










Characterization and structure-based protein engineering of a regiospecific saponin acetyltransferase from Astragalus membranaceus
表征和基于结构的蛋白质工程:黄芪特异性皂苷乙酰转移酶,一篇乙酰基转移酶文章精读分享~
类似一篇综述 酰基转移酶综述-21年-地表最强系列-文献精读-3

摘要
乙酰化有助于许多具有药用重要性的天然产品的生物活性。然而,关于糖部分的乙酰化知之甚少。在这里,我们报道了来自黄芪的一种皂苷乙酰转移酶AmAT7-3。通过逐步的基因挖掘方法发现AmAT7-3,该酶被鉴定为黄芪皂苷IV(1)的木糖C3′/C4′-O-乙酰转移酶。为了阐明其催化机制,我们获得了AmAT7-3/1和AmAT7-3A310G/1的复合晶体结构,揭示了一个由特定序列AADAG决定的大型活性口袋。结合量子力学/分子力学(QM/MM)计算,AmAT7-3的区域特异性是由周围氨基酸(包括#A310和#L290)调节的糖定位决定的。此外,使用半理性设计构建了一个小型突变体库,其中变体A310G和A310W分别被发现能催化特定的C3′-O和C4′-O乙酰化。AmAT7-3及其变体也被用来乙酰化其他生物活性皂苷。这项工作扩展了对皂苷乙酰转移酶的理解,并为皂苷乙酰化提供了高效的催化工具。
引言
皂苷是一类分布广泛且结构复杂的天然产物,具有多种生物活性。乙酰化是皂苷最常见的修饰方式之一,对提高其生物活性起着重要作用。例如,来自皂皮树的乙酰化皂苷QS-7(在皂苷C-4上乙酰化)是一种受青睐的抗癌和抗病毒疫苗佐剂候选物,其在小鼠中的毒性几乎可以忽略不计。人参皂甙-Rs4(在葡萄糖C-6上乙酰化)与未乙酰化的人参皂甙-Rg5相比,显著诱导了SK-HEP-1细胞死亡。在齐墩果酸3-O-阿拉伯吡喃糖苷(C-3和C-4)上的乙酰化使得对A431细胞的细胞毒性提高了至少10倍。尽管全-O-化学酰化策略得到了高度发展,但由于羟基组的相对活性接近,糖部分的位点选择性乙酰化具有挑战性。以前的尝试主要集中在保护变旋异构基团上,而皂苷分子上的乙酰化仍然很少见。在QS-7的化学合成中,构建乙酰化的皂苷残基至少需要四个步骤,即C3-OH烷基化、C2-OH硅烷化、乙酰化和C3醚的去除,产率为47%。因此,使用化学催化剂合成多样化的天然发生的乙酰化皂苷具有挑战性。
皂苷的乙酰化很可能是由酰基转移酶催化的,特别是糖苷乙酰转移酶(ATs)。在这一组中功能鉴定的酶大多利用芳香酰或丙二酰基团作为供体。迄今为止,只有六种针对糖苷的乙酰转移酶被报道,其中四种催化黄酮糖苷C6′-OH(Vh3MAT1、Lp3MAT1和CcAT1/2)和两种修改三萜皂苷C4′-OH(QsACT1和SOAP10)。然而,由于酰基转移酶的快速分化,上述酶在我们的分析中或在以前的报告中显示出高度多样性。例如,皂苷ATs QsACT1和SOAP10的序列相似性仅为20.7%。另一方面,尽管已经报道了七个酰基转移酶的晶体结构,但其中没有一个属于能容纳更大尺寸的皂苷的糖苷ATs。关于糖苷ATs的催化机制和区域特异性知之甚少。因此,发现催化皂苷乙酰化的ATs并揭示其催化机制具有挑战性。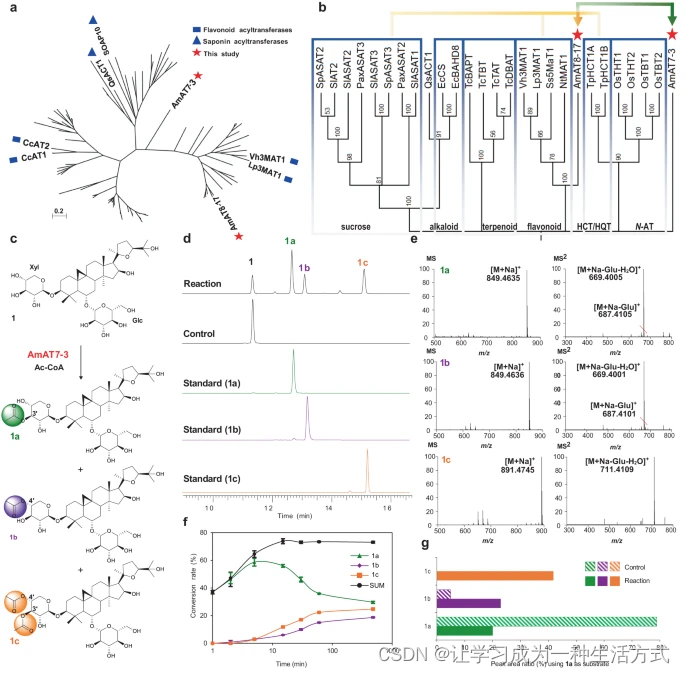
