






摘要:将非侵入性脑刺激(NIBS)技术与脑电生理活动记录相结合,在神经科学领域正日益普及。经颅磁刺激(TMS)与脑电图(EEG)的同步组合尤为成功。遗憾的是,为有效与大脑活动相互作用所需的强磁脉冲,不可避免地会在同步 EEG 采集中引入伪影。因此,需要进行细致且有效的预处理,以高效去除伪影。不幸的是,正如文献中已报道的那样,不同的预处理方法可能会导致结果出现差异。在这里,我们旨在对目前可用的三种主要 TMS-EEG 预处理流程,即 ARTIST、TESA 和 SOUND/SSP-SIR 进行特征描述,为需要在不同方法之间做出选择的研究人员提供参考。与以往的研究不同,我们使用具有已知真实情况(即无伪影且待重建的信号)的合成 TMS-EEG 信号来测试这些流程。通过这种方式,能够精确且定量地评估每个流程的可靠性,为未来的研究提供更可靠的参考。总之,我们发现所有流程的表现都不错,但在重建真实信号的时空精度方面存在差异。至关重要的是,这三种流程对试验间变异性的影响各不相同,ARTIST 会引入真实信号中原本不存在的试验间变异性。
1. 引言
先前的文献中对非侵入性脑刺激(NIBS)实验结果的可变性进行了大量讨论。结果的不一致性归因于个体间和个体内的因素。就前者而言,大脑的个体结构和功能特性被认为是导致个体间差异的主要因素之一,这对 TMS-EEG 研究的结果影响尤为显著。在这方面,TMS-EEG 研究中调查的经典指标,如 TMS 诱发电位(TEP)的振幅和潜伏期,已被发现取决于用弥散张量成像(DTI)技术研究的大脑个体结构特性。另一个可变性来源是波动的大脑状态动态,已发现其会调节TMS 对电生理和行为指标的影响。例如,有研究报道,单脉冲 TMS(spTMS)诱发的 TEP 振幅以及重复TMS(rTMS)对 TEP 振幅的调节作用,都取决于刺激时的皮层兴奋性状态。因此,为了减少不同研究结果的混杂,应控制上述个体内和个体间的变异性因素。
为了在 TMS-EEG 记录阶段减少个体间和个体内的变异性,人们提出了不同的技术。例如,Casarotto等人开发了一个工具箱,用于实时监测记录的 TEP 质量和伪影成分的一致性。另一方面,也可以使用其他技术,如 Lioumis 和 Rosanova 建议的,用结构神经导航来辅助 TMS-EEG 记录。
然而,还有另一个显著的变异性来源,它与个体内和个体间的差异无关,而是源于用于从 TMS-EEG 数据中清除伪影的不同预处理流程。事实上,TMS 在 EEG 记录中既会引入皮层反应,也会引入电磁和生理伪影。TMS 脉冲通过利用导电 EEG 电极和导线的电感和电容效应,与之发生电磁相互作用,并在生理上与头皮的神经肌肉系统相互作用。通过磁感应,磁场会感应出电流,产生伪影电位,其振幅可能会使放大器的电子元件饱和。磁梯度与电极和皮肤之间界面的微小电容相互作用,在记录中产生大的波动和衰减伪影。磁梯度还与头皮肌肉相互作用,产生肌肉伪影。所有这些伪影活动掩盖了源于神经过程的真实 EEG 反应。因此,如果不进行精确且有效的预处理,就无法观察到真实的 TMS 诱发电活动(例如 TEP)。多年来,人们开发了不同的预处理流程,以处理 TMS 在 EEG 信号中引入的伪影。最受欢迎的方法是基于独立成分分析(ICA)和像 TESA 中那样手动识别伪影。其他提出的替代方法包括完全自动化的基于 ICA 的预处理流程(ARTIST),以及一种基于非 ICA 的流程,该流程采用 SOUND(利用源估计的噪声丢弃算法)和 SSP-SIR(信号空间投影 - 源信息重建)来校正与 TMS 相关的伪影。
