






密码子
| 氨基酸 | 三字母缩写 | 一字母缩写 | 密码子 |
|---|---|---|---|
| 丙氨酸 | Ala | A | GCU, GCC, GCA, GCG |
| 精氨酸 | Arg | R | CGU, CGC, CGA, CGG, AGA, AGG |
| 天冬酰胺 | Asn | N | AAU, AAC |
| 天冬氨酸 | Asp | D | GAU, GAC |
| 谷氨酸 | Glu | E | GAA, GAG |
| 谷氨酰胺 | Gln | Q | CAA, CAG |
| 甘氨酸 | Gly | G | GGU, GGC, GGA, GGG |
| 丝氨酸 | Ser | S | UCU, UCC, UCA, UCG, AGU, AGC |
| 组氨酸 | His | H | CAU, CAC |
| 异亮氨酸 | Ile | I | AUU, AUC, AUA |
| 亮氨酸 | Leu | L | UUA, UUG, CUU, CUC, CUA, CUG |
| 赖氨酸 | Lys | K | AAA, AAG |
| 甲硫氨酸 | Met | M | AUG |
| 苯丙氨酸 | Phe | F | UUU, UUC |
| 脯氨酸 | Pro | P | CCU, CCC, CCA, CCG |
| 硒代半胱氨酸 | Sec | U | UGA |
| 硫氨酸 | Thr | T | ACU, ACC, ACA, ACG |
| 缬氨酸 | Val | V | GUU, GUC, GUA, GUG |
| 色氨酸 | Trp | W | UGG |
| 酪氨酸 | Tyr | Y | UAU, UAC |
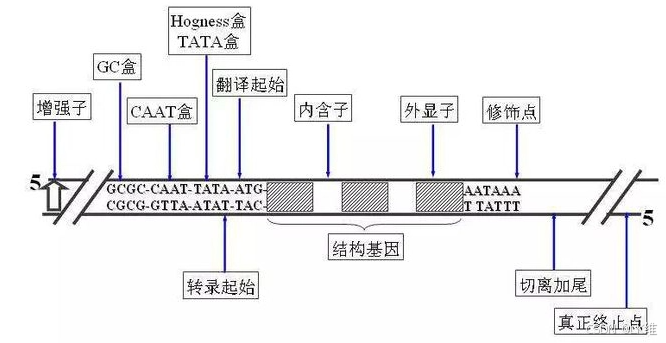
简并碱基符号(IUPAC)
| 符号 | 含义 | 包含的碱基 |
|---|---|---|
| A | 腺嘌呤 | A |
| C | 胞嘧啶 | C |
| G | 鸟嘌呤 | G |
| T/U | 胸腺嘧啶(DNA中)/尿嘧啶(RNA中) | T/U |
| R | 嘌呤 | A 或 G |
| Y | 嘧啶 | C 或 T/U |
| S | 强配对 | G 或 C |
| W | 弱配对 | A 或 T/U |
| K | 背对配对 | G 或 T/U |
| M | 面对配对 | A 或 C |
| B | 不是A | C 或 G 或 T/U |
| D | 不是C | A 或 G 或 T/U |
| H | 不是G | A 或 C 或 T/U |
| V | 不是T/U | A 或 C 或 G |
| N | 任意碱基 | A 或 C 或 G 或 T/U |
终止密码子
| 密码子 | 功能 |
|---|---|
| UAA | 终止 |
| UAG | 终止 |
| UGA | 终止 |
起始密码子
| 密码子 | 功能 | 编码氨基酸 |
|---|---|---|
| AUG | 起始密码子 | 甲硫氨酸 |
图片来自于https://www.163.com/dy/article/FC1ERVVE0532AN5N.html
图片来自于百度百科
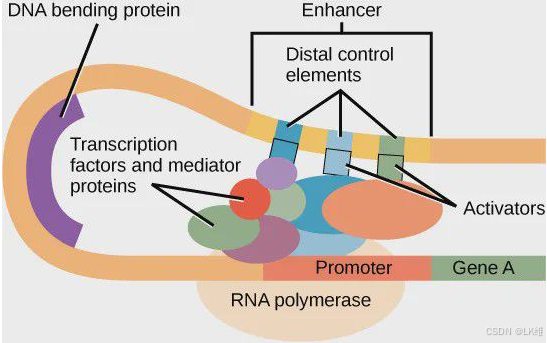
启动子(Promoter)
启动子是DNA上的一个特殊区域,通常位于基因的上游区域,它是RNA聚合酶识别并结合的位点,从而启动基因的转录过程。启动子区域包含多个转录因子结合位点,这些转录因子帮助RNA聚合酶正确地识别和结合到启动子上。启动子的强度可以影响基因表达的水平,强启动子可以驱动更高水平的基因表达。
