反方向)。排除mask沿小脑中线矢状放置,通过丘脑轴向放置。
胼胝体压部(FMA)和膝部(FMI):我们根据(Wakana等,2007)定义的方法,重建了通过胼胝体压部(大脑钳,FMA)到枕叶的胼胝体连接,以及通过胼胝体膝部(小脑钳,FMI)到额叶的连接。FMA的种子和目标mask(及其相反方向)定义为通过枕叶的冠状切面,在顶-枕沟的后端。矢状排除掩模限制在枕皮层和皮层。通过下额-枕白质和桥脑的冠状平面的额外排除掩模防止追踪到纵向纤维。FMI的种子和目标mask(及其相反方向)定义为通过额叶的冠状切面,在前扣带回沟的前端。中矢状排除mask在胼胝体前三分之一的水平中断,同一水平的额外冠状排除mask防止向后投射。
前交叉(AC):前交叉连接两个半球的颞叶,穿过中线。它在中线上的左右定向纤维中播种,目标掩模覆盖了苍白球外侧的白质。停止mask直接放置在两个杏仁核的下方和侧面。一个大的轴向排除mask放置在种子上方的整个皮层背侧。
2.3. 边缘系统纤维
扣带回纤维束子部分(CBT, CBP, CBD):最近(Heilbronner和Haber,2014)提出了一种基于连接特定扣带回、非扣带回额叶和皮层下目标的纤维存在情况,将扣带回纤维束分割成不同部分的方法。因此,我们为扣带回纤维束的三个不同子部分创建了协议。颞叶部(CBT)在颞叶的后部播种,在该部位扣带回纤维束主要呈前-后方向。目标放置在杏仁核后方,种子和目标mask的后方和前方分别放置停止mask。一个排除mask防止追踪到穹窿。背侧段(CBD)在胼胝体后部上方播种,目标位于胼胝体膝的起始处。内囊前肢的矢状排除mask防止追踪到颞叶。最后,扣带回纤维束的围膝部分(CBP)在胼胝体上方前部播种,目标放置在胼胝体下方,目标下方和前方放置停止mask。在 Sylvian裂的前端水平的胼胝体平面防止追踪到CBD。
穹窿(FX):室旁体连接海马体、乳头体、前丘脑核和下丘脑(Catani等,2013)。该纤维束通过在胼胝体中部水平的穹窿体部播种和在海马体中放置目标来重建。在枕叶皮层前端的胼胝体平面防止追踪到后部纤维束,丘脑前端尖端水平的中线周围的双侧矢状平面防止向内囊前肢的侧向传播。为了防止追踪到扣带回,胼胝体压部后方的轴向排除mask和覆盖扣带回海马回区域的小轴向排除mask也被使用。我们应该指出,由于纹状体终端的相对较小尺寸和它与穹窿的接近,穹窿追踪可能追踪到纹状体终端。这是扩散纤维束追踪中的一个常见问题,目前尚未使用与本研究中使用的方法克服(Kamali等,2015;Mori等,2017;Mori和Aggarwal,2014;Pascalau等,2018)。
2.4. 投射纤维
皮质脊髓束(CST):皮质脊髓束,或锥体束,从脊髓延伸至中脑,并分布至运动皮层、前运动皮层和体感皮层。该纤维束从桥脑播种,目标覆盖运动、前运动和体感皮层。使用轴向排除掩模限制追踪到大中脑的脑脚。此外,排除掩模包括两个冠状平面,分别位于目标的前方和后方,以排除追踪到前额皮层和枕皮层,以及一个防止泄漏到小脑脚的平面。
丘脑前辐射和上丘脑辐射(ATR、STR):丘脑前辐射和上丘脑辐射分别将丘脑连接到额叶和前/后中央回。前部丘脑辐射使用穿过丘脑前部的冠状掩模播种(Wakana等,2007),目标冠状掩模位于前丘脑脚。此外,排除掩模包含一个覆盖中脑基底的轴向平面,一个防止通过后丘脑脚泄漏的冠状平面,以及一个防止通过扣带回泄漏的冠状平面。一个覆盖丘脑后部的冠状停止掩模,从中脑基底延伸到胼胝体沟。上部丘脑辐射使用覆盖整个丘脑的掩模播种,目标轴向平面覆盖上丘脑脚。一个轴向平面作为停止掩模位于丘脑腹侧。排除掩模包括两个冠状平面,分别位于目标的前方和后方,以排除追踪到前额皮层和枕皮层。
声辐射(AR):声辐射将丘脑的内侧膝状体(MGN)连接到听觉皮层。它从横向颞回播种,目标覆盖丘脑的MGN。排除掩模包括两个冠状平面,分别位于丘脑的前方和后方,以及一个位于丘脑上方的轴向平面。此外,排除掩模还包括脑干和覆盖视神经束的水平区域。
视辐射(OR):视辐射由丘脑外侧膝状体(LGN)到初级视觉皮层的纤维组成。它在LGN播种,目标掩模由穿过距状裂前部的冠状平面组成。排除掩模包括脑干的轴向阻断、直接位于LGN后方的纤维的冠状阻断(用于选择向背侧卷曲的纤维),以及种子前方的冠状平面,以防止泄漏到纵向纤维。