a 最大似然法构建的具有多种功能的乙酰转移酶树(补充数据1)。b 代表性酰基转移酶与AmAT7-3的最大似然树。在MEGA7中使用1000次自举重复进行了进化分析。黄色和绿色箭头分别指示第一轮和第二轮基因筛选。HCT/HQT,代表了香草酸或奎宁酸酰基转移酶。c AmAT7-3催化的黄芪皂苷IV(1)的乙酰化。d UPLC/MS反应、对照和标准样品的总离子流色谱图。e 产物1a、1b、1c的HR-MS和HR-MS/MS分析,正离子模式。f 使用1作为底物的AmAT7-3酶促反应的时间进程。数据以平均值±标准差呈现(n = 2或3次独立实验)。源数据提供为源数据文件。g 在使用1a作为底物的AmAT7-3反应混合物中,1a-1c的峰面积比。Glc表示葡萄糖基团;Xyl表示木糖基团;Ac-CoA表示乙酰辅酶A;反应:由AmAT7-3催化的乙酰化反应;对照:反应混合物中使用了煮沸的AmAT7-3。
黄芪皂苷是黄芪根的主要生物活性成分,黄芪根是一种源自黄芪(Astragalus membranaceus)的全球流行的草药。黄芪皂苷具有环烷烷型骨架,在其C3-木糖基团上进行多种乙酰化修饰,生成的化合物包括黄芪皂苷II(1d,C2′-O-Ac)、异黄芪皂苷II(1a,C3′-O-Ac)、环首皂苷II(1b,C4′-O-Ac)、黄芪皂苷I(1e,C2′,3′-O-Ac)、异黄芪皂苷I(C2′,4′-O-Ac)和乙酰黄芪皂苷I(C2′,3′,4′-O-Ac)。其中,黄芪皂苷I和II是黄芪根中最丰富的皂苷。黄芪皂苷具有多种生物活性,如心脏保护、神经保护和免疫调节效应,而乙酰化的数量和位置在很大程度上决定了它们的生物活性。由于黄芪根的化学复杂性,需要进行广泛的纯化才能从至少包含189种化合物的提取物中获得这些黄芪皂苷。因此,研究黄芪皂苷的生物合成途径以扩展这些皂苷的来源是必要的。尽管我们之前的研究已经确定了生物合成途径中的骨架形成的氧化鲨烯环化酶和四种糖基转移酶,下游的乙酰化步骤仍然不明确。基于特定的底物(三萜C3-木糖)和反应位点(C2′/C3′/C4′),制造黄芪皂苷的ATs应该与以前报道的糖苷ATs显著不同。
这项工作集中于使用黄芪根作为植物材料,发现并阐明皂苷乙酰转移酶的催化机制。首先,通过逐步的转录组分析发现了一种皂苷乙酰转移酶AmAT7-3,并对其进行了生化特征分析。其次,获得了AmAT7-3(PDB:8H8I)的复杂晶体结构,以深入了解催化机制,并采用半理性设计将AmAT7-3改造成区域选择性酶。还解析了工程变体AmAT7-3A310G(PDB:8HBT)的复杂晶体结构。最后,使用AmAT7-3及其变体修改了具有医药重要性的皂苷分子,从而增强了天然产品的结构多样性。这些结果扩展了对皂苷乙酰转移酶的蛋白质序列、催化机制和蛋白质工程的理解,并为糖苷乙酰化提供了有效的催化工具。
结果与讨论
基因筛选和功能表征
由于糖苷乙酰转移酶的序列多样性,采用了逐步的BLAST搜索来筛选黄芪皂苷乙酰转移酶。首先,我们选取了九种修饰六元环的乙酰转移酶作为模板(补充表1)。它们的酰基受体包括黄酮糖苷、香草酸和蔗糖。在黄芪的转录组(SRR923811)中进行BLASTn搜索,结果发现了16个候选基因(e < 10^-5),这些基因含有保守的基序HXXXD和DFGWG。随后,这些候选基因被克隆到pET-28a(+)载体中,并在大肠杆菌中表达(补充表2)。在包含30μg蛋白、0.1mM黄芪皂苷IV(1)、0.5mM乙酰辅酶A和0.5mM二硫苏糖醇的50mM Na2HPO4-NaH2PO4缓冲液(pH 6.0,100μL)中,对纯化的重组蛋白的功能进行了表征。