增强子(Enhancer)
增强子是DNA上的另一个非编码区域,可以位于基因的上游、下游或甚至基因内部。增强子通过与特定的转录因子结合,增强基因的转录效率。增强子的作用不依赖于其与基因的物理距离,它们可以跨越较远的DNA区域与启动子相互作用,影响基因的表达。增强子的活性可以是组织特异性的,意味着它们在某些细胞类型中活跃而在其他细胞类型中不活跃。
特点
① 在转录起始点5’或3’侧均能起作用;
② 相对于启动子的任一指向均能起作用;
③ 发挥作用与受控基因的远近距离相对无关;
④ 对异源性启动子也能发挥作用;
⑤通常具有一些短的重复顺序。
两种类型
⑴ 细胞特异性增强子:能够在特定的细胞或特定的细胞发育阶段选择性调控基因转录表达的增强子称为细胞特异性增强子。例如,B细胞免疫球蛋白重链基因或轻链基因的增强子,只有在胚胎干细胞分化为B细胞时,才能对Ig基因起正调控作用。此外,α-类和β-类珠蛋白基因簇上游非编码区中均存在红细胞系特异性增强子。
⑵ 诱导性增强子:在特定刺激因子的诱导下,才能发挥其增强基因转录活性的增强子称为诱导性增强子。如激素反应元件(HRE)及金属应答元件(MRE)
超级增强子
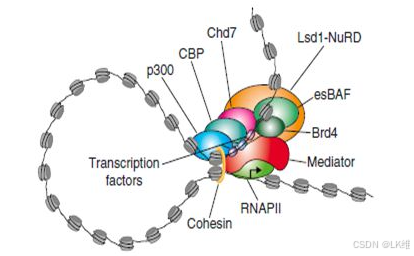
超级增强子是具有转录活性增强子的一个大簇, 富集高密度的关键转录因子(Master transcription factors)、辅因子(Cofactor)和增强子表观修饰标记(Histone modification marks)(见图1)。在功能上超级增强子能够驱动控制细胞身份基因的表达,可以用来解释细胞类型特异的表达模式,在发育生物学、癌症等疾病致病机理研究中显示出巨大的应用潜力。胚胎干细胞中的多个转录因子(Oct4、Sox2、Nanog、Klf4、Esrrb、Nr5a2、Prdm14、Tcfcp2l1、Smad3、Stat3、Tcf3)富集在超级增强子上,在之前的研究中发现这些转录因子在胚胎干细胞中起着十分重要的作用。
Richard A. Young曾激动地预言道:“‘超级增强子’具有广阔的研发前景和价值,必将成为下一个药物研发的黄金靶点!” 因此开展肿瘤相关超级增强子的研究,将有助深入解开肿瘤发病机制,并且可用于指导抗肿瘤药物的高效研发,具有重要的社会意义和经济价值。
转录因子(Transcription Factor, TF)
转录因子是一类蛋白质,能够结合到DNA上的特定序列(如启动子或增强子),并调节基因的转录。转录因子可以是激活因子,增强基因表达;也可以是抑制因子,降低基因表达。转录因子通过其DNA结合域识别特定的DNA序列,并通过其激活域招募或与RNA聚合酶及其他转录辅助因子相互作用,从而调节转录过程。
相互作用
转录因子、启动子和增强子之间的相互作用是复杂的。转录因子可以独立工作,也可以作为大型蛋白质复合体的一部分。增强子通过转录因子与启动子区域的相互作用,增强或抑制启动子的活性。这种相互作用可能涉及染色质重塑,即改变染色质的结构,使DNA更易于或更难于被转录机器访问。
这些调控元件的协同作用确保了基因在适当的时间和地点以适当的水平表达,这对于细胞的发育、分化和响应环境变化至关重要。异常的基因调控可能导致多种疾病,包括癌症和遗传性疾病。

















 1706
1706

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