2.5. 猕猴大脑的调整
尽管上述协议允许在猕猴大脑中进行等效定义,但为了确保解剖学的准确性,一些调整是必需的。对于所有猕猴协议,使用了反向播种方法,因为这被发现可以增加所得纤维束的鲁棒性。此外,对AF和MdLF协议进行了调整以反映猕猴大脑的特点。在AF的情况下,种子放置在STG的尾部,目标直接位于主沟上方,延伸到8Ad的后方(基于Schmahmann和Pandya,2006的示踪数据)。此外,一个放置在STG尾部的目标,位于种子的下方和后方,确保了追踪通过侧裂的尾部进行。对于MdLF,STG后部的单个轴向平面被用作目标。
3.材料和方法
3.1. 数据和预处理
为了评估不同数据质量下的鲁棒性,我们使用了来自人类连接组项目(HCP,尖端扩散MRI)(Sotiropoulos等,2013;Van Essen等,2013)的数据和来自英国生物银行(Miller等,2016)的数据(总体质量更接近临床扫描仪通常可获得的质量)。为了确保协议跨物种的普适性,我们还使用了猕猴大脑的扩散MRI数据。这些数据包括在(Eichert等,2019a;Mars等,2018b)中使用的一组扩展的动物。总体而言,我们考虑的数据集包括来自HCP的1065名受试者(所有可用的HCP S1200受试者,他们有扩散MRI数据),来自英国生物银行的1000名受试者,以及来自6只猕猴的体外高分辨率数据集。对于HCP数据,我们从统计比较和群体图谱中移除了44名被识别出有解剖异常的受试者(详情见HCP质量控制网站),使我们总共有1021名受试者,来自有扩散数据的1065名受试者。
对于HCP和英国生物银行的数据,我们使用了各自公开发布的预处理扩散MRI数据(详细描述见Glasser等,2013;Sotiropoulos等,2013和Alfaro-Almagro等,2018;Miller等,2016)。简而言之,HCP数据是在定制的3T Connectom Skyra(西门子,埃尔朗根)上获取的,使用单极扩散加权(Stejskal-Tanner)自旋回波EPI序列,等向空间分辨率为1.25毫米,三个壳层(b值=1000、2000和3000 s/mm²)和每个壳层90个独特的扩散方向,每个受试者扫描两次(总扫描时间约60分钟)。英国生物银行的数据是在临床3T Skyra(西门子,埃尔朗根)上获取的,包括两个壳层(b值=1000和2000 s/mm²)和每个壳层50个扩散方向,等向空间分辨率为2毫米(每个受试者总扫描时间约6分钟)。在这两种情况下,数据都经过了运动、磁敏感畸变和涡流畸变校正(Andersson等,2003;Andersson和Sotiropoulos,2016)。使用各自的T1加权图像和FSL的FNIRT(Andersson等,2007;Jenkinson等,2012),获得了到标准空间(MNI152)的非线性变换,形变校正后的扩散MRI数据也进行了线性配准。扩散到T1和T1到MNI变换的连接允许获得扩散到MNI的形变场。
对于猕猴数据,我们结合了之前在(Mars等,2018b)中使用的数据和新获取的数据。数据是在7T磁体上使用Agilent DirectDrive控制台(Agilent Technologies,圣克拉拉,加州,美国)获取的,使用2D扩散加权自旋回波协议和单线读取(DW-SEMS,TE/TR:25毫秒/10秒;矩阵大小:128×128;分辨率:0.6×0.6毫米;切片数:128;切片厚度:0.6毫米;扩散数据在53小时内获取)。获取了16个非扩散加权(b=0 s/mm²)和128个扩散加权(b=4000 s/mm²)体积,扩散方向分布在整个球面上。扫描前,大脑浸泡在PBS中,并在扫描过程中放置在fomblin或fluorinert中。这些数据将通过PRIME-DE(Milham等,2018)提供。
使用FSL的FNIRT(Andersson等,2007;Jenkinson等,2012),基于分数各向异性(FA)图,获得了到标准空间(F99)(Van Essen,2002)的非线性变换估计。