最近的研究试图确定预处理流程的选择对重建的 TMS-EEG 信号的影响,特别是对其变异性的影响。例如,Bertazzoli 及其同事测试了四种不同已发表流程(即 ARTIST、TESA、SOUND/SSP-SIR和 TMSEEG)应用于相同 TMS-EEG 数据集的效果。结果表明,所选择的流程对预处理后的 TEP 有显著影响。具体来说,TEP 的振幅和全局平均场功率(GMFP)在不同流程之间有所不同,头皮上的电位地形图,主要是早期反应,显示出从较差到显著的不同相关性。此外,在两个独立的实验阶段中获得的 TEP 的重测信度在不同流程之间也有很大差异。另一项采用类似方法的研究也支持了这些发现,该研究强调了即使是同一清理流程中的微小变化,也可能导致重建的 TMS 诱发电活动在振幅和空间地形图方面产生不同的结果。综上所述,这些研究表明,用于清理数据的方法对最终的 TMS - EEG 信号有很大影响。然而,它们并没有直接确定每个流程的准确性,即该流程在去除不需要的伪影的同时,能在多大程度上保留由电磁场诱发的皮层反应。这是因为它们只是根据 TEP 和 GMFP 对每个流程的结果进行了交叉比较,而事先并不知道要从 TMS 诱导的伪影中恢复的真实信号是什么。
最近,一些研究通过将模拟的 TMS 伪影叠加到已知的真实 EEG 信号上,来探讨不同方法去除 TMS 相关伪影的效果。例如,Atti 等人测试了独立成分分析(ICA)去除各种模拟 TMS - EEG 伪影的成功率,而 Mutanen 等人则比较了 ICA 和SSP - SIR 在去除 TMS 脉冲诱导的肌肉伪影方面的效果。后一项研究报告称,当伪影的地形图与感兴趣信号的地形图存在显著差异时,SSP - SIR 在清理伪影方面表现更好。此外,这两项研究都强调,如果伪影的变异性较小,基于 ICA 的清理方法容易出错。虽然这些研究至少部分地引入了一个事先已知的真实信号,但它们只关注特定的伪影或分析步骤,而没有考虑整个清理流程的影响,而这正是通常应用于 TMS - EEG 数据的处理方式。
所有这些考虑因素和结果原则上表明,每个 TMS - EEG 实验都应该根据具体的实验假设,从数据采集到预处理进行仔细设计。虽然标准化流程对于确保 TMS - EEG 预处理的一致性和可靠性至关重要,但建立可靠的数据采集程序同样关键,以尽量减少实验者的依赖性,并确保记录的活动准确反映大脑刺激,而不是伪影。然而,考虑到研究背景和临床实践,已发表和标准化的流程通常旨在提供一种合理的标准方法,包括几个预处理步骤,每个步骤都会对其他步骤产生不同的影响。因此,我们在这里测试了三种 TMS - EEG 清理流程(ARTIST、TESA、SOUND/SSP - SIR)的性能,这些流程很好地代表了当前预处理程序的技术水平,我们结合了先前方法的优点进行测试。具体来说,我们通过 (1)将真实的TMS - EEG 伪影叠加到真实的 EEG 信号上,以及(2)对整个流程进行测试来实现这一目的。为了定量评估不同预处理方法对 TMS - EEG 数据的影响程度,我们比较了每个流程在从 TMS 诱导的伪影中清理 EEG 信号的准确性。为此,我们使用一个真实的感觉诱发电位 EEG 信号作为真实情况,并在其上叠加通过 ICA 从真实 TMS - EEG 数据集中提取的典型且真实的 TMS 伪影活动。基于这种方法,被 TMS 伪影掩盖的原始信号是完全已知的。因此,可以使用不同的指标来最佳地评估不同流程在恢复 “真实情况” 方面的可靠性。
本研究的主要目标是表征 ARTIST、TESA 和SOUND/SSP - SIR 流程在从 EEG 信号中清理TMS 伪影的预处理准确性。我们旨在了解哪种流程的整体性能最佳,同时也深入确定使用这些流程时出现的最关键问题。这将为进行 TMS - EEG 实验的研究人员提供有价值的信息,也为他们提供有关所测试的流程与他们的数据匹配程度的额外信息,或者相反,是否需要开发自定义的预处理代码。
我们认为,我们的工作朝着定义清理受污染的 TMS - EEG 数据的 “黄金标准” 迈出了一步,这在基础研究和临床研究中都至关重要。在基础研究中,这是促进不同研究和研究小组之间结果更具可比性的先决条件。在临床研究中,TMS - EEG 越来越多地用于研究精神和神经疾病的神经生理基础,通过确保不同中心的测量结果具有可重复性,这对于生物标志物的开发至关重要。
2. 方法