反应在30°C下进行了30分钟。在这些候选物中,AmAT8-17(GenBank存取号OQ915518)在黄芪皂苷IV(1)的C4′-OH位置表现出乙酰化活性,转化率为25%(补充图1)。然而,除了其活性较差外,AmAT8-17无法催化C2′-O或C3′-O乙酰化,这些是黄芪皂苷的主要酰化修饰位点。因此,我们使用AmAT8-17作为模板进行了另一次BLASTn搜索,得到了10个额外的候选基因。其中,一个候选基因AmAT7-3(ON804888)被发现可以修饰黄芪皂苷IV(1),产生了三个极性较低的峰(1a,1b,1c),如图1所示。AmAT7-3的蛋白序列与其他酰基转移酶没有聚类,并形成了一个相对独特的分支,表明其具有高序列特异性(图1a,b)。AmAT7-3、AmAT8-17和六种已知的糖苷ATs的分布解释了仅基于蛋白质序列预测乙酰转移酶功能的挑战。
1a和1b的质谱显示其[M+Na]+离子为849.46 Da,比化合物1高出42.01 Da。1c的[M+Na]+离子比1高出84.02 Da。这些结果表明1a/1b为单乙酰化产物,而1c为双乙酰化产物(图1c-e)。1及其所有三个产物的MS/MS光谱显示了180.06 Da的中性丢失,与一个葡萄糖基团和一个水分子一致,表明乙酰化发生在木糖基团上。通过与参考标准比较,进一步确认了产物的结构。产品1a、1b和1c分别被鉴定为C3′-O、C4′-O和C3′, C4′-O乙酰化的黄芪皂苷IV(图1)。
AmAT7-3在30°C、pH 6.0(50 mM Na2HPO4-NaH2PO4缓冲液)条件下的30分钟反应中显示出最高活性,并且它的活性不依赖于二价金属离子(补充图2)。AmAT7-3的体外转化率达到了85.3%,高于许多之前表征的酰基转移酶。由于AmAT7-3能催化不同位点(C3′和C4′)的乙酰化,我们进行了480分钟的时间进程实验以确定其首选位点(图1d)。产品1a的转化率在最初2分钟内迅速增加,然后逐渐降至36.1%。相反,1b和1c的累积在60分钟内逐渐增加。三种产品的总转化率在30分钟达到平台期。这些结果表明AmAT7-3的首选催化位点为C3′-OH。化合物1b可能是AmAT7-3的直接产物,也可能是由于C3′和C4′上的邻近羟基而导致的1a的酰基迁移产物。为了澄清这一点,使用1a作为底物。与AmAT7-3共孵育时,生成了41.8%的1c(C3′,C4′-O乙酰化1)。相比之下,与煮沸的AmAT7-3共孵育时,未检测到1c(图1f)。这些结果表明AmAT7-3可以修饰1a的C4′位点。此外,从1a到1b的自发转化率约为6.1%,显著低于AmAT7-3催化产物中的1b/1a比率56.5%(图1f对比1c)。这表明AmAT7-3也可以催化1的C4′位点。基于1a的迅速生成,我们推断1c是通过C3′和C4′的顺序乙酰化产生的。
黄芪中黄芪皂苷的推测乙酰化途径
除了异黄芪皂苷II(1a)和环首皂苷II(1b)外,黄芪中的皂苷还包括黄芪皂苷II(1d)和I(1e),它们分别是1的C2′-和C2′, C3′-乙酰化产物(图2a)。由于我们筛选了26个候选基因并且在其中没有发现C2′-乙酰化活性,我们研究了AmAT7-3对C2′-乙酰化产品的贡献。首先,在烟草中确定了AmAT7-3的亚细胞定位,使用pSuper 1300-GFP载体(图2b)。GFP-AmAT7-3的荧光信号限制在细胞质中,表明AmAT7-3主要在活细胞的细胞质中表达。在豆科植物中,细胞质环境倾向于略呈碱性,例如绿豆悬浮培养细胞(pH 7.5)和蚕豆气孔保卫细胞(pH 7.67)。因此,我们在不同的pH条件(Na2HPO4-NaH2PO4,pH 6.0、7.0、8.0)下研究了AmAT7-3产物1a的稳定性,时间为90分钟(图2c)。结果表明,尽管在碱性环境下乙酰化皂苷易于水解