3.2. 纤维方向估计和纤维束追踪
在“XTRACT”之前,使用FSL的bedpostx将(Jbabdi等,2012)描述的交叉纤维模型应用于扩散数据,用于估计纤维方向以指导纤维束追踪。这是一个参数化的球面解卷积模型,考虑了随着更高b值的dMRI信号的非单指数衰减。在每个体素中估计了多达三个纤维方向及其不确定性。XTRACT工具箱读取了标准空间纤维束追踪协议,并执行了概率性纤维束追踪(Behrens等,2007)。如前所述,每个束的纤维束追踪协议使用种子、目标、排除和停止掩模的独特组合以及播种策略来定义(见表1)。所有协议中还使用了一些默认的纤维束追踪终止标准(曲率阈值:±80°,最大流线步数:2000,次要纤维体积阈值:1%,在种子位置存在纤维交叉时随机采样初始纤维,无最小长度限制,循环检查和终止)。人类和猕猴纤维束追踪分别使用了0.5毫米和0.2毫米的步长。如图1所示,掩模被变形到受试者的本地空间,纤维束追踪后,纤维束追踪结果直接重采样到标准空间。所得分布相对于生成的有效流线总数进行了归一化(即未被包含/排除掩模标准拒绝的流线)。
为了获得纤维束图谱,以人群百分比重叠的形式,我们将每个归一化路径分布在阈值处二值化。然后对二值掩模进行队列平均,以得出在给定体素中给定纤维束存在的受试者百分比。
3.3. 连接蓝图
使用连接蓝图(Mars等人,2018b)进一步利用估计的纤维束来估计每个纤维束的“皮层终止”图。具体来说,为每个受试者重建了一个白-灰质边界(WGB)乘以纤维束的矩阵CB,如图2所示。这是通过从每个白-灰质边界位置播种并计算访问整个白质(WM)的次数来实现的,从而得到一个WGB乘以WM的连接C1矩阵。使用纤维束追踪协议获得的纤维束被向量化并连接成一个单一的WM乘以纤维束的C2矩阵。两个矩阵的乘积提供了一个连接“蓝图”,即CB等于C1xC2(WGB乘以纤维束)矩阵。该矩阵的列代表相应纤维束在白-灰质边界表面的终止点,而行显示了每个皮层位置的连接模式(即每个纤维束如何贡献于每个皮层位置的整体连接)。这个过程对HCP受试者和猕猴数据集进行了执行。然后,结果进行了队列平均,以产生连接蓝图图谱。
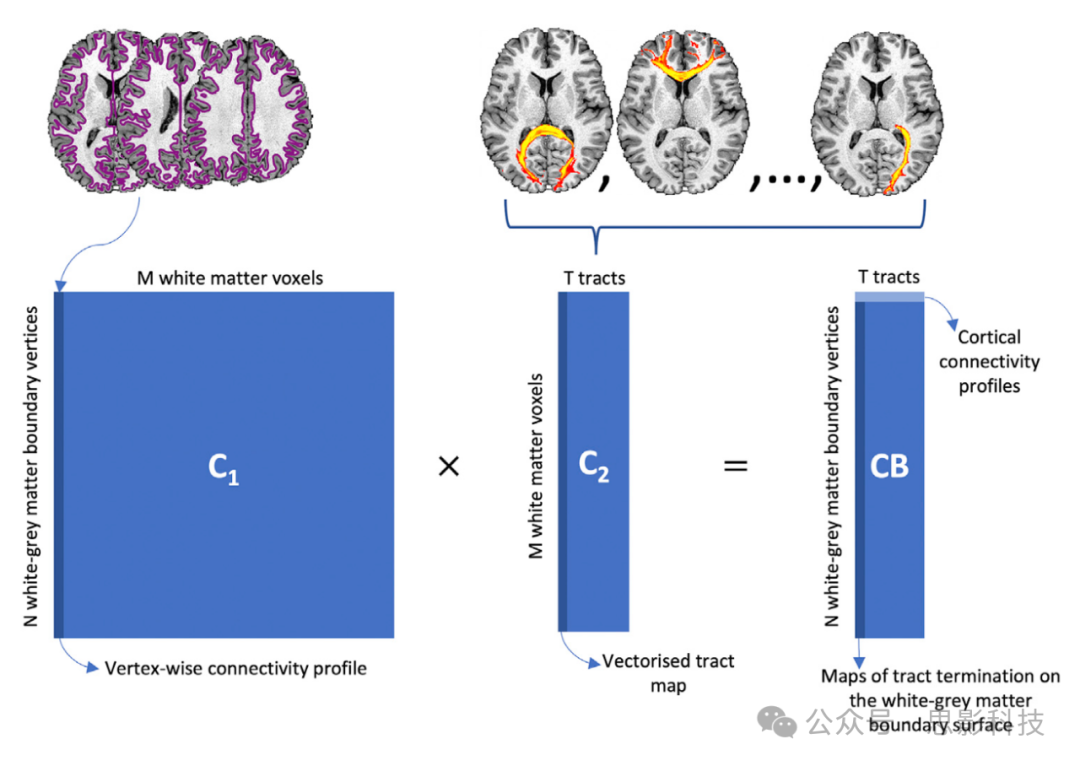
图2. 连接蓝图构建的示意图。
从白-灰质边界(由紫色轮廓表示)播种纤维束追踪,然后计算访问整个白质(WM)的次数,得到C1矩阵(N约60k乘以M约58k)。该矩阵的列代表顶点级的连接性概况。接下来,使用XTRACT产生的纤维束追踪重建被向量化并堆叠,形成一个WM乘以纤维束的矩阵,C2(M乘以T=41)。两个矩阵的乘积得到连接蓝图,CB(N乘以T)。列代表纤维束在白-灰质边界表面的终止图;行代表白-灰质边界连接概况,反映了每个纤维束对每个白-灰质边界顶点连接模式的贡献。
3.4. 评估纤维束侧化
为了证明我们的协议是否产生了符合解剖学预期的纤维束,我们使用大量受试者研究纤维束侧化。根据文献,预期AF是左侧化的(Eichert等,2019b;Nowell等,2016;O’Donnell等,2010;Panesar等,2018;Propper等,2010),IFO、MdLF和SLF3是右侧化的(Hau等,2016;Howells等,2018;Menjot de Champfleur等,2013;Thiebaut de Schotten等,2011a),而SLF1预期是非侧化的(Thiebaut de Schotten等,2011a)。文献表明SLF2是右侧化的,但发现不太确定,因为在某些情况下报告的侧化没有达到显著性(Hecht等,2015;Thiebaut de Schotten等,2011a)。
我们使用纤维束体积评估了纤维束侧化。具体来说,侧化(L)是相对右-左体积(Vr和Vl)差异的计算,通过将归一化纤维束在0.5%处二值化并计算体素数,即L = (Vr - Vl)/(Vr + Vl),与计算纤维束侧化的文献一致(O’Donnell等,2010;Propper等,2010;Thiebaut de Schotten等,2011b)。