我们生成了一个测试EEG信号,用于评估三种预处理流程(以下简称"测试信号")。该测试信号是通过将真实TMS-EEG实验中获得的"伪迹信号"(刺激初级运动皮层M1时记录)与来自Zazio等人数据集中一个受试者的"真实信号"(通过周围刺激产生感觉诱发电位SEPs的EEG信号)进行叠加合成的。随后,我们使用三种待评估的预处理流程(ARTIST、TESA和SOUND)分别对测试信号进行独立处理。在完成完整的预处理流程后,将三种处理后的清洁EEG信号分别与原始真实信号进行对比,以评估哪种预处理流程能更好地还原真实EEG数据(图 1)。
2.1 SEPs(真实信号)
选择 SEP 信号作为真实数据的背后逻辑是,SEP 反映了沿躯体感觉通路的神经结构的激活,而该区域与预期的 M1 TMS 伪影出现的区域相近。我们使用了 Zazio 等人描述的 SEP 数据集中一名受试者的 EEG 数据(31 个通道:Fp1、Fp2、C1、C2、CP3、F3、Fz、F4、CP4、FC5、FC1、FC2、FC6、T7、C3、Cz、C4、T8、CP5、CP1、CP2、CP6、P7、P3、Pz、P4、P8、PO7、PO8、O1、O2;国际 EEG 系统的 10 - 05 布局;采样率 5000Hz) 。这项研究是按照《赫尔辛基宣言》的伦理标准进行的,并得到了 IRCCS Instituto Centro San Giovanni di Dio Fatebenefratelli(布雷西亚,19/2016)伦理委员会的批准。通过对左手腕正中神经进行 500 次电刺激来评估 SEP,刺激强度为个体感知阈值的 200%,刺激频率设定为 3.3Hz。SEP 数据在刺激前后- 100 到 200 毫秒的时间范围内进行分段。有关数据收集和实验设置的更多详细信息,请参考 Zazio 等人的研究。
2.2 TMS 相关伪影信号的生成
“伪影信号” 是从对左侧 M1 进行 TMS 刺激的真实 TMS - EEG 数据集中提取的。招募受试者和实验在德国图宾根大学神经病学与中风系以及赫蒂临床脑研究所进行。所有程序均符合图宾根大学医学院当地伦理委员会批准的《赫尔辛基宣言》(810/2021BO2)。所有参与者均提供了书面知情同意书。实验按照安全指南进行。在整个TMS - EEG 实验过程中,参与者坐在舒适的椅子上。使用 130 通道 Ag/AgCl 烧结环形电极帽(德国 EasyCap GmbH 公司)进行 EEG 记录。通过轻度皮肤摩擦准备电极,并填充导电凝胶(美国 GE 医疗系统公司的电极乳膏),直至达到所需的阻抗(<5kΩ)。EEG 和 EMG 使用 24 位生物信号放大器以 5kHz 的采样率同时记录。双相 TMS 脉冲由与 75mm 线圈相连的刺激器发送。运动热点被确定为在对侧第一背侧骨间肌中(通过双极腹 - 肌腱导联的表面电极记录)产生最高且最一致的运动诱发电位振幅的线圈位置和方向,静息运动阈值被定义为在至少 10 次试验中的 5 次中,能引起峰- 峰值运动诱发电位≥50μV 的最低刺激强度。每位参与者接受 800 次 TMS 脉冲刺激,刺激强度为静息运动阈值的 115%,刺激间隔为 2.5s±0.1s抖动。
我们从这项 TMS - EEG 研究中选择了 4 名健康参与者,通过 ICA 方法提取真实的伪影信号。需要注意的是,TMS 开始后的 EEG 信号不仅包含伪影 EEG 信号,还包含刺激诱发的真实脑电位,即 TEP。TMS - EEG 数据预处理流程的目标之一确实是去除脉冲诱导的伪影,同时尽可能完整地保留包括 TEP 在内的活动。因此,在提取要叠加到 SEP 真实信号上的 “真实伪影信号” 时,我们决定使用 4 名健康参与者的数据,而不是仅使用一名受试者的数据。这是因为我们可以合理地假设,不同受试者之间 TEP 的潜伏期和信号形状的个体间变异性,通常大于大多数 TMS 诱导的伪影(例如,脉冲和衰减伪影)的变异性。因此,当使用来自不同受试者的数据时,ICA 算法将更有效地从 TEP 中分离出伪影成分。此外,当预期存在与受试者相关的伪影变异性时,例如肌肉伪影成分,使用多个受试者将导致更具普遍性的伪影成分。