(分别为pH 6、7、8下的3.9%、27.7%、49.5%),但在pH 7和pH 8下C2′-乙酰化产品1d的比例显著增加。在未水解的产品中,1d在pH 6、7、8下的比例分别为4.8%、34.2%和63.5%。为了更好地理解自发的酰基迁移反应,在50 mM Na2HPO4-NaH2PO4,pH 7.6的条件下,使用1a/1b/1d(C3′/C4′/C2′-OAc)作为底物获得了时间进程。有趣的是,1a和1b在8小时后转化为1d作为主要的乙酰化产物(在乙酰化产品中为58.7-70.2%),而1d转化回1a/1b的部分较小(<25%)(图2d-f)。这些结果表明,AmAT7-3的主要产品1a/1b可以通过酰基迁移转化为黄芪中的主要皂苷1d。当AmAT7-3在烟草中过表达并与1共渗透作为底物时,也可以观察到1d的生成(图2g)。这进一步表明,植物体内的乙酰化产品与体外不同,AmAT7-3可能通过C3′/C4′-O乙酰化和酰基迁移在黄芪皂苷1d和1e的产生中发挥作用。黄芪中C2′-乙酰化皂苷的积累机制需要进一步研究。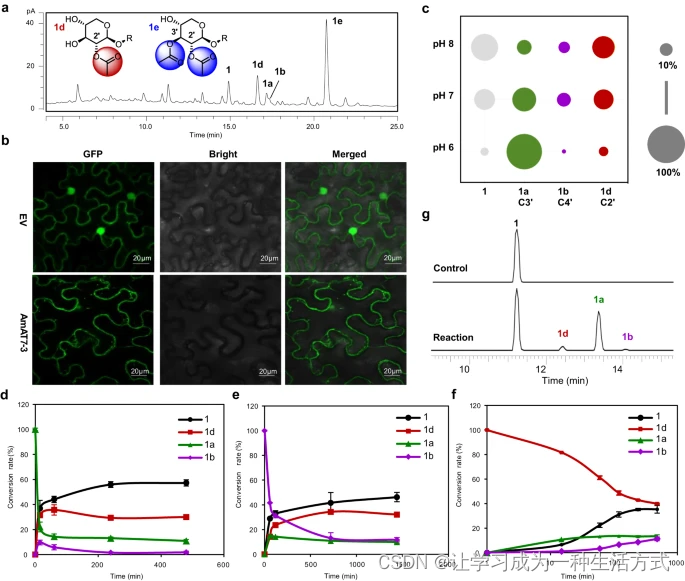
a 黄芪根的UHPLC/CAD色谱图。b AmAT7-3的亚细胞定位。三次独立实验显示了相似的结果。c 在不同pH值下1a降解产物的比例。数据以三次独立实验的平均值呈现。d–f 分别为1a、1b和1d的酰基迁移的时间进程。数据以平均值±标准差(n = 2或3次独立实验)呈现。源数据为(c–f)提供了源数据文件。g 在烟草(N. benthamiana)中AmAT7-3的瞬时表达。呈现了从农杆菌浸染叶片中提取的UHPLC/MS迹线。EV:空的pSuper 1300-GFP载体的荧光信号;反应:在烟草中瞬时表达AmAT7-3;对照:在烟草中瞬时表达空的pEAQ-HT载体。
晶体结构与AmAT7-3的位点特异性催化机制
AmAT7-3是一种罕见的多位点乙酰转移酶,能乙酰化大型皂苷分子。为了深入了解其乙酰化机制,我们解析了AmAT7-3/黄芪皂苷IV(1)复合体的晶体结构(PDB ID: 8H8I),分辨率为2.03 Å。据我们所知,之前尚未报道过皂苷酰基转移酶的晶体结构。我们还尝试获得含有乙酰辅酶A或辅酶A的三元复合体晶体结构,但未能成功。与先前报道的酰基转移酶类似,AmAT7-3的结构被划分为两个大致相等大小的域,其中有一个溶剂通道穿过蛋白质分子(图3a)。值得注意的是,与转移对香豆酰辅酶A至香草酸的SbHCT(PDB ID: 4KEC)的结构相比,AmAT7-3的活性口袋更大,以容纳皂苷分子(图3b-d)。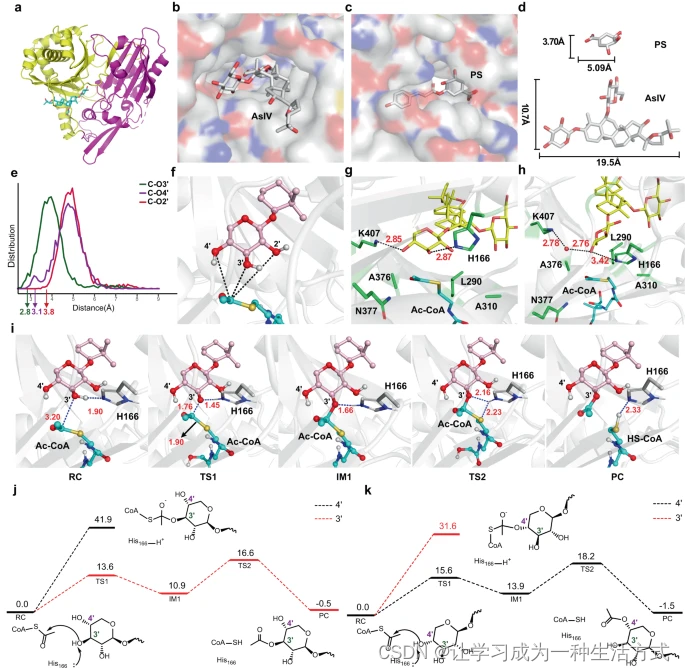