此外,我们探索了皮层终点图中的两半球间差异。我们获得了只包括左半球纤维束/列的连接蓝图CBL,以及只包括右半球纤维束/列的CBR。两个矩阵都进行了行归一化,使得每行中所有元素的总和等于1。随后,我们计算了每对(CBR,CBL)行之间的Kullback-Leibler(KL)散度(一种不相似度量)。我们使用在所有可能对中获得的最小KL散度值评估了每个半球位置i中右-左连接模式的相似性,即min(CBRi, CBLj),其中j跨越所有白-灰质边界位置。
3.5. 尊重源自双胞胎身份的相似性
虽然我们的目标是使自动化纤维束追踪协议对数据质量具有鲁棒性,可重复并且在物种间具有普适性,但我们进一步测试了它们是否能尊重源自受试者间白质解剖固有个体变异的特征。为了证明这一点,我们探索了HCP队列中双胞胎和非双胞胎亚组内纤维束重建的相似性。我们预计同卵双胞胎对将显示出比异卵双胞胎和非双胞胎兄弟姐妹更大的相似性,随后比无关受试者对更大,这与结构连接遗传性的文献一致(Bohlken等,2014;Jansen等,2015;Shen等,2014),并且可能符合双胞胎沟回相似性的文献(Amiez等,2019)。使用HCP队列中可用的72对同卵双胞胎,随机选择的72对异卵双胞胎,72对随机选择的非双胞胎兄弟姐妹和72对随机选择的无关受试者,我们比较了成对受试者间的纤维束,以评估我们的自动化协议是否尊重个体间固有的纤维束变异。对于给定的受试者对和给定的纤维束,我们相关联了两个受试者的归一化路径分布(在MNI空间中并进行阈值处理后)。我们对每个纤维束重复此操作,并计算了跨纤维束的平均相关性。然后对每组受试者对重复此操作,得出每组的平均相关性分布。随后我们比较了不同组之间的这些分布。
3.6. 尊重由于非典型解剖造成的个体差异
我们根据HCP质量控制建议,从HCP队列衍生的图谱中排除了少量因识别出解剖异常的受试者。然而,我们探索了我们的纤维束追踪协议在这些案例中的表现,特别是处理非典型解剖特征和几何畸形的能力。在这些情况下,我们将个体的纤维束追踪结果与相应的纤维束图谱进行了比较,预期前者将尊重解剖异常。
4.结果
4.1. 人类大脑的白质纤维束和白-灰质边界终止图谱
我们将规定的纤维束追踪协议应用于约1000名HCP和1000名英国生物银行受试者。图3展示了从HCP数据集获得的纤维束图谱。为了获得这些图谱,对特定于受试者的MNI变换纤维束进行了二值化,随后平均,以产生图中显示的每个纤维束的人群百分比覆盖率。使用英国生物银行数据获得的图谱显示在补充图1中。
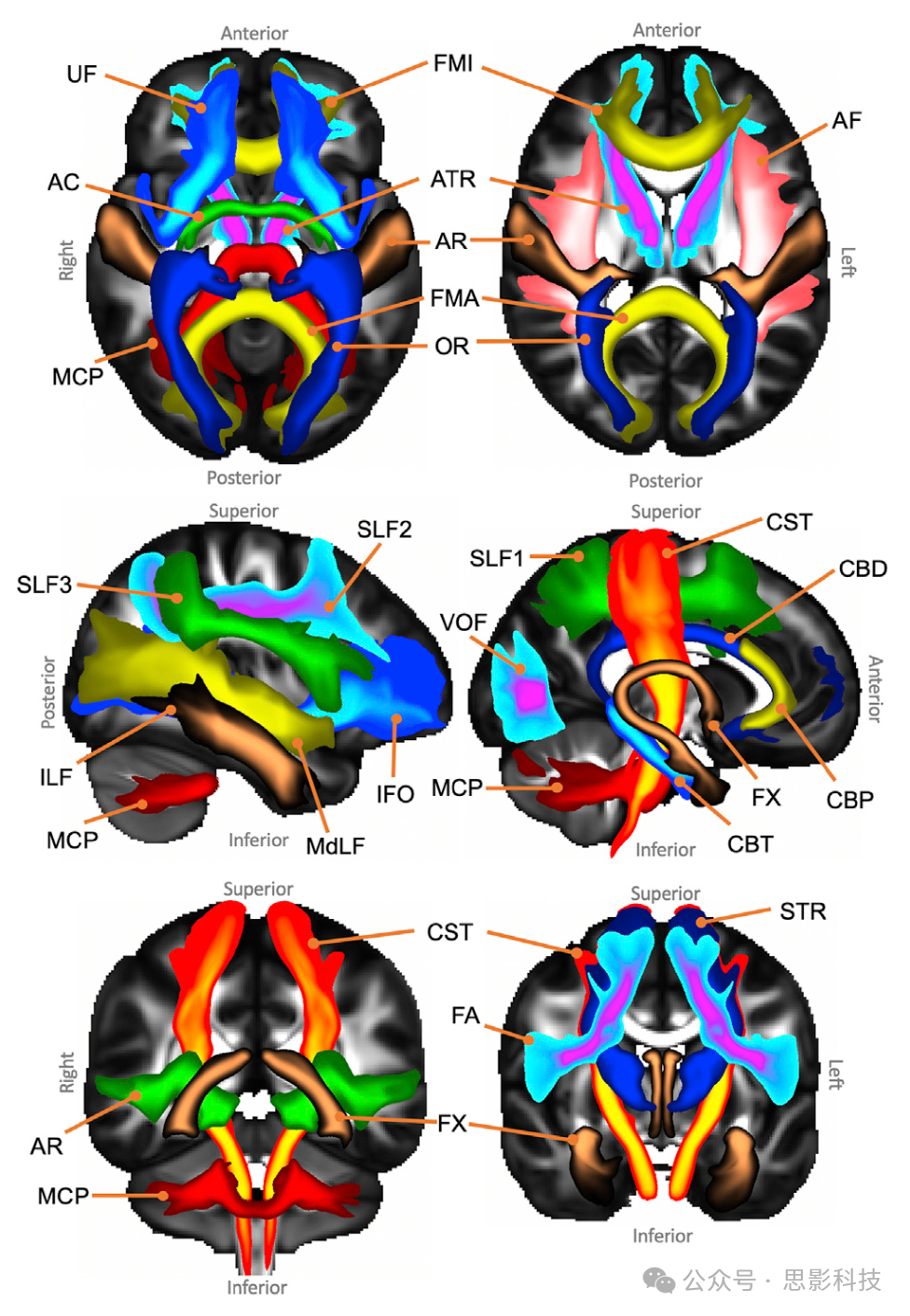
图3. 人群百分比纤维束图谱的轴向、矢状和冠状最大强度投影(为了可视化目的,不同纤维束应用了不同的最大强度投影窗口长度,显示范围=人群覆盖率的5%–100%)。
联合纤维束:弓状束(AF)、额斜束(FA)、下纵束(ILF)、额下枕束(IFO)、中纵束(MdLF)、上纵束I、II和III(SLF)、钩状束(UF)和垂直枕束(VOF)。
投射纤维束:声辐射(AR)、前丘脑辐射(ATR)、皮质脊髓束(CST)、视辐射(OR)和上丘脑辐射(STR)。
边缘系统纤维束:扣带束:围膝部(CBP)、扣带束:颞部(CBT)、扣带束:背部(CBD)和穹窿(FX)。
交叉纤维束:前交叉(AC)、大脑钳(FMA)和小脑钳(FMI)。纤维束图谱是通过对受试者间二值化(阈值为0.5%)的归一化纤维束密度图进行平均而创建的。
连接蓝图也为每个HCP受试者派生,并平均以获得图谱。图4展示了所有HCP受试者平均连接蓝图的列示例,代表每个纤维束在白-灰质边界(WGB)表面的终止点图谱。
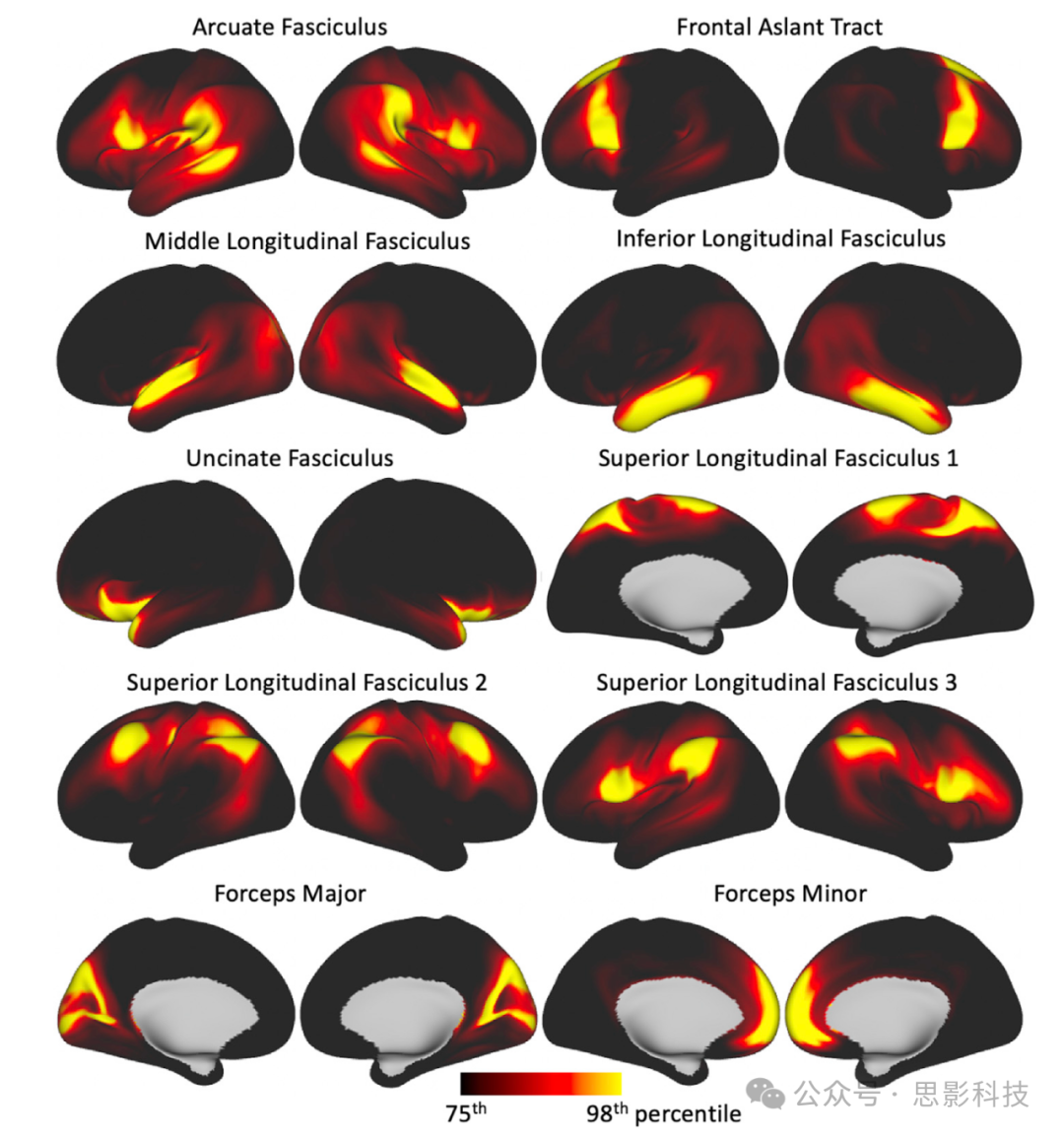
图4. 从HCP队列派生的一部分纤维束的白-灰质边界终点(即平均连接蓝图的列)。
我们还研究了样本量对纤维束图谱创建的影响。对于HCP和英国生物银行的每个队列,我们使用越来越多的受试者数量制作了纤维束图谱,并将每组图谱与1000名受试者的图谱集进行了纤维束间的交叉相关。补充图2显示了每个样本量图谱的纤维束间相关性分布。顶部图表包括一个样本量为10名受试者的图谱集,尽管已显示出高相关性(>0.9),但与使用100名或更多受试者的样本量(>0.98)相比表现相对较差,即从10名受试者的样本量获得的平均相关性明显低于从100名或更多受试者的样本量获得的相关性。