为了提取 TMS 相关的伪影信号,我们首先对 4 名受试者的每个数据集在 TMS 脉冲前后 - 100 到200 毫秒的时间范围内进行分段。通过目视识别并丢弃每个受试者数据中的不良试验,4 名受试者分别得到 748、725、397 和 544 个良好试验(试验总数 = 2414)。同时识别并去除每个受试者的不良通道,并进行插值处理。然后,切除 TMS 脉冲周围的伪影(脉冲周围 - 2 到 5 毫秒);这一步骤被认为是必要的,因为(1)去除伪影周围的时间是所有预处理流程的标准程序;(2)保留脉冲伪影的最相关部分会使 ICA 算法完全受其大振幅的主导。然后将 4 名受试者的 TMS - EEG 记录连接起来,并输入到 FASTICA 算法中。通过 FASTICA 提取的 ICA 成分经过目视检查,其中 6 个被确定为 “仅伪影” 成分,这意味着它们仅携带与 TMS 相关的伪影信号或眼动伪影。通过将这 6 个伪影成分投影回传感器空间,我们获得了一个完整的EEG 伪影信号。之后,仅从伪影信号中保留在真实信号中也存在的 31 个通道,并随机选择 500 个时间段(与 SEP 信号中的时间段数量相同)。在这个过程结束时,伪影信号(图 1)仅包含与 TMS 相关的伪影和眼动成分,可以叠加到真实信号上,以生成下一节中描述的 “测试信号”。
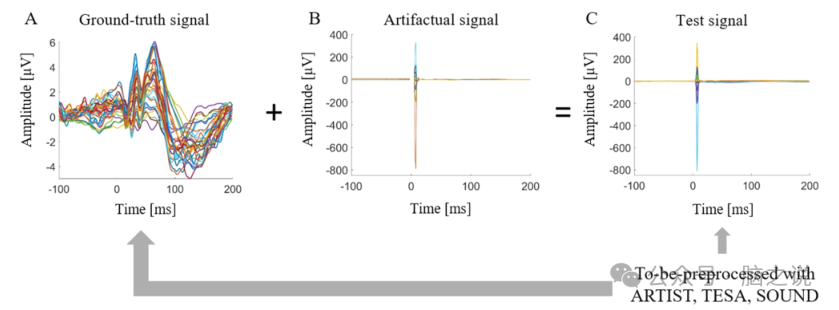
图1 方法工作流程。A)SEP 数据的真实干净信号;B)从TMS - EEG 数据集中提取的 6 个 ICA 成分确定为伪影后生成的伪影信号;C)真实信号和伪影信号叠加生成测试信号。然后,测试信号分别用三种预处理流程(ARTIST、TESA、SOUND)进行预处理,将每个得到的清理后的 EEG 信号与真实信号进行比较,以确定最可靠的流程。
2.3 测试信号的生成
在将真实信号与伪影信号组合以获得最终的 “测试信号” 之前,我们必须实施一个信号对齐程序,以便 (1)使 SEP 数据在振幅上与标准 TEP 具有可比性,(2)使真实信号和伪影信号更加相似。
因此,对真实信号进行了以下调整:首先,去除并插值刺激前后 - 1 到 3 毫秒之间由正中神经刺激引入的噪声。之后,仅为了运行 FASTICA 而应用高通滤波器(0.1Hz),以从真实数据中去除眼动成分。这样做是因为真实数据随后必须与三个预处理信号(ARTIST、TESA 和 SOUND)进行比较,而这些预处理信号会在预处理过程中去除眼动成分。然而,如 2.2 节所述,从 TMS - EEG 数据中提取并与干净信号叠加的一个伪影成分已经包含眼动信息。如果同时保留 TMS - EEG 和 SEP 数据集中的眼动伪影,可能会使预处理流程产生混淆。通过这种方式,不仅可以有力地测试这三种被测试的预处理流程在去除 TMS 相关伪影方面的效果,还可以测试它们去除真实眼动相关信号的能力。此外,真实信号中的 SEP 是通过刺激左手在右侧半球诱发的,因此,为了使 SEP 活动与从涉及左侧 M1 刺激的数据集中提取的 TMS 相关伪影信号位于同一半球,对真实数据进行了半球翻转。此外,对真实信号和从 TMS - EEG 数据中提取的伪影信号的基线活动进行比较后,发现两个信号之间存在偏移。因此,在将真实信号与伪影信号叠加之前,通过分别对真实信号和伪影信号进行逐次试验的基线校正来校正偏移。这个过程使两个信号更加相似。最后,由于 SEP 的偏转幅度通常比 TEP 低,为了使我们的真实信号与经典的 TMS 诱发电活动具有可比性,将 SEP 的振幅乘以 2 进行缩放。
此时,真实信号可以与伪影信号叠加,生成最终的测试信号,以便使用下一节中描述的三种流程进行预处理。
2.4 应用于测试信号的预处理流程
2.4.1 ARTIST
全自动的 ARTIST 流程原则上具有通过自动化所有预处理步骤来减少因用户选择而导致的个体差异的优势。对于测试信号的预处理,ARTIST 丢弃了两个不良通道(T7 和 T8)。此外,它将 500 次试验中的72 次标记为不良试验。关于 ARTIST 用于消除与 TMS相关的衰减伪影成分的第一轮 ICA,ARTIST 发现有一个成分携带这种伪影,因此在第一轮 ICA 中消除了一个伪影成分。在旨在消除除衰减伪影之外的所有其他伪影成分的第二轮ICA 中,ARTIST 从信号中提取的 25 个成分中丢弃了 7 个伪影成分。