a 黄芪皂苷IV与AmAT7-3的复合晶体结构。b AmAT7-3的活性口袋表面。c SbHCT的活性口袋表面。d 黄芪皂苷IV与香草酸,分别为AmAT7-3和SbHCT的底物,分子尺寸比较。e 乙酰辅酶A的乙酰基C与黄芪皂苷IV糖基O位点之间的距离分布。f 乙酰基C与黄芪皂苷IV各糖基O位点之间的距离。g 分子动力学模拟中构象-1的反应瞬态及其与周围氨基酸的相互作用。h 构象-2的反应瞬态及其与周围氨基酸的相互作用。i 由AmAT7-3催化的O3′乙酰化反应的QM/MM优化反应状态。j 构象-1中AmAT7-3/黄芪皂苷IV/乙酰辅酶A的QM/MM计算能量剖面。k 构象-2中AmAT7-3/黄芪皂苷IV/乙酰辅酶A的QM/MM计算能量剖面。AsIV代表黄芪皂苷IV,PS代表对香豆酰香草酸,RC代表反应物复合体,TS1代表过渡状态1,IM1代表中间体1,TS2代表过渡状态2,PC代表产物复合体。能量以千卡/摩尔表示,距离以埃表示。
为了理解多位点乙酰化的机制,我们进行了分子动力学(MD)模拟,以研究底物1的结合和动态变化。由于无法通过对接获得合理的乙酰辅酶A的位置(补充图3),因此基于SbHCT的已知结构通过叠加将乙酰辅酶A打包到结构中。乙酰基C与各糖基O位点之间的距离分布显示在图3e中。观察到的最短距离为Cx-O3′(2.8 Å),Cx-O4′(3.1 Å)和Cx-O2′(3.8 Å),表明O3′和O4′位点可接近乙酰化,而O2′位点可能较不可接近。由于AmAT7-3的活性口袋相对宽敞,我们在整个MD模拟过程中观察到木糖基糖环的显著旋转运动。基于旋转角度,我们使用75度阈值将底物分为两个主要构象(补充图4)。在构象-1中,O3′和O4′位点分别与K407和H166形成氢键。这种氢键约束了底物,使O3′位点与Cx靠近,暗示这是一个潜在的选择性乙酰化位点(图3g)。随着糖环的旋转,它为溶剂水分子进入创造了空间,导致构象-2的出现。在这种构象中,O4′位点接近Cx,而O3′位点保持距离(图3h)。因此,乙酰化的区域选择性可以通过两个主要构象中底物的定位合理解释。
为了进一步支持上述预测,对构象-1或构象-2中接近攻击构象的Cx-O3′或Cx-O4′距离的两个反应瞬态进行了量子力学和分子力学(QM/MM)计算(补充图5-6)。根据先前的研究,邻近的His残基可能在乙酰化反应中充当酸碱催化剂。我们的计算显示,反应由糖基O原子向His166的质子转移启动,这与糖基O对乙酰基Cx的亲核攻击同时进行,导致四面体中间体的形成。随后,另一个质子从His166转移到Sx,促进了C-S断裂并产生了产品1a/1b(图3i,补充图7)。为构象-1和构象-2的两个反应瞬态计算的QM/MM能量剖面分别呈现在图3j和图3k中。检查这些能量剖面揭示了在构象-1中对O3′位点的乙酰化比O4′位点更受青睐,而在构象-2中则相反。MD预测的两种构象的比例与实验观察结果一致,表明O3′的乙酰化比O4′的乙酰化更受青睐(补充图4)。
工程化AmAT7-3的区域选择性及其突变体的立体选择性机制
为了进一步阐明邻近氨基酸如何影响木糖基(Xyl)组的构象并调节AmAT7-3的区域选择性,我们分析了AmAT7-3/黄芪皂苷IV(1)的复合结构。除了已确立的H166的关键作用(补充图8)外,发现距离底物5Å内的十个氨基酸参与构建活性腔的内部结构。这些氨基酸包括K407, R38, D311, A312, A310, L290, I34, I288, I374和A376(图4a,补充数据2),它们可能在决定底物构象中起决定性作用。为了研究它们的作用,构建了一个突变体库,选择了七个氨基酸作为替代残基,包括Ala/Val(非极性,脂肪族)、Trp/Phe(非极性,芳香族)、Gly(极性,无电荷)、Asp(酸性)和Arg(碱性)。使用纯化的蛋白质对突变体的功能进行了表征,采用与野生型相同的程序。黄芪皂苷IV(1)和乙酰辅酶A分别用作底物和酰基供体。