










Discovering a mitochondrion-localized BAHD acyltransferase involved in calystegine biosynthesis and engineering the production of 3β-tigloyloxytropane
发现一个定位于线粒体的BAHD酰基转移酶,参与打碗花精生物合成,并工程化生产惕佫酰假托品
同样还有一篇酰基转移酶综述 酰基转移酶综述-21年-地表最强系列-文献精读-3
类似的酰基转移酶文章 表征和基于结构的蛋白质工程:黄芪特异性皂苷乙酰转移酶-文献精读14

摘要
茄科植物通过3α-和3β-托品醇的酯化生成托品烷生物碱(TAs)。尽管已知Littorine合酶负责3α-托品醇的酯化,进而导致莨菪碱的生物合成,但与3β-托品醇酯化相关的基因尚不清楚。在此,我们报告了来自颠茄的一个BAHD酰基转移酶,惕佫酰假托品合酶(TS),催化3β-托品醇和巴豆酰辅酶A(tigloyl-CoA)形成惕佫酰假托品,这是打碗花精生物合成的关键中间体,也是治疗神经退行性疾病的潜在药物。与其他定位于细胞质的BAHD酰基转移酶不同,TS定位于线粒体。通过分子对接和定点突变揭示了TS的催化机制。随后,在烟草中合成了惕佫酰假托品。发现一种细菌CoA连接酶(PcICS)可以合成巴豆酰辅酶A,这是惕佫酰假托品生物合成的酰基供体。通过表达TS突变体和PcICS,工程化的大肠杆菌可以从巴豆酰和3β-托品醇合成惕佫酰假托品。这项研究有助于表征托品烷生物碱的酶学和化学多样性,并提供了一种生产惕佫酰假托品的方法。
引言
植物次生代谢产物作为药物被人类利用已有数千年历史。托品烷生物碱(TAs)是一类次生代谢产物,其特点是具有包含一个含氮桥的环庚烷环的8-氮杂二环[3.2.1]辛烷核心骨架,通常被称为托品烷部分或托品烷核心1。已从茄科、旋花科、红树科、古柯科及其他科发现了300多种TAs2。这些化合物包括由茄科某些属合成的抗胆碱药物莨菪碱和东莨菪碱,以及由古柯科合成的麻醉药物可卡因3。
最近对茄科植物颠茄的研究全面揭示了莨菪碱和东莨菪碱的生物合成途径4,5。莨菪碱和东莨菪碱的托品烷部分由3α-托品醇(也称为托品醇)贡献,托品醇由托品酮还原酶I(TRI)使用托品酮作为底物生物合成6。此外,托品酮可以通过托品酮还原酶II(TRII)还原为3β-托品醇(也称为假托品醇)(图1)。打碗花精是一种由3β-托品醇衍生的TAs,广泛分布于植物界,包括茄科中的一些重要蔬菜,如西红柿、马铃薯、茄子和辣椒6,7,8,9。打碗花精强烈抑制糖苷酶,可能会导致营养吸收问题,可用于代谢障碍的治疗10,11,12,13。尽管在生产TAs的植物中是次要成分,一些3β-托品醇酯在药学上具有重要意义14,15。例如,惕佫酰假托品(也称为惕佫列定或托品林)是治疗神经退行性疾病的阿托品替代品;与阿托品相比,惕佫酰假托品引起的副作用更少,包括喉咙干燥、瞳孔散大和头痛16,17,18。此外,3β-苯甲氧基托品烷(也称为莨可卡因)是一种潜在的眼科和脊髓麻醉剂19。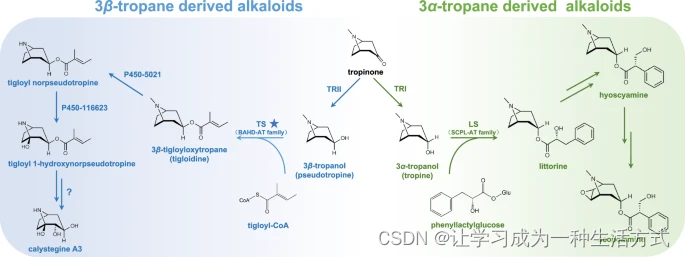
TRI:托品醇生成还原酶或托品酮还原酶I,TRII:假托品醇生成还原酶或托品酮还原酶II,LS:Littorine合酶,TS:惕佫列定合酶,本研究的关键发现。3β-托品烷生物碱的生物合成途径以蓝色标记,3α-托品烷生物碱的生物合成途径以绿色标记。P450-5021:惕佫酰假托品脱甲基酶。P450-116623:巴豆酰去伪莨菪氧化酶。
关于莨菪碱、东莨菪碱和可卡因的生物合成途径的综合知识极大地促进了它们在工程植物和微生物中的生产5,20,21,22。然而,对3β-托品醇衍生物生物合成的了解有限。因此,利用代谢工程或合成生物学方法高效生产有价值的3β-托品醇酯(如惕佫酰假托品和3β-苯甲氧基托品烷)的能力也受到限制。从结构上推测,打碗花精可以直接从3β-托品醇脱甲基和羟基化23,24,25。然而,最近的工作挑战了这一长期以来的推测,并提出了以下路线图:3β-托品醇被酯化为惕佫酰假托品(打碗花精生物合成的关键中间体),随后经过脱甲基和羟基化反应,然后进行水解反应,最终生成打碗花精26。尽管Robins及其同事在20世纪90年代报道了与3β-托品醇酯化相关的催化活性出现在曼陀罗和颠茄中27,28,但负责3β-托品醇酯生成的酰基转移酶的基因仍需进一步研究。
酯化是小分子最重要的化学修饰之一,涉及添加酰基以生成酯和酰胺。两类酶家族,丝氨酸羧肽酶样酰基转移酶(SCPL-AT)和BAHD酰基转移酶(BAHD-AT),作为主要的酰基转移酶,参与植物代谢物的酯化29。SCPL-ATs通常使用1-O-β-葡萄糖作为酰基供体29。例如,在莨菪碱和东莨菪碱的生物合成中,SCPL-ATLittorine合酶(LS)通过酯化催化3α-托品醇和苯乳糖的缩合生成Littorine30(图1)。有趣的是,最近的一项研究表明,紫松果菊中的一种SCPL-AT菊酸合酶(EpCAS)使用绿原酸作为酰基供体31。然而,BAHD-ATs通常使用酰基-CoA硫酯。例如,古柯植物中的可卡因合酶(EcCS),作为BAHD-AT家族的一员,通过催化甲基外喹宁(带有托品烷部分的中间体)与苯甲酸之间的酯化反应,负责可卡因(一种古柯树生产的托品烷生物碱)的生物合成32。
SCPL-ATs和BAHD-ATs独立进化33。代谢中的一个迷人方面是复杂的细胞内分隔。许多次生代谢物生物合成相关的酶位于细胞质和多个细胞器中。SCPL-ATs通常定位在液泡中,这种定位对其功能至关重要5,20。一般来说,BAHD-ATs没有定位信号,自由存在于细胞质中29。参与莨菪碱和东莨菪碱生物合成的酶分布在细胞质、液泡和内质网中5。有趣的是,尽管线粒体是氮代谢的中心,而托品烷生物碱是含氮次生代谢物34,但尚未报道涉及托品烷生物碱合成的酶在线粒体中的定位。