4.2. 对数据集的鲁棒性
为了探索对不同数据质量的鲁棒性,我们比较了纤维束图谱和队列内外纤维束重建的受试者间变异性。为了比较图谱,将HCP图谱集中的每个纤维束与其对应的英国生物银行纤维束图谱进行了交叉相关(每个纤维束图谱应用了30%的人群阈值)。纤维束间的平均相关性为0.80(标准差=0.07)。
受试者间相关性是通过交叉相关随机受试者对的纤维束(即对每个纤维束在MNI空间中的归一化路径分布进行相关)并对每对受试者的纤维束进行平均相关性计算获得的。这一过程在队列内和队列间的许多受试者对中重复进行。为了避免HCP中可能的家族结构引起的偏差,我们将受试者限制在339名无关受试者。我们通过随机选择339名(性别匹配)受试者来匹配英国生物银行数据中的受试者数量。跨队列比较是通过将HCP无关受试者池中的随机受试者与英国生物银行的随机受试者进行相关,得到每个纤维束339个相关性分布。队列内比较得出的平均相关性值分别为HCP的0.51(标准差=0.03)和英国生物银行的0.54(标准差=0.03)(p=8×10^-46)(见图5)。跨队列比较得出的平均相关性为0.41(标准差=0.03),低于队列内比较(p=1×10^-110和2×10^-110),但考虑到数据质量差异和两个队列受试者的年龄差异(HCP:22-35岁,英国生物银行:40-69岁,我们选择的HCP受试者的平均年龄为28.6(标准差=3.7),英国生物银行为62.6(标准差=7.5)),这一结果还是相当可比的。实际上,我们发现HCP队列中的纤维束体积平均比英国生物银行队列大(HCP队列大6.5% ± 11.3%),这与文献中关于年龄相关的白质体积变化一致(Lebel等,2012;Rathee等,2016;Westlye等,2010)。补充图3显示了为HCP和英国生物银行队列重建的每个纤维束的纤维束体积分布。
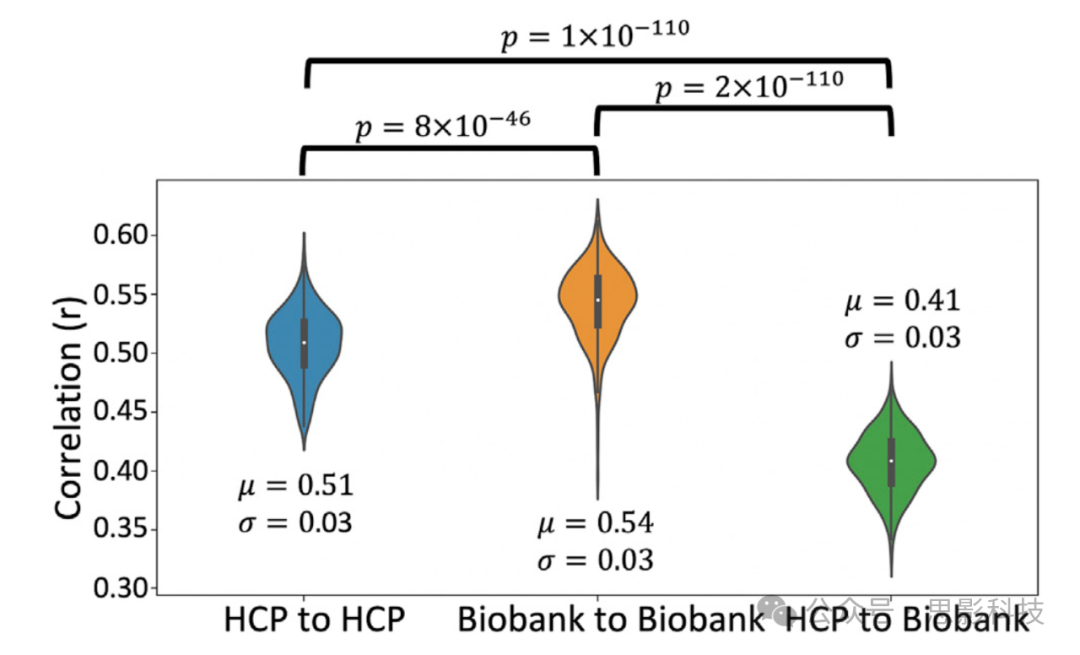
图5. 跨队列鲁棒性总结。
展示了339对受试者在队列内和跨队列间的平均纤维束相关性图。相关性是基于归一化纤维束密度图(阈值为0.5%)进行的。μ是跨纤维束和受试者对的相关性的中位数,σ是标准差。通过Mann-Whitney U检验获得显著性。校正后的p值为0.05/3 = 0.017。
4.3. 物种间的普适性 - 猕猴大脑的白质纤维束和白-灰质边界终止图谱
我们按照上述方法重复进行了纤维束追踪,并从猕猴队列中获得了图谱。我们的纤维束追踪协议定义允许在人类大脑和猕猴大脑中提取同源纤维束。
生成了以人群百分比形式的纤维束图谱,如图6和补充图4所示,而图7展示了使用猕猴数据派生的连接蓝图的平均值。
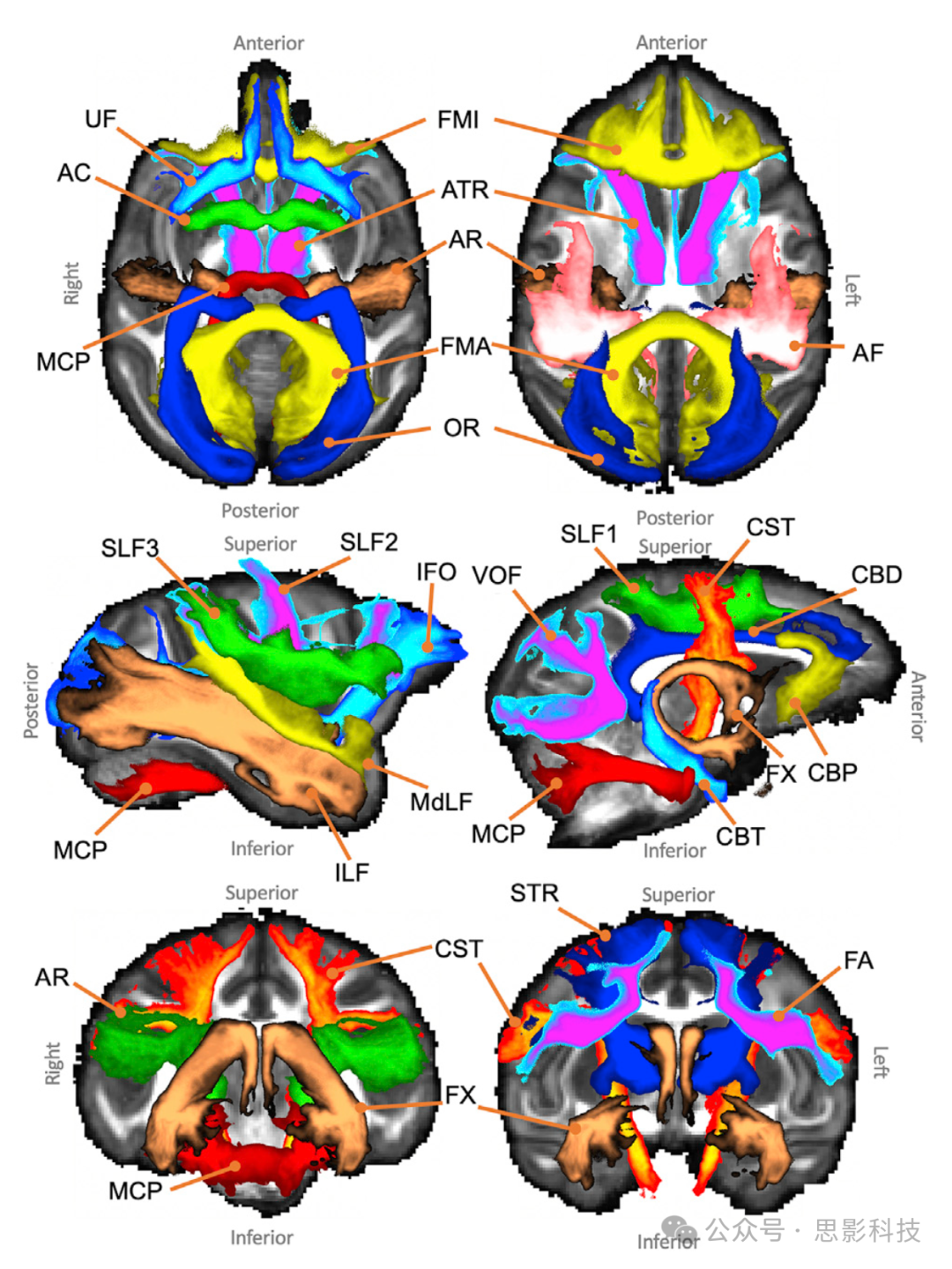
图6. 猕猴受试者的人群百分比纤维束图谱的轴向、矢状和冠状最大强度投影(为了可视化目的,不同纤维束应用了不同的最大强度投影窗口长度,显示范围=人群覆盖率的30%–100%)。
联合纤维束:弓状束(AF)、额斜束(FA)、下纵束(ILF)、额下枕束(IFO)、中纵束(MdLF)、上纵束I、II和III(SLF)、钩状束(UF)和垂直枕束(VOF)。
投射纤维束:声辐射(AR)、前丘脑辐射(ATR)、皮质脊髓束(CST)、视辐射(OR)和上丘脑辐射(STR)。
边缘系统纤维束:扣带束:围膝部(CBP)、扣带束:颞部(CBT)、扣带束:背部(CBD)和穹窿(FX)。
交叉纤维束:前交叉(AC)、大脑钳(FMA)和小脑钳(FMI)。纤维束图谱是通过对受试者间二值化(阈值为0.1%)的归一化纤维束密度图进行平均而创建的。
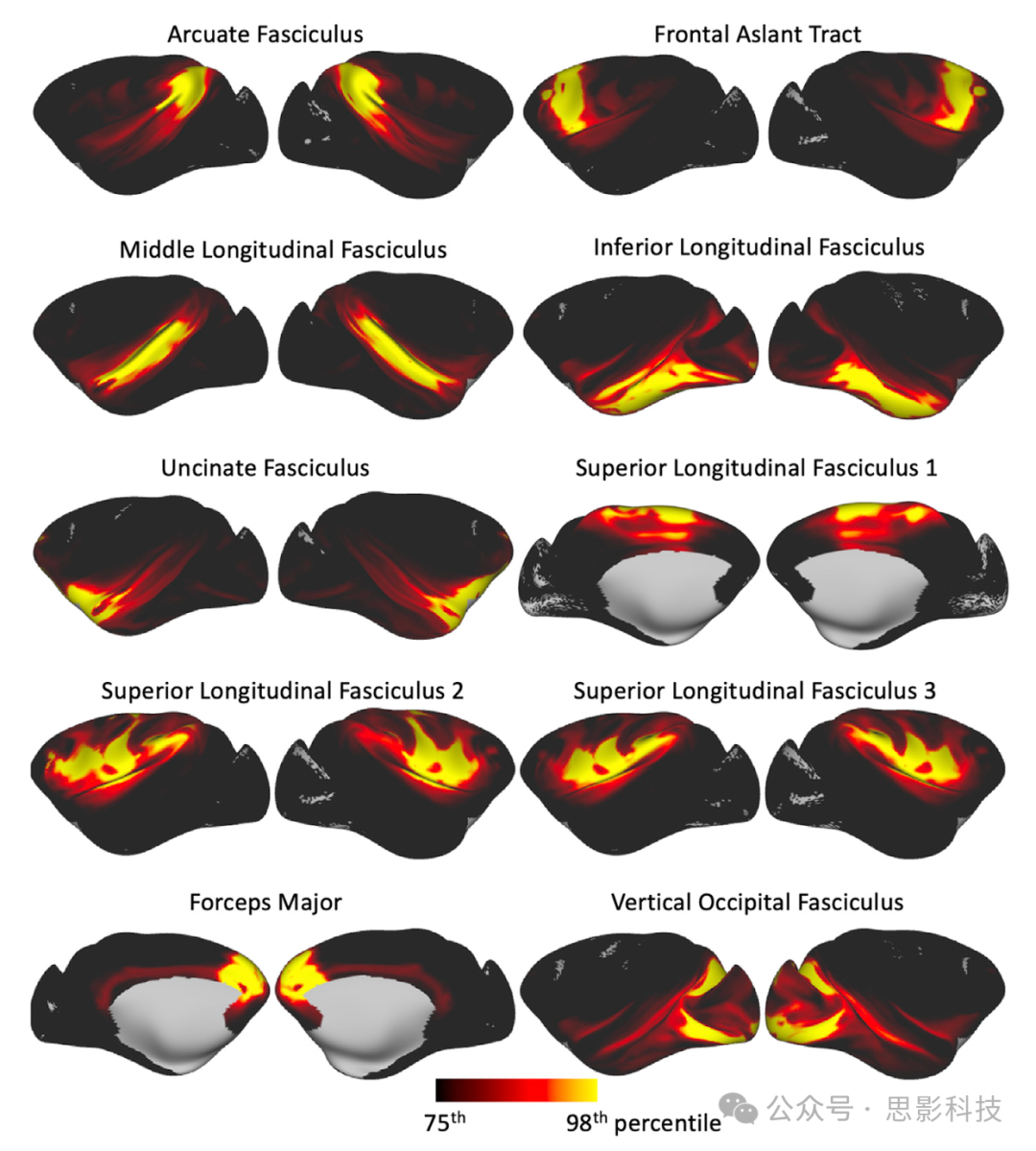
图7. 从猕猴受试者派生的一部分纤维束的白-灰质边界终点(即平均连接蓝图的列)。
4.4. 纤维束侧化
图8展示了人类大脑中一部分纤维束的侧化估计,这些纤维束的侧化在文献中已有报道。如图8所示,AF在HCP和英国生物银行队列中均表现为左侧化,IFO、MdLF和SLF3在两个队列中均表现为右侧化。SLF1在HCP队列中对称,但在英国生物银行队列中达到右侧显著性。SLF2在不同队列中也有变化,在HCP中表现为左侧化,在英国生物银行中表现为右侧化。