有趣的是,几个突变体显著增强了对C3′-OH的区域选择性(图4b-d)。例如,L290V突变体特异性修饰C3′-OH,其转化率为79.1%,而野生型为35.1%。另一方面,四个突变体特异性修饰了1的C4′-OH,如A310W,其转化率为53.1%(野生型为19.8%)。更有趣的是,A310的突变体表现出不同的选择性(图4d)。A310D/G/R选择性催化1的C3′-OH乙酰化,而A310F/W特异性青睐C4′-OH。此外,我们分析了野生型和突变体的动力学参数(补充图9-10)。AmAT7-3A310G对1a的Km和kcat/Km值分别为76.7 μM和0.6 mM−1s−1,而AmAT7-3A310W对1b的为168.3 μM和0.19 mM−1s−1。与野生型相比,相似范围的动力学参数表明,这些突变改变了区域特异性,而不降低催化效率。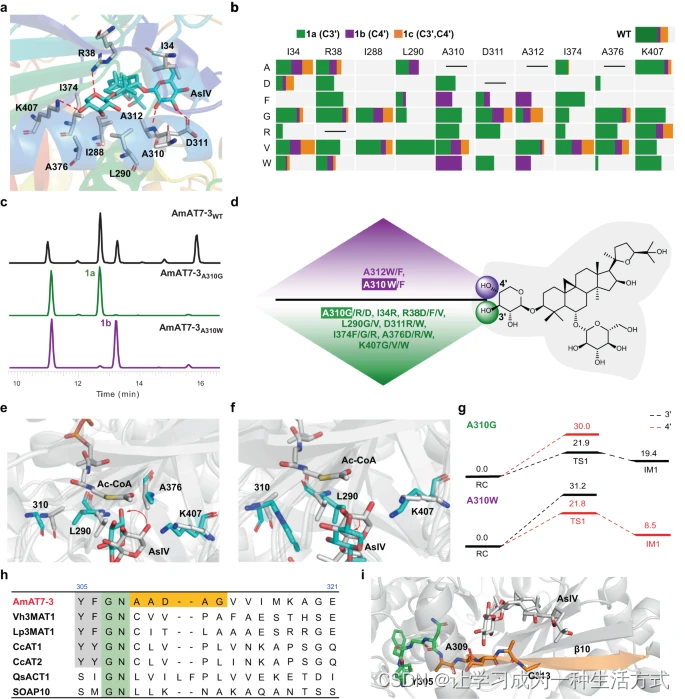
a 活性口袋中1周围5Å范围内的氨基酸。b AmAT7-3的突变体库及不同突变体产生的1a/1b/1c比例。c 由AmAT7-3及其突变体催化的反应样品的UHPLC/MS色谱图。d 具有特异性C3′-OH或C4′-OH乙酰化活性的突变体。e AmAT7-3A310G(青色)与野生型(来自构象-1,灰色)的QM/MM优化反应物复合体的叠加。f AmAT7-3A310W(青色)与野生型(来自构象-1,灰色)的QM/MM优化反应物复合体的叠加。g AmAT7-3A310G/W/1/Ac-CoA的QM/MM计算能量剖面(以千卡/摩尔为单位)。AsIV代表黄芪皂苷IV(1)。h AmAT7-3与其他黄酮或皂苷乙酰转移酶的序列比对。i AmAT7-3的β10桶中YFGN和AADAG基序的位置。RC代表反应物复合体,TS1代表过渡状态1,IM1代表中间体1。
a 化合物1-8的结构和生物活性。b 由AmAT7-3或化学试剂催化的6的乙酰化反应。PPT表示人参三醇,a/b/c表示单/双/三乙酰化产物。c 由AmAT7-3催化的乙酰化产物的转化率。O:通过与标准品比较确定;#:通过NMR制备并鉴定;*:以前未报道的化合物。d 由AmAT7-3A310G和AmAT7-3A310W催化的乙酰化产物的转化率。
除了木糖基团(1-2),AmAT7-3还能识别包括葡萄糖基(4-6)、阿拉伯糖基(3)和葡萄糖醛酸(7-8)在内的其他糖基(图5c)。制备了四种单乙酰化产物,其结构通过NMR光谱分析表征(补充图25-53,补充表3-6)。其中,3a、3b和7a以前未报道。NMR光谱显示所有四个产物中出现了新的碳信号(δC 171.0-171.3)和新的甲基信号(δH 1.97-2.11,s,3H),表明成功进行了乙酰化。乙酰化位点通过HMBC相关进一步确认。例如,在产物3a中,观察到H-3(δH 3.33)与C-2(δC 27.0)/C-4(δC 39.9)以及新出现的羰基碳(δC 171.3)之间的HMBC相关,从而确定化合物3a为C3′-O乙酰化的紫云英皂苷II。同样,产物6a、7a和3b分别被确认为C6′-OH、C3′-OH和C4′-OH的乙酰化产物。这些结果展示了AmAT7-3在与各种糖基及皂苷分子不同位点反应中的多功能性。根据配体相互作用模式的检查(补充图54),底物通用性背后的结构基础可能源于活性口袋的大体积,有助于容纳较大的皂苷分子。皂苷的定位可能由诸如R38和E39等保守氨基酸决定,这些氨基酸与糖基形成氢键。