在本研究中,我们功能性地鉴定了一种来自颠茄的线粒体定位的BAHD酰基转移酶,惕佫酰假托品合酶(TS)。这种酶催化3β-托品醇和巴豆酰辅酶A(tigloyl-CoA)生成惕佫酰假托品(图1)。我们还分析了TS的组织表达模式,观察了其亚细胞定位,研究了其在植物中的代谢作用,并分析了其催化活性和机制。最终,在工程化大肠杆菌中高效生产了惕佫酰假托品。TS的鉴定丰富了关于托品烷生物碱生物合成的知识,并通过合成生物学提供了一种生产3β-托品醇衍生生物碱的生物技术方法。
结果
TS的表达与颠茄中惕佫酰假托品的分布相关
为了鉴定负责惕佫酰假托品生物合成的基因,我们首先分析了颠茄不同器官中惕佫酰假托品的水平。惕佫酰假托品在颠茄的初生根和次生根中被检测到,而在颠茄的其他组织中(包括成熟种子、成熟果实、花、茎、青果和叶)未检测到(图2A)。在初生根和次生根中分别检测到1.18 μg.g−1干重(DW)和12.40 μg.g−1 DW的惕佫酰假托品。惕佫酰假托品的根特异性分布表明相应的基因表现出相似的表达模式,即根特异性或根优先。通过HMMER搜索分析,从颠茄转录组中鉴定了46个BAHD-AT基因。随后,利用代谢物和基因表达关联分析(MGAA)筛选出生成惕佫酰假托品的候选基因。MGAA结果显示,一个被命名为TS的推定BAHD基因(aba locus 5896)在初生根和次生根中特异性表达,因此与惕佫酰假托品紧密聚集(图2B)。在分离全长cDNA序列后,通过qPCR确认了TS的表达模式(图2C)。

A. 惕佫酰假托品的组织分布。3株独立植物用于惕佫酰假托品的组织分布分析。数据以均值±标准差表示,P = 0.0032。DW,干重。 B. 代谢物与BAHD基因家族表达模式的关联分析。 C. 不同器官中TS的相对表达水平,由qPCR检测。3株独立植物用于TS的相对表达分析。数据以均值±标准差表示,P = 0.0007。 D. BAHD-AT基因家族的系统发育分析。系统发育树分为0–6个分支。 E. 第3分支中的BAHD酰基转移酶及其底物:TS,本研究中鉴定的BAHD酰基转移酶;EcCS,古柯植物的可卡因合酶,古柯树的BAHD酰基转移酶;EcBAHD8,古柯植物中EcCS的同源物;PaxASAT1-4,矮牵牛中的酰基糖酰基转移酶;SlyASAT1-3,番茄中的酰基糖酰基转移酶;SsiASAT1-3,角堇中的酰基糖酰基转移酶;ShaASAT2,野生番茄中的酰基糖酰基转移酶;LeSAT1,紫草中的紫草素O-酰基转移酶;LeAAT1,紫草中的紫草素O-酰基转移酶;CrMAT,长春花中的minovincinine-19-羟基-O-乙酰基转移酶;CrDAT,长春花中的去乙酰长春碱4-O-乙酰基转移酶。所有功能性鉴定的BAHD-ATs均标注了其对应的登录号。统计分析根据双侧独立样本t检验进行。
BAHD酰基转移酶的系统发育分析
为了确定TS的进化关系,构建了包括颠茄的46个BAHD-AT和来自其他植物的196个功能性鉴定的BAHD-AT的系统发育树(图2D及补充数据1–2)。系统发育树分为0–6个分支,这些分支与先前的报道相同35。系统发育分析显示,TS聚集在第3分支(图2D)。在第3分支中,包括TS和一些功能性鉴定的BAHD-ATs,如PaASAT1-4,SlyASAT1-3,LeSAT1,LeAAT1,CaMAT和CaDAT(图2E)。这些BAHDs识别短链酰基-CoA硫酯,包括乙酰-CoA、异丁酰-CoA和异戊酰-CoA,作为酰基供体36,37,38,39。尽管如此,EcCS和EcBAHD8也包含在第3分支中(图2E);这两种BAHD酶使用带有3β-莨菪环骨架的甲基外奎宁作为酰基受体催化可卡因的生成32。因此,这些结果表明TS识别带有3β-莨菪环骨架的酰基受体和短链酰基-CoA硫酯的酰基供体。
TS催化3β-托品醇的酯化
为了研究TS的功能,我们在工程化的大肠杆菌中生产重组TS蛋白并纯化。首先,我们使用化学方法合成了3β-托品醇酯和巴豆酰-CoA(补充图S1–S24及补充数据3)。在包含3β-托品醇和巴豆酰-CoA的反应系统中,TS生成的产物具有[M + H]+ m/z为224.1645,与正品惕佫酰假托品一致(图3A, B)。然而,当TS被煮沸时,未检测到产物(图3A)。在包含3α-托品醇和巴豆酰-CoA的反应系统中,TS未生成产物(图3A)。上述结果表明,TS通过酯化反应催化3β-托品醇与巴豆酰-CoA缩合生成惕佫酰假托品。

A. TS酶促反应实验,使用巴豆酰-CoA作为酰基供体和3β-托品醇作为酰基受体。 B. 惕佫酰假托品的质谱(MS)数据。 C. VIGS-TS颠茄幼苗中TS的相对表达水平。P < 0.0001。 D. VIGS-TS颠茄幼苗中3β-托品醇的含量。P = 0.0024。 E. VIGS-TS颠茄幼苗中惕佫酰假托品的含量。P < 0.0001。 F. VIGS-TS颠茄幼苗中3β-乙酰氧基托品烷的含量。P < 0.0001。 G. VIGS-TS颠茄幼苗中巴豆酰去伪莨菪碱的含量。P < 0.0001。 H. VIGS-TS颠茄幼苗中巴豆酰1-羟基去伪莨菪碱的含量。P < 0.0001。 I. VIGS-TS颠茄幼苗中打碗花精A3的含量。P = 0.0002。对照组为通过空质粒转化获得的对照株。VIGS-TS为TS沉默株。每组各使用了15株独立植物进行VIGS实验。箱形图的中线表示中位值;下界和上界分别表示第一和第三四分位数;须线延伸至最小值和最大值。 J. TS过表达的颠茄毛状根培养物中TS的相对表达水平。P < 0.0001(OE-1),P < 0.0001(OE-2),P < 0.0001(OE-3),P < 0.0001(OE-4)。 K. TS过表达的颠茄毛状根培养物中3β-托品醇的含量。 L. TS过表达的颠茄毛状根培养物中惕佫酰假托品的含量。