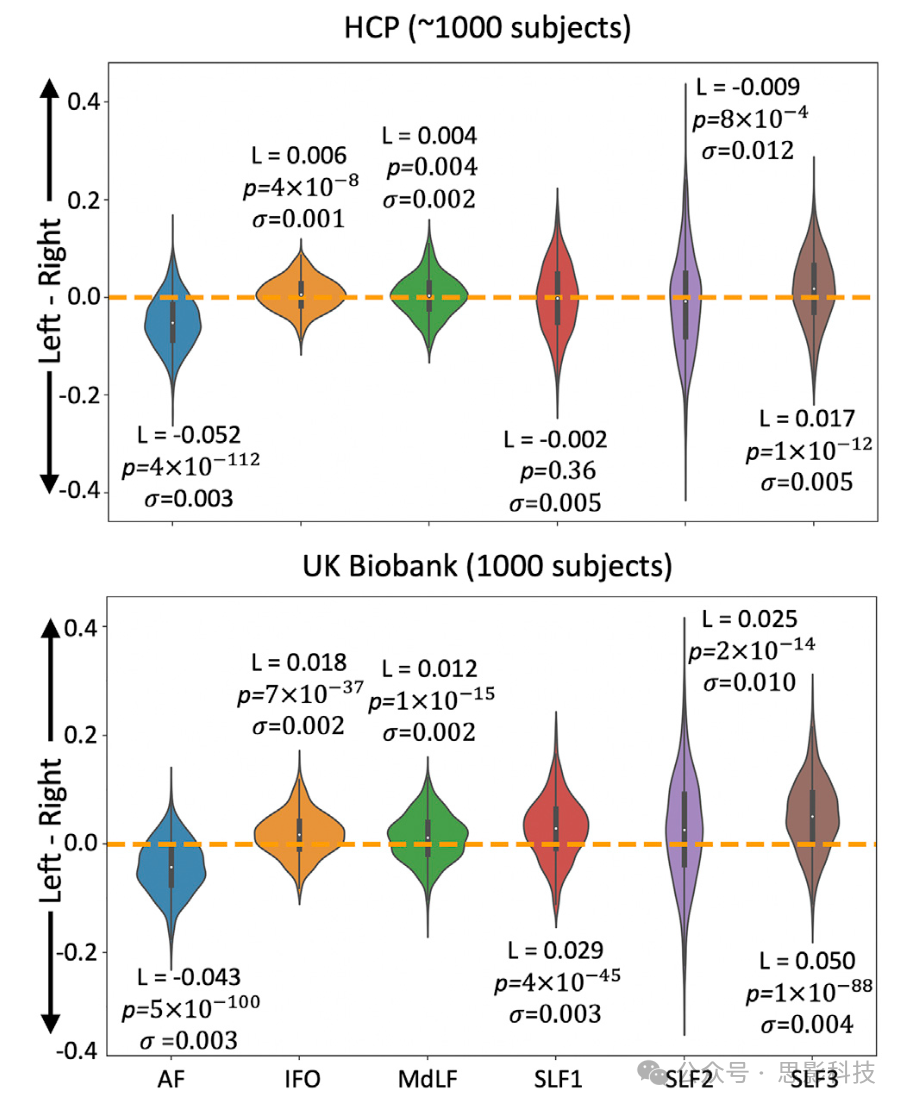
图8. 使用HCP(顶部)和英国生物银行(底部)数据对弓状束(AF)、额下枕束(IFO)、中纵束(MdLF)和上纵束(SLFs)的白质纤维束侧化的总结。
L是队列中白质纤维束侧化的中位数,p是从Mann-Whitney U检验获得的p值,σ是给定白质纤维束侧化的方差。使用了0.5%的阈值来二值化纤维束并获得其体积。校正后的p值为0.05/12 = 0.0042。
除了基于体积的侧化测量外,还使用由纤维束派生的连接蓝图在白-灰质边界表面评估了两半球间的连接模式差异。采用类似于(Mars等,2018b)的方法,计算了Kullback-Leibler(KL)散度,以探索人类大脑两半球间的连接相似性。对于右半球表面的每个位置,最小KL散度值评估了左半球上最相似的连接模式。通过这种方式,我们可以探测在左右半球间显示出不同连接模式的皮层位置,并评估哪些纤维束对这些差异有所贡献。在最小KL散度高的区域,我们预期会观察到相应顶点之间纤维束贡献概况的差异。图9a显示了所有白-灰质边界表面位置获得的最小KL散度值,与Glasser分区的一部分(Glasser等,2016)叠加。左右连接模式差异性高的区域通常局限于额叶和颞顶联合(TPJ)区域。图9b展示了特定白-灰质边界位置在右半球的纤维束对连接模式的贡献,以及这些与左半球最佳匹配位置的连接模式的比较。展示了三个对应于不同程度的两半球间差异性的示例。可以看到,高差异性是由被发现侧化的纤维束介导的。例如,选择在下额沟(IFSa)的一个体素的两半球间差异主要由侧化的SLF3对其连接模式的差异驱动。对于选择在颞顶枕联合(TPOJ1)的中等差异性顶点,我们可以看到小的两半球间分歧主要由AF和SLF3驱动。相反,低差异性区域,如第四视觉区(V4),在贡献于连接概况的纤维束方面显示出很小的两半球间差异。
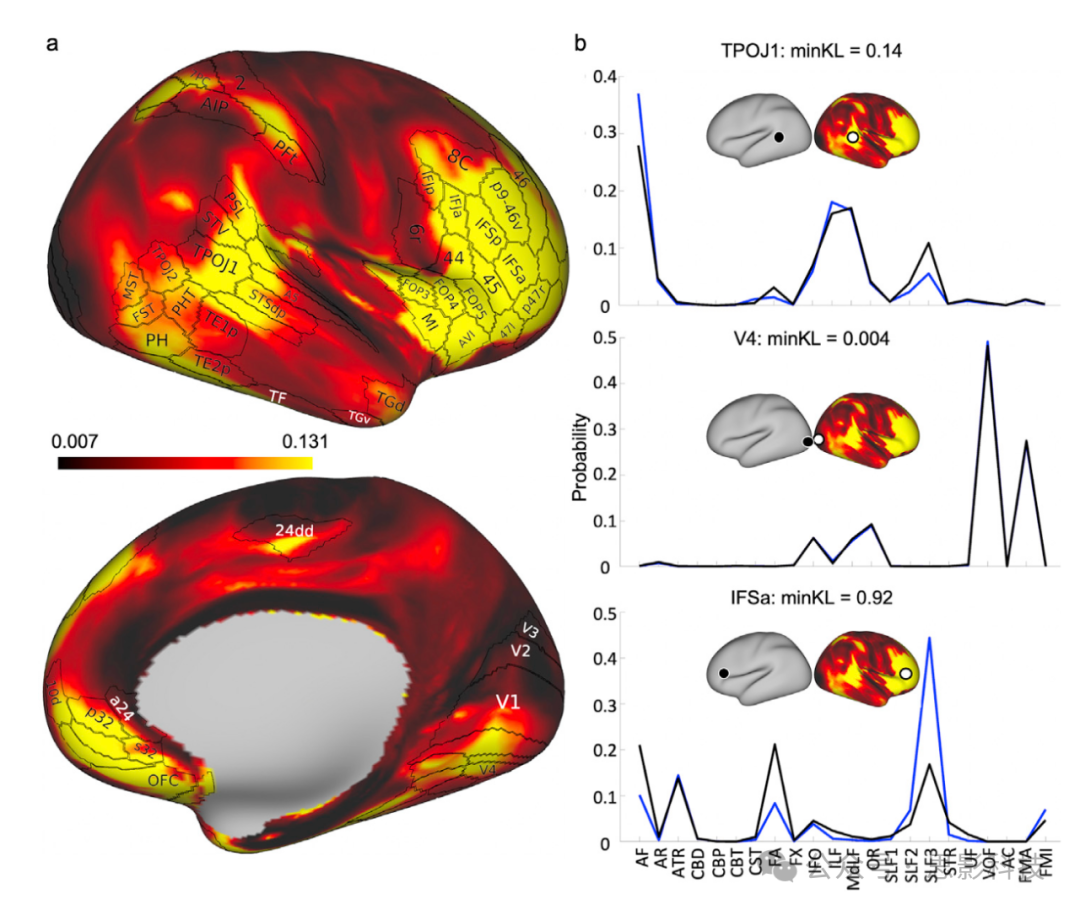
图9. 右半球和左半球之间的最小Kullback-Leibler(KL)散度。
(a) 右半球上的最小KL散度,以及Glasser分区的一部分(Glasser等,Nature,2016),突出显示了差异性区域。
(b) 通过从图9a的右半球选择感兴趣的顶点(白圈)并提取左半球上最相似的顶点(黑圈),可以研究纤维束对位置连接的贡献差异如何导致分歧。黑线对应于右半球顶点的纤维束贡献,蓝线对应于左半球。例如,在V4(中间),最小KL散度很小,这反映在几乎相同的潜在纤维束贡献上。具有中高范围差异性的区域 - TPOJ1(顶部)和IFSa(底部) - 可以看到它们潜在的纤维束贡献有更大的差异,主要由AF和SLFs的差异驱动。
4.5. 双胞胎身份引起的相似性
为了探索纤维束追踪协议是否保留了白质解剖的个体变异性,我们比较了HCP中72对同卵双胞胎、异卵双胞胎、非双胞胎兄弟姐妹和无关受试者的纤维束重建。图10显示了每组的平均受试者间交叉相关性分布(即每个受试者的纤维束平均相关性)。如图10所示,同卵双胞胎对平均(中位数)与其对应的双胞胎相关性更高(0.588,标准差=0.036),相比于异卵双胞胎对(0.545,标准差=0.031)、非双胞胎兄弟姐妹对(0.543,标准差=0.034)和无关受试者对(0.507,标准差=0.029)。Kruskal-Wallis检验显示子组中位数之间存在统计学上显著的差异:χ2=122.3,p=2×10^-26。
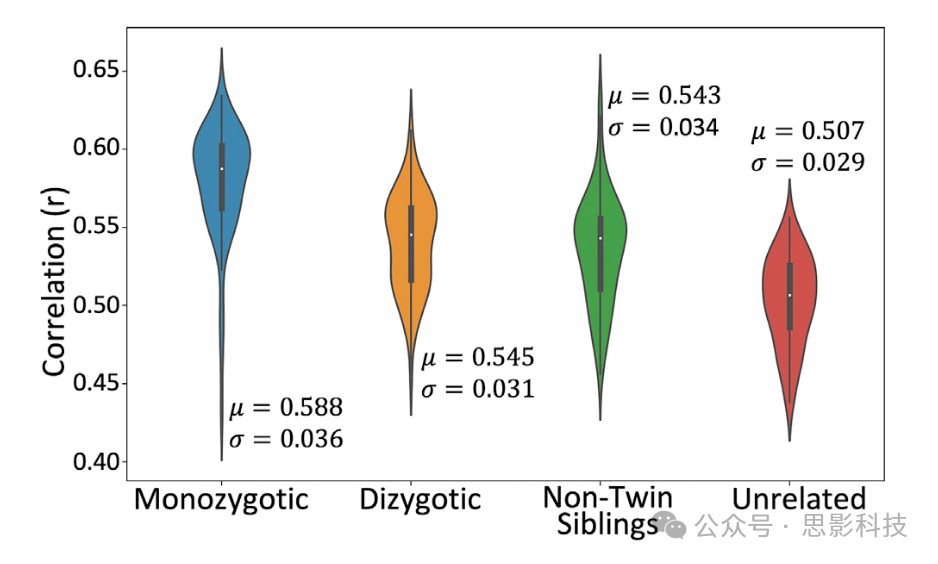
图10. 使用每组72对受试者的双胞胎/非双胞胎白质纤维束相似性。相关性是基于归一化纤维束密度图(阈值为0.5%)进行的。μ是跨纤维束和受试者的组中位数,σ是标准差。使用Kruskal-Wallis检验来确定各组是否来自相同的中位数:χ² = 122.3,p = 2 × 10^-26,平均排名 = 222.6(同卵双胞胎),145.1(异卵双胞胎),134.4(非双胞胎兄弟姐妹)和69.9(无关受试者)。