虽然对于化合物6-8生成单一产品,但AmAT7-3催化的化合物2-5形成了2到3种产品,类似于观察到的化合物1的反应。为了提高区域选择性,我们使用区域选择性突变体AmAT7-3A310G和AmAT7-3A310W来催化2-5的乙酰化(图5d)。正如预期的那样,突变体显示出比野生型酶更高的区域选择性。例如,野生型酶对5的转化率为23.1%,包含两种产品比例为14:9。A310G变体催化5生成单一产品,转化率为96.3%。这些发现进一步确认了310位点残基在塑造活性腔并突出区域特异性乙酰化在未来制备单乙酰化产物中的潜力的关键作用。
总之,我们使用逐步BLAST方法表征了来自药用植物黄芪的皂苷乙酰转移酶AmAT7-3。我们解析了AmAT7-3及其变体A310G与黄芪皂苷IV的复合体晶体结构。AmAT7-3显示出与报道的乙酰转移酶不同的独特蛋白序列,并且是少有的具有晶体结构的皂苷乙酰转移酶。它可以催化黄芪皂苷IV(1)的C3′/C4′-O乙酰化以及其他七种药用重要皂苷的乙酰化。总共制备了四种产物,其中三种以前未报道过。结构分析揭示了AmAT7-3具有由特定序列AADAG决定的大型活性口袋,其区域选择性由关键氨基酸如A310、L290、K407、D311和A312通过糖环在活性口袋中的定位控制。为了增强其区域特异性,我们基于半理性设计构建了一个小型突变体库。变体A310G和A310W分别被发现可以催化黄芪皂苷IV(1)的特异性C3′-O和C4′-O乙酰化。此外,这些突变体在修改生物活性皂苷2-5时显示出更高的区域特异性。这项工作提供了糖苷乙酰转移酶的蛋白序列、催化机制和半理性工程的洞察,并提供了高效的皂苷乙酰化催化工具。
方法
通用方法
从成都慕斯生物技术有限公司购买了以下化合物:黄芪皂苷IV(1),紫云英皂苷II(3),人参皂苷Rh1(6),甘草次酸3-O-β-D-葡萄糖苷(7),筑泽皂苷IVa(8),异黄芪皂苷II(1a),环头皂苷II(1b),3-O-3′,4′-二乙酰基-β-D-木糖吡喃基-6-O-β-D-葡萄糖吡喃基环黄芪甲素(1c),黄芪皂苷II(1d)和黄芪皂苷I(1e)。黄芪皂苷VII(2),牛蒡苷3-O-β-D-葡萄糖苷(4)和蟾蜍苷3-O-β-D-葡萄糖苷(5)按先前报道的方法制备27,37,43。乙酰辅酶A从上海柱子生物科技有限公司购买。甲醇和乙腈(UPLC级)从美国费希尔科学公司购买。
植物材料
黄芪(A. membranaceus)种子从中国河北省安国市丰华种子站购买。根据先前发表的方法27培育两周后获得幼苗,并用于基因克隆。在中国黑龙江省大兴安岭采集黄芪根样本,用于化学分析。
系统发育分析
采用基于JTT矩阵的最大似然方法推断进化历史44,并在MEGA7软件中进行进化分析45。启发式搜索的初始树通过应用邻接连接和BioNJ算法自动获得,以估算使用JTT模型的成对距离矩阵。选择了对数似然值最优的拓扑结构。自举值为1000。
基因克隆和蛋白表达
使用TranZolTM试剂盒(中国全式金生物科技公司)提取总RNA,并使用FastQuant RT试剂盒(中国天根生物科技公司)逆转录为cDNA。设计特异性引物来扩增AmAT7-3和AmAT8-17,随后将其克隆到pET-28a(+)载体中(美国Invitrogen公司)。使用补充数据2中的特异性引物和Fastpfu Fly菌落试剂盒(中国全式金生物科技公司)构建突变质粒。测序后,将重组质粒转化到大肠杆菌BL21(DE3)(中国全式金生物科技公司)。在37°C下培养细菌株于加有50μg/mL卡那霉素的LB培养基中,直到OD600达到0.6。然后加入异丙基-β-D-硫代半乳糖苷(IPTG,0.1 mM)诱导蛋白表达,并在16°C下继续培养20小时。