P = 0.0007(OE-1),P = 0.0024(OE-2),P = 0.0044(OE-3),P = 0.0025(OE-4)。 M. TS过表达的颠茄毛状根培养物中3β-乙酰氧基托品烷的含量。*P = 0.0136(OE-1),P = 0.0087(OE-2),P = 0.0048(OE-3),P = 0.0039(OE-4)。 N. TS过表达的颠茄毛状根培养物中巴豆酰去伪莨菪碱的含量。 O. TS过表达的颠茄毛状根培养物中巴豆酰1-羟基去伪莨菪碱的含量。 P. TS过表达的颠茄毛状根培养物中打碗花精A3的含量。对照组(CK)为转化了pBI121的8个独立转化根培养物。OE表示所有过表达TS的独立转化根培养物(每条线的三次生物重复),包括OE-1、OE-2、OE-3和OE-4。数据以均值±标准差表示。统计分析根据双侧独立样本t检验进行。DW,干重。
此外,研究了TS催化的最佳pH和温度条件。在pH范围6.0–10.6内,检测了3β-托品醇和巴豆酰-CoA处理的TS的催化活性。在pH 8.6时检测到TS的最大活性(补充图S25A)。大多数BAHD蛋白的最佳pH条件在8到10的狭窄碱性范围内。例如,EcCS、CiHCT2和NtMAT1的最佳pH条件分别为9.4、9.0和8.532,40,41。此外,确定了在20°C到50°C之间处理3β-托品醇和巴豆酰-CoA的TS的催化活性(补充图S25B)。在30°C时检测到TS的最大活性。接下来,在最佳条件(pH 8.6和30°C)下测量了酶动力学常数。
BAHD-AT家族蛋白通常表现出底物宽容性32,42,43,44,45。因此,我们测试了TS的不同酰基供体。当使用3β-托品醇和乙酰-CoA作为底物时,TS生成了[M + H]+ m/z值为184.1332的3β-乙酰氧基托品烷(补充图S26)。当使用3β-托品醇和苯甲酰-CoA作为底物时,TS生成了[M + H]+ m/z值为246.1489的3β-苯甲氧基托品烷,但产量极少(补充图S27)。当使用巴豆酰-CoA、乙酰-CoA和苯甲酰-CoA作为酰基供体时,TS对3β-托品醇的Km值分别为0.36 mM、0.39 mM和0.43 mM(表1)。此外,这些结果之间没有显著差异。然而,TS对巴豆酰-CoA、乙酰-CoA和苯甲酰-CoA的Km值分别为0.02 mM、0.09 mM和0.92 mM,差异显著(表1)。这些结果表明,TS对巴豆酰-CoA的亲和力高于乙酰-CoA或苯甲酰-CoA。TS对巴豆酰-CoA的催化效率(Kcat/Km)为338332.70 M−1.S−1,比对乙酰-CoA和苯甲酰-CoA的催化效率分别高出743倍和26473倍(表1)。酶促实验表明,TS主要催化惕佫酰假托品的生成。
沉默TS扰乱了植物体内打碗花精A3的生物合成
为了确定TS在植物体内3β-托品醇酯生物合成中的作用,通过病毒诱导的基因沉默(VIGS)在颠茄幼苗中抑制TS。如图3C所示,TS在颠茄根中被有效抑制。与TS表达水平下降一致的是,TS沉默株中的3β-托品醇、3α-托品醇和托品醇糖苷含量显著高于对照株(图3D和补充图S28)。TS沉默株中的惕佫酰假托品、3β-乙酰氧基托品烷、巴豆酰去伪莨菪碱、巴豆酰1-羟基去伪莨菪碱和打碗花精A3的含量显著低于对照株(图3E-I)。然而,TS沉默植物中的3α-托品醇衍生物Littorine、莨菪碱和东莨菪碱的含量与对照植物相比没有显著差异(补充图S28)。植物体内和体外实验均表明,TS在打碗花精A3的生物合成途径中催化惕佫酰假托品的生成。
TS过表达增加了毛状根中惕佫酰假托品的含量
此外,我们在颠茄毛状根培养物中过表达TS,以进一步研究其在惕佫酰假托品生物合成中的作用,并探索其在工程中的应用。基因组PCR和qPCR分析表明,TS整合到了颠茄基因组中,并且在TS过表达根系中的表达水平显著增加(图3J,补充图S29)。TS过表达株中的惕佫酰假托品和3β-乙酰氧基托品烷的含量显著高于对照株(图3L, M),而3β-托品醇、巴豆酰去伪莨菪碱、巴豆酰1-羟基去伪莨菪碱和打碗花精A3的生产没有差异(图3K, N-P)。此外,TS过表达株中的3α-托品醇衍生物的含量与对照株相比没有显著差异(补充图S30)。总体而言,这些结果表明,TS过表达可以增加颠茄毛状根中惕佫酰假托品的生产。
TS定位于线粒体
先前的研究报告指出BAHD-ATs定位于细胞质29,31,46。在分析TS中的靶向信号序列时,我们确定TS在其N端含有一个潜在的线粒体定位信号序列(补充图S31)。随后在烟草原生质体中研究了TS的亚细胞定位。TS与YFP(TS-YFP)融合及其线粒体定位信号肽(N端的32个氨基酸)与YFP(N32-YFP)融合后在烟草原生质体中表达。YFP信号与线粒体特异性荧光染料MitoTracker Red的信号重叠(图4A)。此外,我们对不含N端32个氨基酸的TS进行了亚细胞定位分析。结果表明,不含32个氨基酸的TS(TSDel-N32)仍表现出线粒体定位(补充图S32)。因此,我们推测TS的线粒体定位信号不仅位于其N端,还分布在其他区域47,48,49,50,51。当TSDel-N32在大肠杆菌中表达时,其催化活性显著下降(补充图S33A),表明TS的32个氨基酸序列在其酶活性中起着关键作用。根据AlphaFold2生成的TS三维结构,其32个氨基酸序列构成了TS的核心支架(补充图S33B)。

A. 使用共聚焦显微镜观察TS-YFP和N32-YFP在烟草原生质体中的定位。YFP黄色荧光来自YFP。MitoTracker Red,标记线粒体的MitoTracker Red荧光。Merge YFP + RED,为黄色荧光和MitoTracker Red荧光的合并图像。Chlorophyll,叶绿素自发荧光。Bright,明场图像。上述所有通道的重叠图像被合并。TS-YFP,TS与YFP融合。N32-YFP,TS N端32个氨基酸的线粒体定位信号肽与YFP融合。所有烟草转化和显微分析均在不同植物上独立进行三次。 B. 通过Western blot分析使用VDAC抗体检测颠茄线粒体。Western blot分析在不同植物上独立进行两次。 C. 来自颠茄线粒体的粗蛋白催化巴豆酰-CoA和3β-托品醇生成惕佫酰假托品。