2.4.2 TESA
TESA 半自动流程标记了 3 个要剔除的通道。此外,TESA 将 16 次试验标记为伪影时间段。对于仅旨在丢弃携带 TMS 相关肌肉伪影成分的第一轮 ICA,TESA 从 28 个成分中丢弃了 1 个。在第二轮 ICA 中,从 27 个成分中发现 1个与眼动伪影相关的不良成分和 7 个与电极噪声相关的不良成分,因此在第二轮 ICA 中总共从信号中丢弃了 8 个不良成分。
2.4.3 SOUND/SSP - SIR
SOUND/SSP - SIR 不是一个自动化流程,因为它要求用户标记不良通道(如果用户选择不完全将此步骤委托给 SOUND)、不良试验、从该流程中仅有的一轮 ICA 中提取的眼动伪影成分,以及用 SSP - SIR 提取的携带 TMS 相关肌肉伪影的独立成分。在本次应用 SOUND 清理流程时,用户未标记要剔除的通道。此外,39 次试验被标记为不良试验。关于识别携带眼动伪影成分的 ICA 轮次,30 个成分中有 2 个被标记为眼动伪影。最后,SSP - SIR 识别出的前 2 个独立成分因携带肌肉相关伪影而被标记为要剔除。
2.5 预处理质量评估
为了评估每个预处理流程的重建质量,我们研究了 1)真实信号和重建信号在统计上是否存在差异;2)预处理对试验间变异性有何影响;3)真实信号和重建信号在多大程度上线性相关。
我们主要将分析重点放在一组感兴趣的通道(P3、CP3、CP5、CP1、C3)上,预计在这些通道上SEP 和 TMS 伪影的影响最为显著,同时也关注三个感兴趣的时间窗口(TOI),这些时间窗口是根据在真实数据中观察到的关键偏转来定义的。然而,我们也报告并讨论了整个感兴趣时间窗口(5 - 200 毫秒)和所有通道的结果。所有后续分析均使用基于 NumPy 和 SciPy 的自定义 Python 代码,以及部分基于 FieldTrip 的自定义 MATLAB 代码进行。
在进行任何后续分析之前,将真实 EEG 数据和预处理后的 EEG 数据均重新参考到 Cz 电极,并进行基线校正(基线期为 - 100 到 - 15 毫秒)。这三种流程都使用共同平均法进行重新参考,尽管在不同步骤进行(见图 2)。因此,我们进一步选择中央电极 Cz 进行最终的统一重新参考,因为共同平均法可能会潜在地掩盖全局效应。

图2 每个预处理流程中的步骤顺序。报告了各流程间的通用参数。
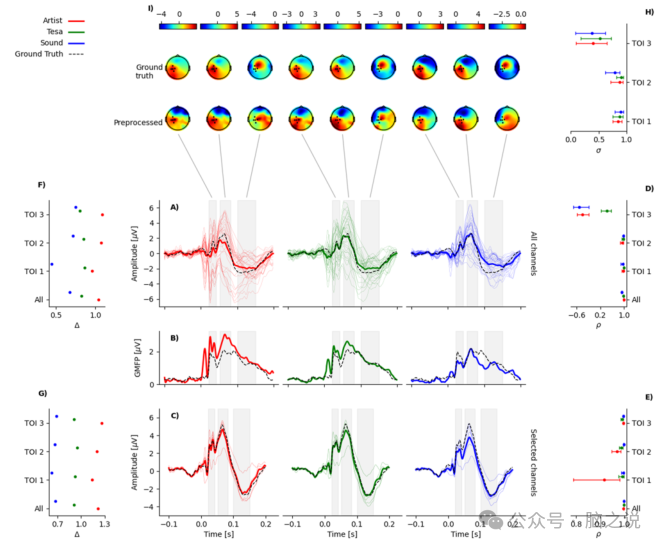
图3 流程质量评估结果总结。该图按行和列进行概念性组织。第一行涉及空间信息,而最后三行分别涉及所有通道、全局平均场功率和选定通道的时间信息。列表示试验间变异性比率(第一列)、平均 EEG 活动(中间列)、皮尔逊相关性(最后一列)。此外,每个流程的绘图内容用颜色编码(红色:ARTIST,绿色:TESA,蓝色:SOUND)。
对于每个流程及其对应的真实信号,我们分别计算 1)每个通道的试验平均值,2)将感兴趣的通道(P3、CP3、CP5、CP1、C3)汇总后的平均值,3)所有通道汇总后的平均值。在这些信号的每一个上,我们进行非参数统计比较,原假设为重建信号和原始信号之间没有差异。我们使用基于聚类的置换检验,仅利用时间维度作为聚类形成特征进行多重比较校正,因为通道数量较少且它们之间的距离不均匀,不允许进行可靠的空间聚类。测试参数如下:5000 次置换;聚类形成阈值 = 0.05;聚类显著性阈值 = 0.05;双尾检验并进行 Bonferroni 校正;感兴趣时间 = 5 - 200 毫秒。除了信号平均值之外,还像 Esser 等人那样,计算所有预处理信号和真实信号的全局平均场功率。
3. 结果
在图 3 中,展示了三种流程预处理后的 EEG 信号和所有通道平均的真实信号(图3 A)以及选定通道(P3、CP3、CP5、CP1、C3)平均的信号(图3 C)。此外,图3 B展示了三种预处理信号的 GMFP 与相应真实信号的 GMFP 的对比。