蛋白纯化与结晶
通过超声破碎并离心去除碎片后,用Ni-NTA柱(中国全式金生物科技公司的Proteinlso Ni-NTA树脂)纯化上清,并使用Amicon Ultra-15 Ultracel-30K离心滤器(Merck Millipore)浓缩蛋白。使用特异性引物将AmAT7-3进一步克隆到pET28a-TEV载体中,并通过GE healthcare的SuperdexTM柱进行尺寸排阻色谱纯化。使用TransGen Biotech的蛋白定量试剂盒测定蛋白浓度,并通过SDS-PAGE检查纯度(补充图4)。
纯化的蛋白(AmAT7-3和AmAT7-3A310G各20 mg/mL)与5 mM乙酰辅酶A和5 mM黄芪皂苷IV(1)在冰上孵育1小时。AmAT7-3/1和AmAT7-3A310G/1的晶体通过悬挂滴蒸发法准备。在含有1μL蛋白溶液和1μL储备液(1.6 M硫酸镁,100 mM MES/氢氧化钠pH 6.5)的悬挂滴中获得立方晶体,3周后收集。在上海同步辐射设施的BL19U1光束线上进行晶体衍射和数据收集,使用HKL-2000程序套件。通过分子替换法构建初始晶体学相位,基于alphafold 2预测的AmAT7-3结构。使用CCP4、Wincoot和Phenix进行蛋白模型构建和细化。结构的详细信息列在补充表7中。使用Pymol显示晶体结构中配体的Fo-Fc遗漏图(补充图55)。
UHPLC/MS分析酶促反应产物
为了表征催化活性,通过在30°C下将30μg纯化蛋白、0.1 mM黄芪皂苷IV(1)、0.5 mM乙酰辅酶A和0.5 mM二硫苏糖醇混合在50 mM Na2HPO4-NaH2PO4缓冲液(pH 6.0)中反应30分钟,制备100μL反应混合物。用200μL甲醇终止反应,浓缩至干,重新溶解于200μL甲醇,并离心21130g 20分钟。将所得上清液用ThermoFisher Scientific的Vanquish UHPLC系统与带有加热电喷雾电离源的Q-Exactive四极杆-轨道阱质谱仪联用进行UHPLC/MS检测。样品在Waters T3柱(2.1×100 mm, 1.8μm)上使用0.1%甲酸(体积分数,A)和乙腈(B)在50°C下分离。使用梯度程序:0-1分钟,8% B; 1-19分钟,8-47% B; 10-16分钟,47-65% B; 16-19分钟,65-100% B; 19-22分钟,100% B。流速为0.3 mL/min。MS分析在正负离子模式下进行,参数为:喷雾电压:±3.5 kV;毛细管温度:350°C;剪切气体:45 arb;辅助气体:10 arb;探头加热器温度:400°C;S-lens RF水平:60 V;分辨率:全MS 70000,MS/MS 17500;扫描范围:m/z 100-1200;阶梯NCE:35 eV。数据使用XcaliburTM 4.1软件处理(Thermo Fisher Scientific)。使用HPLC(SSI-III)联用ELSD 6000(Alletch Chrom, USA)分离反应混合物。



















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