接下来,我们从颠茄的次生根中分离出线粒体和上清液。电压依赖性阴离子通道(VDAC)用于检测线粒体52,53,54。线粒体标记蛋白VDAC在线粒体部分中检测到,但在上清液部分中未检测到(图4B)。这些结果表明我们成功从颠茄中获得了线粒体。随后检测了线粒体和上清液粗蛋白的催化活性。惕佫酰假托品的生成由线粒体粗蛋白催化,但上清液中的粗蛋白没有催化活性(图4C)。
这些结果表明TS定位于线粒体;这种细胞内隔离与先前报道的定位于细胞质的BAHD-ATs不同29。
TS的催化机制及其催化活性的改进
已经广泛研究了以苯丙素为酰基受体的BAHD-ATs的催化机制55,56,57,58;然而,BAHD-ATs识别结构与苯丙素截然不同的3β-托品醇作为酰基受体的机制仍然未知。为了探讨TS的催化机制并为工程提供信息,通过分子对接构建了一个复合模型(补充图S34)。我们观察到HXXXD基序中的His162与3β-托品醇的氧形成了一个氢键(3.0Å),这是BAHD-ATs中普遍保守的催化残基,表明His162是TS催化口袋中的碱性催化剂(图5A)。与我们的假设一致,当His162突变为丙氨酸时,TSH162A完全失去了催化活性(图5B)。此外,观察到由His162、Ile35、Gln39、Asn298、Leu300、Tyr280和Trp340形成的3β-托品醇结合口袋(图5C)。为了检验这些残基在结合口袋中的作用,生成了TS的定点突变体。Ile35、Gln39、Tyr280、Asn298、Leu300和Trp340突变为丙氨酸后,突变体的催化活性显著降低(图5D)。综上所述,我们的研究发现了3β-托品醇结合口袋中的这些残基在调控TS活性中起关键作用。

A. 含有3β-托品醇和巴豆酰-CoA的催化口袋示意模型。 B. 使用巴豆酰-CoA作为酰基供体时,TS和TSH162A的相对活性比较。 C. 与3β-托品醇结合的底物口袋中的关键残基。 D. 使用巴豆酰-CoA作为酰基供体时,TS和TS突变体(3β-托品醇结合残基突变为丙氨酸)的相对活性比较。P < 0.0001(TSI35A),P < 0.0001(TSQ39A),P < 0.0001(TSY280A),P < 0.0001(TSN298A),P < 0.0001(TSL300A),P < 0.0001(TSW340A)。 E. 3β-托品醇和酰基供体在TS催化下转化为3β-托品醇酯。数据以均值±标准差表示。重组蛋白从TS和每个突变体的三株独立转化体中获得并进行活性测试。统计分析根据双侧独立样本t检验进行。
酶动力学分析结果表明,TS对苯甲酰-CoA的利用效率低,表明TS更偏好使用短链酰基-CoA硫酯作为酰基供体,而不是带有芳香族氨基酸的苯甲酰-CoA。TS严格控制酰基供体识别的机制很有趣。我们假设苯甲酰-CoA相对于短链酰基-CoA硫酯的较大空间位阻,不适合TS的底物口袋,导致3β-苯甲氧基托品烷的低产量。通过分子对接,我们将TS的底物口袋与使用苯甲酰-CoA作为酰基供体的EcCS进行了比较。在TS中观察到一个距苯甲酰-CoA苯环2.5Å的Phe46残基(图6A),而在EcCS中的对应残基(Ile45)距苯环4.5Å(图6C)。当TS中的Phe46突变为异亮氨酸时,苯甲酰-CoA苯环与Ile46的距离为4.1Å(图6B)。当TS的第46个氨基酸为苯丙氨酸时,与巴豆酰-CoA和乙酰-CoA的距离分别为3.6Å和5.0Å(图6D, E)。然而,当TS的第46个氨基酸为异亮氨酸时,距离分别增加到4.9Å和6.3Å(图6F, G)。随后,我们进行了体外酶促实验以验证分子对接实验的结果。与我们的假设一致,F46I突变显著增强了TS在合成3β-苯甲氧基托品烷中的活性(图6H和表1),并显著降低了TS在合成惕佫酰假托品中的活性(表1)。TSF46I对苯甲酰-CoA的Kcat/Km值为99.32M−1.S−1,比TS的值高7.77倍(表1)。TSF46I对巴豆酰-CoA和乙酰-CoA的Kcat/Km值分别为3588.34M−1.S−1和16.17M−1.S−1,分别为TS的1.06%和3.5%(表1)。上述结果表明,Phe46是控制使用短链酰基-CoA硫酯作为酰基供体的关键氨基酸残基。

A. 含有苯甲酰-CoA的TS催化口袋。 B. 含有苯甲酰-CoA的TSF46I催化口袋。 C. 含有苯甲酰-CoA的EcCS催化口袋。 D. 含有巴豆酰-CoA的TS催化口袋。 E. 含有乙酰-CoA的TS催化口袋。 F. 含有巴豆酰-CoA的TSF46I催化口袋。 G. 含有乙酰-CoA的TSF46I催化口袋。 H. 使用苯甲酰-CoA作为酰基供体的TS和TSF46I的酶促实验。
为了进一步提高TS的催化活性,采用了共识蛋白设计。基于对His162范围内5Å内残基进行的保守替换分析,我们观察到只有Ser40不符合TS的保守替换模型(补充图S35A)。虚拟突变预测显示,将Ser40替换为保守的苏氨酸可能会改变3β-托品醇入口通道周围的氢键网络(补充图S35B, C)。Thr40与相邻螺旋中的Asn135建立了新的氢键(补充图S35B, C)。同样,TSS40T合成3β-托品醇酯的Kcat值显著高于其他样本,而Km值没有显著差异(表1)。TSS40T对3β-托品醇和巴豆酰-CoA的Kcat/Km值分别为35153.35 M−1.S−1和572303.6 M−1.S−1,比野生型TS高3.00倍和1.69倍(表1和补充图S35D, E)。上述结果表明,与野生型TS相比,TSS40T突变体能够更高效地合成3β-托品醇酯,包括惕佫酰假托品、3β-乙酰氧基托品烷和3β-苯甲氧基托品烷。