尽管从视觉上可以观察到重建信号跟踪真实信号的方式存在差异,但统计置换检验表明,对于所有流程以及两种平均模式,均未发现显著不同的时间聚类。图 3 D 和 E中所示的整个时间窗口的高时间相关性证实了这一结果。从进一步报告的所有ρ值可以看出,所有流程和 TOI 中,原始信号和重建信号之间的线性相关性都很高,除了在所有通道平均的情况下,第三个 TOI 的时间相关性小于 0.5,在某些情况下甚至为负,其他所有时间相关性均大于 0.9。
图 3 F 和 G展示了感兴趣时间内三种流程的试验间变异性比率 Δ 的值。从这些数据可以看出,SOUND预处理总是降低所考虑样本的试验间变异性,而 ARTIST 则增加了它。相反,TESA 流程的应用对试验间变异性影响不大。
最后,图 3I展示了每个 TOI 以及重建信号和原始信号的活动地形图,以及相应的空间相关值(图 3 H)。在所有 TOI 中,除了第三个 TOI,所有流程在皮尔逊相关性方面都能较好地保持空间结构,在第三个 TOI 中,三种流程的 σ 值都接近或低于 0.5。
图 4 展示了每个流程和 TOI 的试验间变异性比率的地形图。可以清楚地看到,ARTIST 系统地增加了所有通道的Δ值,SOUND 降低了它,而 TESA 则使其保持相当稳定,除了 C1 通道,其 Δ=2.15。图4 B中的插图报告了跨通道和感兴趣 TOI 平均的 Δ 值,证实了相同的趋势。
图 5 进一步展示了每个流程和 TOI 的时间相关性地形图。为了清晰起见,色图在 ρ≤0 处进行了低切。可以注意到,在感兴趣的通道中,所有流程在所有 TOI 中与原始信号的线性相关性都很好。在图5 B中也可以看出这一点,其中报告了感兴趣通道上平均的皮尔逊时间相关性。然而,地形图也显示了对于远离刺激时空点的通道和 TOI,原始信号和重建信号之间的线性相关性如何下降。
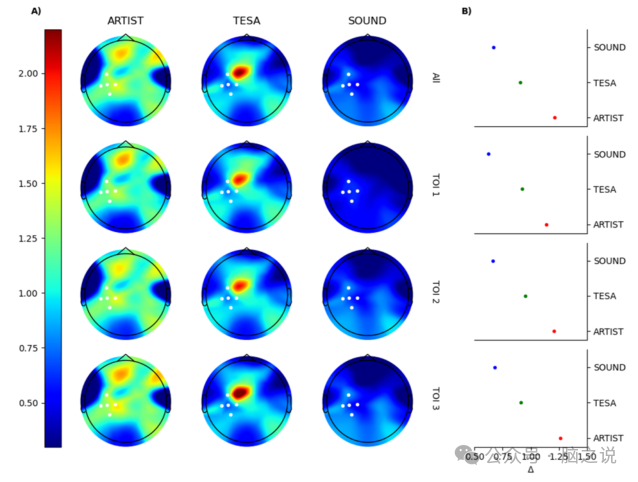
图4 A)每个流程和感兴趣时间窗口(TOI)的试验间变异性比率 Δ 的地形图。所有地形图均设置为相同的颜色刻度以便比较。B)每个 TOI 和整个时间内,感兴趣通道(P3、CP3、CP5、CP1、C3)的试验间变异性比率平均值。每个流程的结果用颜色编码(红色:ARTIST,绿色:TESA,蓝色:SOUND)。
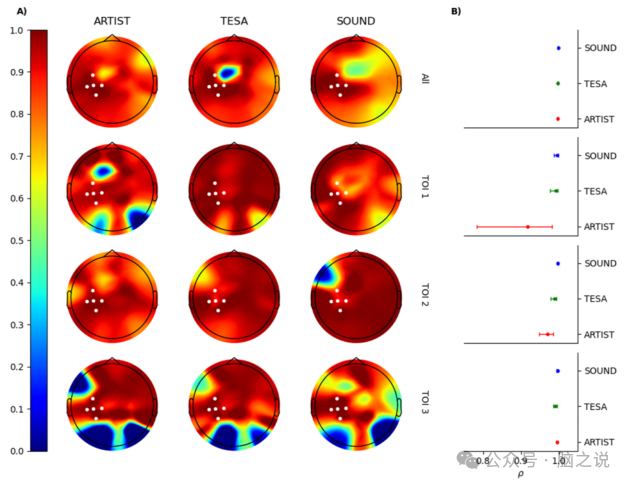
图5 A)每个流程和感兴趣时间窗口(TOI)的时间相关性ρ的地形图。所有地形图均设置为相同的颜色刻度以便比较,为清晰起见,所有 ρ≤0 的值均用深蓝色编码。B)每个 TOI和整个时间内,感兴趣通道(P3、CP3、CP5、CP1、C3)的时间皮尔逊相关性平均值。条形表示 95% 置信区间。
4. 讨论
在这项工作中,我们比较了三种具有代表性的 TMS - EEG 数据集预处理流程的准确性,即 ARTIST、TESA和 SOUND/SSP - SIR。与以往的工作不同的是,我们采用了真实信号恢复方法,即从真实的 EEG 数据和真实的 TMS - EEG 伪影合成一个待清理的数据集。由于确切知道要恢复的信号,我们能够将合成数据输入到各个流程中,并从定性和定量两个方面评估从 TMS 诱导的伪影中提取真实 EEG 信号的可靠程度。此外,我们像在真实实验分析中一样将流程作为一个整体来考虑,从而一次性解决预处理步骤的顺序和组合所产生的影响。
一般来说,所有流程在恢复真实信号方面都表现出良好的性能,这一点通过对所有通道和选定通道平均值进行的置换聚类统计显示,每个预处理信号与真实信号之间均无显著差异得以证明。在所有通道和感兴趣通道上观察到的整个感兴趣时间窗口(TMS 后 5 - 200 毫秒)内的高时间相关值(图 3D - E、图 5)也证实了这一结果。所有预处理流程与它们的真实信号都保持着很强的线性相关性,这支持了它们的整体有效性。
我们还检查了三个不同的感兴趣时间窗口(TOI),这些时间窗口是根据真实数据中的主要偏转选择的,以及一组预计携带大部分伪影和感兴趣信号的通道(‘P3’、‘CP3’、‘CP5’、‘CP1’和‘C3’)。这些分析能够更详细地描述流程的性能,并揭示了一些关键差异。