4.6. 尊重非典型解剖
除了探索双胞胎中纤维束重建的相似性外,我们进一步研究了自动化纤维束追踪是否在解剖异常情况下尊重个体变异。使用HCP质量控制,识别出一些具有明显解剖异常的受试者。图11和补充图5展示了这些受试者的例子,并突出了队列平均纤维束与个体受试者纤维束追踪结果之间的差异,这些差异反映了白质中海绵状血管瘤、囊肿和发育性静脉异常(DVAs)的存在。个体纤维束追踪结果尊重解剖异常,而基于图谱的纤维束掩盖了它们。
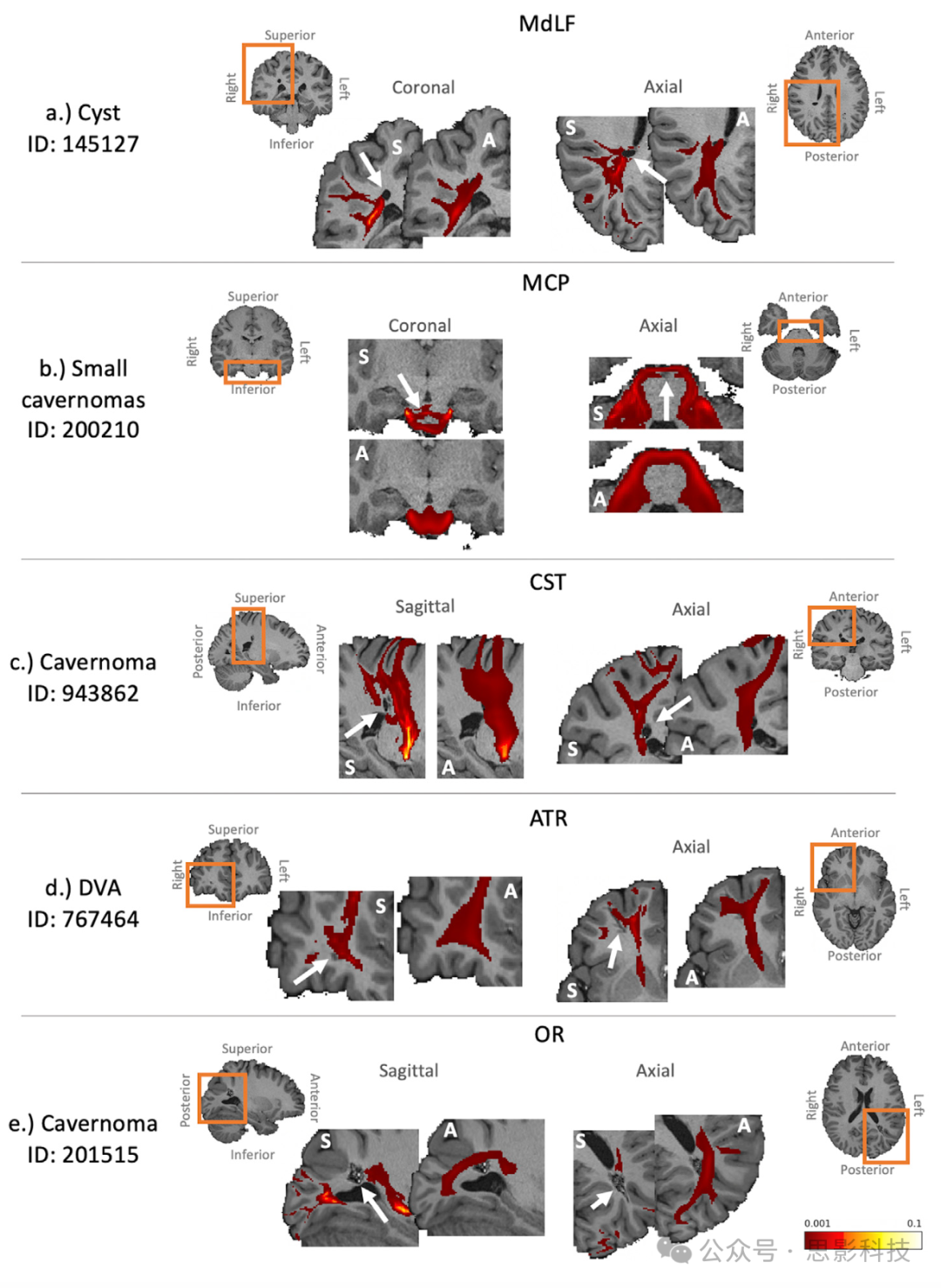
图11. 一些被发现有解剖异常的受试者的纤维束追踪结果示例,显示了队列平均纤维束不尊重潜在的解剖异常。在每个案例中,解剖异常(如HCP质量控制网站所述)由白色箭头指示,受影响的纤维束被呈现。个体受试者提取的纤维束(标记为“S”)和相应的队列平均纤维束(标记为“A”)被叠加在放大的个体T1加权扫描上(橙色框显示放大区域)。纤维束以0.1%的阈值显示。a)右顶叶的囊肿影响MdLF。b)脑干中的小海绵状血管瘤影响MCP。c)右顶叶中的小海绵状血管瘤影响CST。d)右额叶中的发育性静脉异常(DVA)影响ATR。e)左枕叶中的海绵状血管瘤影响OR。
5.讨论
我们介绍了一个新的工具箱(XTRACT),用于自动化概率性纤维束追踪,并提供了用于提取人类和猕猴大脑中白质束的标准化协议。我们已经证明,这些协议在应用于不同图像质量的数据以及非人灵长类动物种类的数据时是鲁棒的。我们使用比以往努力更多的数据量生成了人类白质纤维束图谱,以及使用少量但高质量的体外数据集生成了猕猴图谱。我们进行了间接验证,说明重建的纤维束在左/右不对称方面符合以往文献的预期。我们还展示了,尽管使用标准空间协议自动生成纤维束,这些协议仍然尊重潜在的个体变异性,如双胞胎身份引起的相似性和尊重解剖异常所反映的。该工具箱、纤维束追踪协议和图谱作为FMRIB软件库(FSL)(版本6.0.2及以后)的一部分,可免费公开获取。
在纤维束追踪领域的一个当前问题是,文献中的协议定义通常缺乏细节或未考虑数据共享。在这里,我们提供了一个直接共享标准化协议掩模和纤维束图谱的平台,允许跨研究标准化纤维束追踪协议,有助于提高可重复性。此外,我们使协议定义在物种间具有普适性,以直接促进比较解剖学研究。解剖学家/纤维束追踪者可以交换他们的纤维束定义/方法,并希望在定义和标准化白质纤维束追踪方面达成共识协议,这是当前一个多中心联盟的重点(参见Schilling, Landman, Descoteaux等)。我们相信XTRACT可以为这些努力做出贡献。
Wassermann等人(2016)也提出了一个纤维束追踪协议定义的平台,其中设计了一种白质查询语言(WMQL)。我们的方法使用了与WMQL类似的逻辑,因为它也依赖于掩模和对通过这些掩模的流线进行布尔运算。然而,我们的协议是通用的,其在人类和非人灵长类动物大脑中的实用性已在此处展示。此外,一个主要的概念差异是我们不依赖于自动化的灰质(GM)分区,如WMQL所做的,而是依赖于手动定义的白质掩模。WMQL(白质查询语言)依赖于在白质中播种并通过受试者特定的大脑分区定义皮层端点。使用这种方法追踪到灰质(GM)中存在挑战;在皮层沟回处存在瓶颈,之后纤维束扩散开来,而大多数纤维束追踪算法在解决这种扩散问题上存在困难(Maier-Hein等,2017)。为了缓解这些问题,我们的协议不依赖于GM掩模。我们专注于使用主要位于白质的感兴趣区(ROI)重建感兴趣纤维束的主体。为了获得白质纤维束的皮层终止点,我们采取了相反的方法,从GM表面追踪到白质,从而遵循纤维束预期合并而非扩散的方向。然后我们将表面到白质的纤维束图与纤维束主体的图相乘,以创建纤维束的白质到GM表面投影(连接蓝图)。这避免了一些追踪到表面时的主要问题(Eichert等,2019b;Mars等,2018b)。