惕佫酰假托品在本氏烟中的生物合成
为了进一步评估TS的功能及其在烟草中惕佫酰假托品工程中的应用,我们在本氏烟叶片中重建了3β-托品醇酯的生物合成途径。将六个上游生物合成基因(新大赤楠的鸟氨酸脱羧酶,EnODC;颠茄的腐胺N-甲基转移酶,AbPMT;颠茄的N-甲基腐胺氧化酶,AbMPO;颠茄的III型聚酮合酶,AbPYKS;颠茄的托品酮合酶,AbCYP82M3;和曼陀罗的托品酮还原酶II,DsTRII)与TS一起瞬时共表达在烟草叶片中(图7A)。进行了表达黄色荧光蛋白(YFP)的对照实验。这些六种酶和TS表达的叶片生成了托品酮、杂化莨菪碱、3β-托品醇、惕佫酰假托品和3β-乙酰氧基托品烷(图7B–G)。对照叶片未产生任何惕佫酰假托品或中间产物。为了进一步促进从烟草叶片中收获惕佫酰假托品,我们用TSS40T替代了野生型TS。与野生型TS相比,TSS40T分别增加了惕佫酰假托品和3β-乙酰氧基托品烷的水平1.33倍和5.02倍。在烟草叶片中检测到的最高含量分别为6.14 μg.g−1 DW和0.13 μg.g−1 DW(图7F, G)。

A. 在本氏烟中重建惕佫酰假托品的生物合成途径。 B. LC-MS检测烟草叶片中的目标代谢物。 C. 托品酮的含量。 D. 杂化莨菪碱的含量。 E. 3β-托品醇的含量。 F. 惕佫酰假托品的含量。P = 0.003。 G. 3β-乙酰氧基托品烷的含量。P = 0.0068。对照组为表达YFP的烟草叶片。TS(TSS40T)为共表达六个托品烷生物合成基因和TS(TSS40T)的烟草叶片。数据以均值±标准差表示。每条线的三株独立植物的叶片用于代谢物分析。统计分析根据双侧独立样本t检验进行。
尽管烟草叶片共表达了参与惕佫酰假托品生物合成的七个基因,但产量仍然较低。这种低产量可能是由于植物中巴豆酰-CoA供应不足所致。增加巴豆酰-CoA的合成可能会促进惕佫酰假托品的积累。巴豆酰-CoA是植物中由异亮氨酸降解形成的代谢物59,60,但与巴豆酰-CoA形成相关的代谢基因尚未被鉴定。
我们假设CoA连接酶催化巴豆酰和CoA生成巴豆酰-CoA。在细菌中,绿杆假单胞菌含有一种异丁酰CoA合成酶(PcICS),该酶催化异丁酸和CoA之间的反应生成异丁酰CoA61。我们假设PcICS使用巴豆酰作为底物,因为异丁酸和巴豆酰的结构相似。与这一假设一致,纯化的PcICS催化CoA和巴豆酰生成巴豆酰-CoA(图8A, B)。不幸的是,当共表达六个上游生物合成基因、TSS40T和PcICS并将巴豆酰添加到烟草叶片时,惕佫酰假托品的水平并未显著增加(补充图S36),可能是由于3β-托品醇供应不足所致。

A. PcICS酶促实验。 B. 巴豆酰-CoA的质谱(MS)数据。 C. 惕佫酰假托品的半生物合成路线。 D. 惕佫酰假托品的生产。左Y轴,惕佫酰假托品的含量。右Y轴,3β-托品醇转化为惕佫酰假托品的转化率。HS-CoA表示辅酶A(CoA)。每种工程化大肠杆菌的三株独立转化体用于惕佫酰假托品的半生物合成。数据以均值±标准差表示。
然后,我们进行了进一步的实验。当在3β-托品醇供给下在烟草叶片中表达TS时,惕佫酰假托品的产量远高于惕佫酰假托品的从头生物合成(补充图S37)。当在供给足够的3β-托品醇和巴豆酰的烟草叶片中表达TS时,惕佫酰假托品的产量略有增加(补充图S37)。当在供给3β-托品醇和巴豆酰的烟草叶片中共表达PcICS和TSS40T时,惕佫酰假托品的产量显著增加,达到293.86µg.g−1 DW(补充图S37)。这些结果表明,通过供给足够的底物促进了烟草叶片中惕佫酰假托品的生产。
惕佫酰假托品在大肠杆菌中的生物合成
由于植物中惕佫酰假托品的产量较低,我们构建了一个高效的平台来生产这种用于治疗神经退行性疾病的潜在药物。因此,我们构建了一个工程化大肠杆菌生物反应器,通过供给现成的底物巴豆酰和3β-托品醇来生产惕佫酰假托品(图8C)。
为了生产惕佫酰假托品,在表达TS的大肠杆菌细胞中,在含有250 mg.L−1 3β-托品醇的LB培养基中进行发酵。尽管TS和TSS40T在植物和体外催化3β-托品醇和巴豆酰-CoA生成惕佫酰假托品,但我们在这个发酵系统中未检测到惕佫酰假托品(图8D)。这种差异可能归因于大肠杆菌中缺乏巴豆酰-CoA。因此,为了生产惕佫酰假托品,需要一种能够催化CoA和巴豆酰合成巴豆酰-CoA的CoA连接酶。因此,使用PcICS对大肠杆菌进行工程化以与TS或其突变体一起生产惕佫酰假托品。最高产量(357.40 mg.L−1,转化率为90.9%)在发酵60小时后检测到,此时共表达TSS40T和PcICS(图8D)。
讨论
打碗花精是一类潜在的抗营养因子,在多种茄科食品中被发现,如土豆和西红柿23。最近,涉及打碗花精生物合成的两个P450酶的发现,揭示了打碗花精生物合成途径的完整解释26。然而,导致惕佫酰假托品形成的关键酯化步骤仍未明确。在本研究中,我们成功鉴定了一种BAHD酰基转移酶,即惕佫酰假托品合酶(TS),其催化惕佫酰假托品的形成。考虑到打碗花精的糖苷酶抑制活性及其在代谢疾病治疗中的潜在应用,我们的发现为培育无打碗花精的茄科作物以及在植物或微生物中合成有价值的打碗花精提供了目标。
适当的亚细胞定位对酶活性至关重要62,63,64。尽管BAHD-ATs通常定位于细胞质29,但TS定位于线粒体(图4)。特别是TS N端的32个氨基酸在TS活性中起重要作用,但对于TS的线粒体定位来说是充分但非必要的。TS的线粒体定位虽意外但合理。众所周知,作为异亮氨酸降解产物的巴豆酰-CoA在线粒体中生成59,60,65,66。TS的线粒体定位表明该酶可能以快速且经济的方式利用酰基供体。早期研究表明,线粒体是生产需要乙酰-CoA作为底物的萜类的良好细胞器67,68。因此,通过TS的这种线粒体定位特性,可以实现更高效的BAHD-AT介导的酯生物合成的代谢工程策略。