从图 3D - E和图 5 中不同 TOI 的时间相关性可以看出,预处理的稳健性在时间上最接近刺激的时间点(TOI 1)最高。这对于 TESA 和SOUND/SSP - SIR 尤其如此,而 ARTIST 预处理后的信号在 TOI 1 中对于感兴趣通道的置信区间已经略大,这表明其线性相关性不太精确。尽管如此,所有三个预处理信号在所有 TOI 中,特别是在具有最强感兴趣效应的通道周围,都显示出较高的相关值(见图 5)。最后,在空间相关性(图 3H)中也可以观察到相同的趋势,即所有流程在第三个 TOI 中的 σ 值都急剧下降。这种时间和空间上的一致性表明,预处理通常在保留靠近刺激热点 / 时间的神经效应方面特别有效,这些效应可能携带最强的 TMS 诱发神经信号。相反,研究人员在解释远离刺激部位 / 时间的预处理数据时必须特别小心,因为结果可能强烈依赖于所选择的流程。
关于早期成分中保留的这种时间一致性,必须考虑到在我们的特征分析中,我们在刺激前后 - 1 到 5 毫秒之间对数据进行了切割和插值。在大多数研究中,TMS 伪影的切割和插值时间更长,即不同研究中为 5 到 15 毫秒,尽管在适当的放大器设置下,TMS 伪影可能小于 5 - 6 毫秒。这种差异有利于我们的评估过程,因为它可以更有效地评估流程在早期成分(5 - 70 毫秒)方面的性能。评估流程在这些早期偏转上的可靠性非常重要,因为它们被认为是大脑的真实反应,并且被认为反映了与运动皮层相连区域的快速激活。特别是,可以注意到在 TOI 1 中研究的偏转方面,SOUND/SSP - SIR 和 TESA 在信号重建的准确性上略优于 ARTIST,尤其是当只考虑感兴趣通道时。这种诱发电位与 TMS 相关的 N45 偏转最为相似。在过去几年中,这种 TEP 已被证明是 TMS - EEG 实验中兴奋 / 抑制平衡的一个指标。事实上,像地西泮和阿普唑仑这样的 GABAAR 正变构调节剂会增加 N45。同样,NMDAR 拮抗剂右美沙芬也会增加 N45。此外,Darmani 等人发现选择性α5 - GABAAR 拮抗剂 S44819 会降低 N45 的振幅。综上所述,目前的证据表明,N45 可能是 TMS - EEG 实验中兴奋 - 抑制平衡的一个潜在生物标志物。因此,以精确和稳健的方式可靠地获取其振幅变化至关重要,从而在这个感兴趣的时间窗口中表征不同 TMS - EEG 预处理流程的可靠性。
预处理质量评估中的另一个关键点是流程对试验间变异性的影响。进行这项分析的假设是,预处理流程绝不应增加原始信号中已有的试验间变异性。然而,从图 4可以看出,情况并非总是如此。我们发现 ARTIST 总是增加真实信号的原始试验间变异性(Δ≥1);至关重要的是,当考虑最接近刺激部位的通道时,这种增加更为明显。不同的是,TESA 相对于其真实信号使试验间变异性保持相当稳定,而 SOUND/SSP - SIR甚至降低了它(Δ<1)。简而言之,虽然 TESA 和 SOUND/SSP - SIR 不会引入原始真实数据中不存在的变异性,但 ARTIST流程会。ARTIST 在预处理信号中引入的这种额外的虚假变异性,对于那些对比较用 ARTIST 预处理的 TMS - EEG 数据集的实验条件感兴趣的研究人员来说,是一个关键信息。事实上,试验间变异性的增加可能会对一种非常常见的情况产生强烈影响,即在这种情况下,通过基于置换的统计方法比较不同实验条件下的 EEG 数据。在这种情况下,更大的变异性会影响通过置换计算的统计经验分布,可能会掩盖真正的神经效应。
在解释这些结果时,必须指出我们研究的一些局限性,特别是考虑到合成测试信号的生成方式可能会干扰流程的性能评估,因为不同流程对于伪影和噪声的识别与去除有其特定的方法。事实上,我们通过 ICA 分解来分离 TMS - EEG 伪影,选择这种方法是在提取真实伪影成分的时间和准确性之间的最佳折衷。必须指出的是,原则上可以通过利用相关过程的物理学原理,更精确且真实地生成合成信号。可以使用从高分辨率磁共振解剖扫描计算出的精确正向模型,并在传感器层面模拟生理上真实的 EEG 活动,来生成干净的 EEG 真实信号。另一方面,原则上,TMS 诱导的伪影可以通过考虑电极的电容和电感效应,并使用先前获得的正向模型在神经肌肉区域重新创建 “组织效应” 来生成。然而,这需要在利用正确的生物物理建模方面付出巨大努力,这超出了当前工作的范围,但可能是进一步研究的主题。