我们使用不同质量的成像数据集重建了纤维束,并为人类连接组项目(HCP)和英国生物银行队列生成了图谱。在人类队列内部和跨队列的纤维束重建比较表明,该方法和协议在受试者间和对数据质量具有鲁棒性。HCP和英国生物银行队列分别提供了高质量数据和更典型质量数据的例子。队列内比较显示,在不同质量的数据中观察到类似的受试者间纤维束相关性,英国生物银行观察到更大的受试者间纤维束相关性。这可能反映了与HCP数据相比,分辨率较低的英国生物银行数据的细节水平降低,但也反映了两个队列中受试者的平均年龄差异。此外,我们还使用一小部分猕猴队列生成了图谱。为了补偿受试者数量少,我们使用了高质量和高分辨率的体外数据。相应的结果展示了我们方法对猕猴大脑的普适性。最近获取更大猕猴队列数据的努力正在进行中(采用HCP风格的协议)(Autio等,2019;Milham等,2018),我们的工具将是这些新非人灵长类动物大脑研究计划的有用资源(Thiebaut de Schotten等,2019)。
作为间接验证我们的结果,我们研究了左右纤维束侧化。我们将我们的侧化结果与文献中的先验知识进行了比较。对于两个人类队列(HCP和英国生物银行),我们发现重建的弓状束(AF)强烈左侧化,而上纵束III(SLF3)、下额-枕束(IFO)和中纵束(MdLF)右侧化,这与文献预期一致(Eichert等,2019b;Hau等,2016;Hecht等,2015;Nowell等,2016;Panesar等,2018;Thiebaut de Schotten等,2011a)。对于上纵束I(SLF1)和上纵束II(SLF2),结果不太明确,先前的研究(受试者数量较少)结论不一(Hecht等,2015;Howells等,2018;Thiebaut de Schotten等,2011a)。这可能是由于观察到的大方差(在SLF2的情况下),可能反映了一些潜在的相互作用,如利手性(Howells等,2018)。
我们通过使用从重建的纤维束获得的连接蓝图来研究侧化,进行了进一步的合理性检查。通过使用KL散度来评估两半球间连接模式的差异,我们发现与语言相关的区域(已知是侧化的)在两个半球间具有不同的连接模式(见图9a)(Hiscock和Kinsbourne,2008)。
虽然我们的方法是一种鲁棒的自动化方法,用于一致地重建纤维束,但它也尊重了潜在的解剖变异。我们通过评估同卵双胞胎、异卵双胞胎、非双胞胎兄弟姐妹和无关受试者对之间的纤维束相似性来证明这一点。我们的结果显示,与无关双胞胎相比,双胞胎对之间的相似性更大,这符合遗传性文献的预期(Bohlken等,2014;Shen等,2014)。
我们进一步证明了自动化方法尊重潜在的解剖变异,通过探索解剖异常受试者的纤维束追踪结果与队列平均结果的差异。在所展示的案例中,我们展示了该工具箱能够尊重非典型解剖。这些示例案例当然不是详尽无遗的,但提供了关于工具箱在存在相对较小的解剖异常(由病理引起)时的表现。然而,我们应该指出,我们的方法是基于配准的,因此在存在几何畸形的情况下的追踪性能可能取决于异常的范围和位置以及它对配准的影响(Zhang等,2008)。较小的畸形,如局灶性白质异常/高信号强度,不太可能降低可靠性,特别是考虑到我们的协议定义中使用了包容性ROI,如图11所示,并与最近使用纤维束衍生特征的研究中的类似发现一致(Horbruegger等,2019;Ressel等,2018)。对于较大的畸形(如肿瘤、水肿),即使通过执行条件配准(即在计算扭曲场时屏蔽大型畸形)可以实现一些补偿,追踪精度也可能降低,可能需要考虑特定案例的替代方案(Fekonja等,2019)。尽管如此,许多病理并不引起几何畸形(例如精神/神经发育/心理健康障碍的范围),我们预期我们用于描绘主要白质纤维束的方法在这些情况下是鲁棒的。
6.结论
总之,我们开发并展示了一套鲁棒且标准化的纤维束追踪协议,用于跨物种自动描绘白质束,以及使用这些协议的平台。展示的工具箱(XTRACT)连同纤维束追踪协议和人类/猕猴纤维束图谱,作为FMRIB软件库(FSL版本6.0.2及以后)的一部分,可免费获取。考虑到数据和协议共享的好处,我们预期这个工具箱将有助于提高纤维束追踪领域的可重复性,并促进比较神经解剖学研究。

















 6737
6737

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