基于三元复合模型(图5A),我们发现TS的酰基受体结合口袋由Ile35、Gln39、His162、Tyr280、Asn298、Leu300和Trp340组成,使3β-托品醇在适当的催化构象中稳定(图5B)。结合TS突变体的酶活性实验,我们提出了TS的催化机制。BAHD-ATs保守的HXXXD结构域中的His162是一个通用碱,它去质子化3β-托品醇的3-羟基,使其能够对酰基供体的羰基碳进行亲核攻击(图5E)。在此过程中,CoA从四面体中间体中作为离去基团释放,生成3β-托品醇酯(图5E)。
与报道的BAHD-ATs相似,TS表现出酰基供体的多样性。TS主要催化惕佫酰假托品的形成,因为它对巴豆酰-CoA的亲和力远高于对乙酰-CoA和苯甲酰-CoA(表1)。这可能是由于苯甲酰-CoA的芳香环比短链酰基-CoA硫酯具有更大的空间位阻69。对于TS,Phe46通过空间位阻严格控制酰基供体的识别。当使用苯甲酰-CoA作为底物时,Phe46的芳香环阻碍了苯甲酰-CoA的进入。将Phe46突变为异亮氨酸显著减少了空间位阻,显著提高了苯甲酰-CoA的利用效率,同时大大降低了巴豆酰-CoA的利用效率(图6,补充图S35和表1)。我们对TS催化机制的发现提供了关于BAHD-ATs介导的酯化的见解,可用于设计多样化的酯。
高效酶是通过合成生物学生产天然产物的关键。设计了一个更高活性的TS突变体TSS40T(补充图S35)。TSS40T的Thr40与Asn135形成了一个新颖的氢键,这在TS中不存在。这些分子内相互作用通过刚性化活性中心在酶稳定性中起重要作用70。因此,Thr40和Asn135之间的氢键可能有助于3β-托品醇结合口袋的稳定性,从而提高酶的催化活性。
与体外酶活性实验一致,TSS40T在工程化烟草中也表现出比TS更高的酯化活性,工程化烟草中构建了3β-托品醇酯的从头生物合成途径(图7)。虽然我们的结果证明了从烟草中合成惕佫酰假托品的可行性,但惕佫酰假托品的低水平阻止了烟草成为惕佫酰假托品生产的理想平台。大多数代谢通量指向杂化莨菪碱而非惕佫酰假托品,中间体供应不足可能限制了惕佫酰假托品的生产(图7)。为了更高效地生产惕佫酰假托品,我们设计了工程化大肠杆菌菌株进行惕佫酰假托品的半生物合成。为了克服大肠杆菌中缺乏巴豆酰-CoA的限制并提高效率,我们采用了绿杆假单胞菌的PcICS61。优势显而易见。一方面,利用PcICS的底物多样性,从CoA和巴豆酰合成巴豆酰-CoA,增加了巴豆酰-CoA的供应(图8C)。另一方面,TS的产物CoA可以作为PcICS的底物(图8C)。在这种CoA循环系统中,昂贵的底物CoA源自大肠杆菌自身,并被持续再生和利用(图8C)。结合高效TS突变体TSS40T的利用,我们构建了一个仅使用两种现成底物(3β-托品醇和巴豆酰)的惕佫酰假托品半生物合成生产系统,实现了357.40 mg.L−1的惕佫酰假托品生产,转化率为90.9%(图8D)。
总结,在本研究中,我们通过鉴定定位于线粒体的BAHD酰基转移酶惕佫酰假托品合酶(TS)揭示了惕佫酰假托品形成中缺失的一步。我们揭示了TS的催化机制并成功提高了其活性。最终,我们在烟草和大肠杆菌中重建了惕佫酰假托品的生物合成。我们的研究有助于表征托品烷生物碱的酶学和化学多样性,并提供了一种通过合成生物学生产惕佫酰假托品的方法。
方法
化学合成
由于市面上没有惕佫酰假托品、3β-乙酰氧基托品烷、3β-苯甲氧基托品烷和巴豆酰-CoA的标准品,这三种化合物在我们实验室进行了化学合成。使用的方法在之前的文献中有描述。将酸转化为酰氯衍生物,然后与3β-托品醇反应制备3β-托品醇酯71,72。巴豆酰-CoA硫酯通过PyBOP催化从CoA和游离酸合成73。
生成虎掌酰氯/乙酰氯/苯甲酰氯:逐次加入巴豆酰/乙酸/苯甲酸(10mmol,1当量)到草酰氯(20mmol,1.50当量),并加入一滴N,N-二甲基甲酰胺。2.5小时后,在减压条件下除去过量的草酰氯,得到淡黄色液体的虎掌酰氯/乙酰氯。
惕佫酰假托品/3β-乙酰氧基托品烷/3β-苯甲氧基托品烷:在带有分子筛(0.4nm)的圆底烧瓶中,将3β-托品醇(0.79mmol,1当量)溶解在5ml无水四氢呋喃中,然后加入4-二甲氨基吡啶(DMAP;0.04mmol)和三乙胺(1.58mmol,2当量)。通过连续氮气流建立和维持惰性气氛。将混合物在冰浴中冷却,然后逐滴加入虎掌酰氯/乙酰氯(2.37mmol,3当量)。在25°C下搅拌2小时,加入饱和Na2CO3溶液(10ml)和蒸馏水(20ml)稀释,然后用氯仿提取(3×30ml)。有机部分用无水硫酸钠干燥,在35°C下减压浓缩。然后,通过薄层色谱和高效液相色谱(HPLC)进一步纯化产物。
巴豆酰-CoA:将巴豆酰(50μmol,5.0当量)、PyBOP(16μmol,1.6当量)和K2CO3(40μmol,4.0当量)溶解在3ml新蒸馏的四氢呋喃(THF)中,在氩气环境下进行反应。将CoA(10μmol,1.0当量)溶解在1ml H2O中(通过超声去除O2),然后逐滴加入到THF溶液中。25°C搅拌3小时后,将反应混合物直接通过C18半制备反相柱进行HPLC分析。
使用高分辨率串联质谱(MS/MS)分析碎片离子以初步确定其结构(补充图S1-S4)。进一步使用核磁共振(NMR),特别是1H-NMR、13C-NMR和2D-NMR,确认合成化合物的结构(补充图S5-S24和补充数据1)。
生物信息学分析