在这种情况下,必须考虑到 ARTIST 和 TESA 都使用相同的 ICA 分解方法来去除 TMS 和其他噪声伪影。相反,SOUND/SSP - SIR 是基于递归逆建模(SOUND)和信号源投影(SSP)的混合方法。ICA 算法依赖于在压缩时间维度后创建通道数据的多元直方图(或分布),并对多元通道数据集应用变换,以找到数据的 “统计独立” 表示。然后通过找到一种变换,使变换后通道的多元直方图(即成分)尽可能不像正态分布的乘积,从而实现统计独立性。从这个意义上说,单个成分直方图已经被变换为近似最大非正态分布,原则上这可能有利于像 ARTIST 和 TESA 这样基于ICA 的方法。尽管这种局限性给 SOUND/SSP - SIR 带来了潜在的负面偏差,但该流程在定性和定量方面都表现得非常好,特别是在靠近刺激部位和通常受 TMS 诱导伪影影响最大的早期成分方面。
流程处理噪声去除的不同方式也可以解释在试验间变异性比率中发现的差异。事实上,ICA 也被用于清理真实数据,而 SOUND/SSP - SIR 利用了基于信号源投影和递归逆建模方法的优势。因此,SOUND/SSP - SIR 预处理中试验间变异性的降低可以归因于其处理噪声成分的不同方式,可能清理了真实数据中仍然存在但未被标准 ICA 进一步分离的噪声。虽然 TESA 得到的结果原则上是预期的,但 ARTIST 即使基于相同的 ICA 分解方法,却广泛引入了原始信号中原本不存在的变异性,这是一个必须考虑的不利影响,特别是当在记录阶段已经使用不同技术努力最小化变异性时,如引言中所述。
我们方法的另一个局限性是,由于在原始 TMS - EEG 数据集中听觉成分已经被有效地掩盖,我们的流程没有处理由 TMS 点击声引起的听觉伪影。然而,鉴于最近开发出了在源头掩盖这种伪影的有效方法,这应该不再被视为一个主要问题。事实上,这里评估的流程都没有提供针对听觉伪影的任何特定校正。此外,我们的特征描述设计在评估初级运动皮层的 TMS - EEG 流程性能方面是最优的,因为我们的真实信号位于该区域。
最后,我们必须认识到一个局限性,即我们通过将 ICA 提取的噪声成分线性叠加到干净的真实信号上来生成测试信号的方式,可能导致信号的噪声结构比真实的 TMS - EEG 数据集更简单。如前所述,合成一个更现实的 TMS - EEG 测试数据集需要大量的建模工作,这超出了当前工作的范围。然而,我们认为我们的结果是有效且可推广的,因为原则上这种局限性应该会促进像 ARTIST 这样基于 ICA 的流程的性能。但实际情况并非如此,基于 ICA 的流程表现更差。相反,ARTIST 的表现尤其在试验间变异性方面存在问题。
总之,就本研究中所描述的而言,所有预处理流程在刺激部位的时空邻近区域重建原始信号方面都可以被认为是稳健的,尽管 TESA 和 SOUND/SSP - SIR 在早期成分中的变异性略小于 ARTIST。另一方面,对于所有流程,远离刺激时间和部位的结果都必须谨慎解释。至关重要的是,从试验间变异性的分析中得出的最显著结果表明,在预处理数据将用于置换检验的情况下,TESA 和 SOUND/SSP - SIR 比 ARTIST 更值得推荐,因为 ARTIST 引入的更大试验间变异性可能最终会掩盖实验条件之间的差异。我们认为本文提供的信息对基础研究和临床环境中的研究人员都有价值。此外,我们认为我们的结果在定义 TMS - EEG 实验 “黄金标准” 这一关键任务中又迈出了一步。
参考文献:Towards the definition of a standard in TMS-EEG data preprocessing.

















 892
892

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