通过使用隐藏马尔可夫模型(HMM)搜索BAHD-ATs(PF02458)基因,利用hmmsearch程序对来自Medicinal Plant Genomics Resource(Medicinal Plant Genomics Resource)的颠茄转录组进行搜索,以确认候选基因属于BAHD-AT家族。批量CD-Search(Welcome to NCBI Batch CD-search)进一步确认候选基因属于BAHD-AT家族。使用TBtools-II v2.042构建代谢物和BAHD基因家族表达模式关联分析的热图75。使用MAFFT v7.475的E-INS-I方法进行氨基酸序列比对。使用IQ-TREE v2.1.276和最大似然模型(LG + F + G4)构建系统发育树。树根使用藻类酶进化枝(进化枝0),并根据先前的方法分类35。此外,使用iTOL v677注释和可视化该树。
TS的表达分析
从4个月大的颠茄植物中采集次生根、初生根、成熟种子、成熟果实、花、茎、绿色果实和叶,用于RNA提取和通过TIANGEN Biotech(北京,中国)提供的试剂盒进行cDNA合成。使用iQ5系统(Bio-Rad,美国)进行实时定量PCR分析TS的组织表达图谱,以PGK作为参考基因78。扩增通过使用SYBR Green PCR MasterMix(Novoprotein,上海,中国)进行。用于qPCR的引物列在补充表S1中。
MBP标记的TS纯化和酶促实验
为了获得足够的蛋白质进行生化表征,根据大肠杆菌的密码子使用偏好合成了TS的编码序列,形成了密码子优化版本TSopt。通过带有BamHI和PstI限制位点的一对引物扩增TSopt的编码序列,然后插入pMAL-c5x以生成原核表达质粒pMAL-c5x-TS。用于构建载体的引物列在补充表S1中。随后将pMAL-c5x-TS质粒导入大肠杆菌BL21(DE3)进行表达。蛋白质表达在16°C下用0.25mM IPTG诱导过夜。使用淀粉树脂(Smart-Lifesciences)纯化MBP标记的TS蛋白。脱盐后,立即对新鲜蛋白进行酶促实验。3β-托品醇与三种不同的酰基供体(巴豆酰-CoA、苯甲酰-CoA和乙酰-CoA)结合,形成三组酶促实验的底物。反应混合物为含有20μg纯化TS的50mM甘氨酸-NaOH缓冲液,1mM 3β-托品醇和巴豆酰-CoA(或苯甲酰-CoA或乙酰-CoA)。
为了探索TS的最佳反应条件,测试了从pH 6到10.6的pH范围和从20°C到50°C的温度范围,并使用巴豆酰-CoA和3β-托品醇。最终,在pH 8.6和30°C下测量酶动力学。使用不同浓度的3β-托品醇(0.01–2mM)和巴豆酰-CoA(或苯甲酰-CoA、乙酰-CoA)(0.01–1mM)进行酶动力学分析。
亚细胞定位分析
使用TargetP-2.0(TargetP 2.0 - DTU Health Tech - Bioinformatic Services)进行TS的亚细胞定位预测。使用带有BglII和HindIII限制位点的一对引物扩增TS的编码序列、其N端32个氨基酸和不含32个氨基酸的TS,然后将其插入植物表达载体pGD3G-YFP以生成pGD3G-TS-YFP、pGD3G-N32-YFP和pGD3G-TSDel-N32-YFP。用于构建载体的引物列在补充表S1中。随后将构建物转移到农杆菌GV3101中,并共转化到本氏烟叶片中。培养48小时后,叶片制备成原生质体并用MitoTracker Red(贝尔泰姆,上海,中国)处理。使用共聚焦显微镜(Olympus FV1200)分析TS的亚细胞定位。MitoTracker Red发出红色荧光。TS-YFP、N32-YFP和TSDel-N32-YFP发出黄色荧光。
为了进一步确定TS在颠茄中的定位,从2个月大的颠茄植物的根中提取线粒体。进行差速离心分离线粒体52后,进行以下步骤:I. 将10克新鲜根冷冻在液氮中并充分研磨成细粉。II. 加入线粒体提取缓冲液(70mM蔗糖,210mM甘露醇,10mM HEPES,1mM EDTA,1mM PMSF,pH 7.5),并用Dounce组织研磨器均质10次循环。III. 将上一步获得的均质液在600×g、4°C离心10分钟,收集上清液。IV. 在1200×g、4°C离心10分钟,收集上清液。V. 在11000×g、4°C离心20分钟。此阶段获得的上清液为细胞质,沉淀为线粒体。Western blot进一步确定线粒体和细胞质含量。使用抗VDAC1抗体(兔,1:2000稀释)进行Western blot作为线粒体标记52,53,54。抗VDAC1抗体从Orizymes购买,目录编号为PAB220312。山羊抗兔IgG抗体(山羊,1:5000稀释)作为二抗使用,从Biosharp购买,目录编号为BL003A。使用Tanon 5200全自动化学发光图像分析系统(TANON,上海,中国)检测化学发光信号。然后,使用超声处理细胞质和线粒体(20kHz,超声3秒,停止7秒,50次),并使用BCA法测量蛋白质浓度。然后,用线粒体/细胞质粗蛋白孵育底物进行酶促实验。反应混合物为50mM甘氨酸-NaOH缓冲液(pH 8.6),含50μg线粒体/细胞质粗蛋白,1mM 3β-托品醇和巴豆酰-CoA。
病毒诱导的基因沉默(VIGS)
使用XhoI和KpnI限制酶将538bp的TS片段插入烟草嵌条病毒(TRV)载体(pTRV2)中,以构建质粒pTRV2-TS。用于构建载体的引物列在补充表S1中。然后,将pTRV1和pTRV2-TS共导入农杆菌GV3101,并瞬时转化18天大的颠茄子叶。作为对照组,共导入pTRV1和pTRV2质粒。28天后,对根进行表达和代谢物分析。本研究中使用的VIGS操作过程在之前的文献中有报道80。
在颠茄毛状根中过表达TS
通过带有BamHI和SacI限制位点的一对引物扩增TS编码序列,然后将其插入pBI121以生成植物表达载体pBI121-TS。用于构建载体的引物列在补充表S1中。为了评估TS过表达对托品烷生物合成的影响,通过农杆菌介导的转化建立了颠茄的根培养物。颠茄的根培养物按照已有的方法建立4,30,81。根培养物在含有100ml Murashige和Skoog液体培养基的烧瓶中生长,在25°C下避光培养28天后收获。使用八条独立转化pBI121的根线作为对照根培养物(CK组)。随机选择四条显著增加目标基因表达的转化线(OE-1, OE-2, OE-3和OE-4)进行代谢物分析。
代谢物分析
包括颠茄的根和毛状根以及烟草叶片在内的冷冻干燥根培养物被研磨成细粉,用于生物碱提取。根据之前报道的方法82进行这些托品烷的提取和检测。简而言之,将250mg干燥根粉放入1ml提取缓冲液(含0.1%甲酸的20%甲醇)中,在200rpm和25°C下振荡提取2小时。每个提取物通过0.22μm尼龙66过滤器(津腾,天津,中国),随后将滤液稀释100倍进行后续分析。使用Orbitrap Exploris 120 LC-MS(赛默飞世尔科技,匹兹堡,PA,美国)分析提取物中的生物碱含量。使用电子喷雾电离(ESI)正离子模式和全MS模式进行测量。仪器参数设置如下:鞘气流速35,辅助气流速10,喷雾电压3.00kV,毛细管温度350°C,S-lens RF水平50,辅助气体加热温度350°C。在MS/MS分析中高能碰撞解离的水平为30。
使用Thermo Scientific(匹兹堡,PA,美国)获得的CORTECS UPLC HILIC柱(2.1mm × 100mm,1.6μm)分析3β-乙酰氧基托品烷、杂化莨菪碱、托品酮、3α-托品醇和3β-托品醇。系统流速为0.3ml/min,温度为35°C。每次进样的样品溶液为3μl;洗脱程序见补充表S2。
使用Thermo Scientific(匹兹堡,PA,美国)获得的Hypersil GOLD C18柱(2.1mm × 100mm,1.9μm)分析惕佫酰假托品、3β-苯甲氧基托品烷、托品酮、莨菪碱、东莨菪碱、托品醇苷、巴豆酰去伪莨菪碱、巴豆酰1-羟基去伪莨菪碱和打碗花精A3。每次进样的样品溶液为3μl。系统流速为0.3ml/min,温度为35°C。详细的洗脱程序见补充表S3。
分子模拟
使用AlphaFold2 v2.3.0构建TS的蛋白模型并预测底物口袋83。底物(3β-托品醇、乙酰-CoA、巴豆酰-CoA和苯甲酰-CoA)从PubChem数据库(https://pubchem.ncbi.nlm.nih.gov/)下载,并使用AutoDock Tools v1.5.684对接到TS的辅因子结合位点。然后,针对不同底物和TS进行独立对接运行,选择结合能较低且方向有利的复合结构。使用PLIP v2.3.085分析底物与TS之间的相互作用。使用PyMOL 2.1(PyMOL | pymol.org)查看分子相互作用并处理图像。
共识蛋白设计
共识蛋白设计基于这样的假设:在给定位置,与非保守氨基酸相比,相应的共识氨基酸对蛋白质稳定性的贡献大于平均水平86。通过NCBI服务器的BLASTP,从公共数据库中获得与TS氨基酸序列具有50%以上同一性的基因,以进行共识蛋白设计。对中心在TS中His162周围5Å范围内的残基进行保守替换分析。通过TBtools-II v2.042绘制的序列标识图显示保守替换75。
惕佫酰假托品在本氏烟中的生物合成
为了在本氏烟中从头合成3β-托品醇酯,将EnODC、AbPMT、AbMPO、AbPYKS、AbCYP82M3、DsTRII和AbTS(或其突变体)共表达在烟草叶片中。通过带有AgeI和XhoI限制位点的一对引物扩增这些基因的编码序列,然后插入pEAQ-HT以生成一系列瞬时表达载体。用于构建载体的引物列在补充表S1中。随后将这些瞬时表达载体转移到农杆菌GV3101中,并瞬时转化到烟草叶片中。培养5天后,收获叶片并进行代谢物分析。
在烟草的半合成实验中,根据烟草密码子优化的PcICS构建到pEAQ-HT中。然后,通过GV3101介导的过程在烟草叶片中表达AbTS(或其突变体)和PcICS。2天后,以1mM浓度喂养3β-托品醇和巴豆酰。培养3天后,收获叶片并进行代谢物分析。
惕佫酰假托品在大肠杆菌中的生物合成
将TS(或其突变体)和PcICS的编码序列插入pETDuet-1中,以生成一系列原核表达载体。用于构建载体的引物列在补充表S1中。然后,将这些原核表达载体转入大肠杆菌(BL21)中。将工程化的大肠杆菌培养在100ml LB液体培养基中,培养至OD600为0.6。接着,将IPTG添加到最终浓度为0.25mM,并将底物250mg.L−1(巴豆酰和3β-托品醇)添加到培养基中。每6小时添加1ml培养液进行产物含量分析。
统计和可重复性
使用GraphPad Prism 8软件进行常规统计分析和酶动力学分析。使用双尾Student’s t检验计算样本或基因型之间的显著差异。各种实验中使用的生物重复的详细信息在“方法”部分以及主要图形和补充图例中提供。在所有必要的地方,没有使用统计方法来预先确定样本量。没有从分析中排除任何数据。随机选择四条显著增加目标基因表达的转化线(OE-1, OE-2, OE-3和OE-4)进行代谢物分析。
研究报告摘要
关于研究设计的更多信息可在链接到本文的Nature Portfolio Reporting Summary中获得。
数据可用性
用于转录组分析的数据来自Medicinal Plant Genomics Resources(Medicinal Plant Genomics Resource)。用于构建系统发育树的序列(46个颠茄的BAHD酰基转移酶和196个功能鉴定的BAHD酰基转移酶)列在补充数据1中。BAHD酰基转移酶的系统发育分析显示在补充数据2中。本研究中合成的化学物质的1H-NMR、13C-NMR和2D-NMR数据列在补充数据3中。TS的编码序列已上传到NCBI,其登录号为OP677554。源数据随本文提供。



















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









