









Crystal structure of vinorine synthase, the first representative of the BAHD superfamily
Vinorine合成酶的晶体结构:BAHD超级家族的首个代表

摘要
Vinorine合成酶是一种酰基转移酶,在植物Rauvolfia中抗心律失常单萜吲哚生物碱ajmaline的生物合成中占据中心地位。Vinorine合成酶属于苯甲醇酰基转移酶、花青素-O-羟基肉桂酸酰基转移酶、苯甲酰基/肉桂酸酰基-氨基苯甲酸转移酶、去乙酰维多林酰基转移酶(BAHD)酶超级家族,其成员参与多种重要药物的生物合成,例如吗啡、紫杉醇以及抗癌药物长春新碱和长春碱的前体维多林。Vinorine合成酶的X射线结构解析分辨率为2.6 Å。尽管序列相似性较低,vinorine合成酶的双结构域结构显示出与几种CoA依赖性酰基转移酶(如二氢硫辛酰转乙酰酶、多酮相关蛋白A5和肉碱乙酰转移酶)的结构有惊人的相似性。所有BAHD家族的保守残基都位于结构域1中。HXXXD基序的His160在催化过程中充当一般碱的角色,位于反应通道的中心,位于两个结构域的交界处,并且从两侧都可接触到。该通道贯穿整个分子,使底物和辅基能够独立结合。Asp164远离催化位点,似乎更多地具有结构作用而非催化作用。令人惊讶的是,DFGWG基序,这对于催化反应至关重要且是BAHD家族特有的,位于远离活性位点的位置,并且似乎只起到结构作用。Vinorine合成酶是BAHD超级家族中第一个已解析的蛋白质结构。
酰基-CoA依赖的BAHD1超级家族是一个快速发展的酶家族,最近才被定义(1)。BAHD这一名称来源于从植物物种中分离出的该家族前四种酶的首字母。该家族的成员在多种次生代谢产物的生物合成中发挥重要作用。由于最近在拟南芥基因组中已发现大约70个与BAHD相关的基因(2),因此该家族可能在不久的将来会显著扩展,并且在大多数情况下,它们的生物化学功能仍需进一步探索。许多存在于药用植物和真菌中的BAHD成员在生物合成途径中扮演着非常特定的代谢角色。最著名的成员包括参与卡瑟兰杂环生物碱维多林(3)合成的酶,该化合物是抗癌药物长春新碱和长春碱的前体;罂粟生物碱吗啡(4);二萜生物碱紫杉醇(5、6、7);花青素(8、9、10);以及一些植物抗生素(11),还有与花香气相关的酶(12)。
该家族中的一个典型酶是Vinorine合成酶(VS;EC 2.3.1.160),它在植物属Rauvolfia中抗心律失常的单萜吲哚生物碱ajmalan型化合物的内源性形成中占据核心地位。Vinorine合成酶位于复杂的生物合成途径的中间,从色氨酸胺和单萜类化合物secologanin开始,最终形成具有九个手性碳原子的六元环系统ajmaline(图1)。Ajmaline是来自印度植物Rauvolfia serpentina的抗心律失常药物,约有3000年的药用历史。Vinorine合成酶催化的是酰基-CoA依赖的可逆生物合成反应,将生物碱16-epi-vellosimine转化为ajmalan型生物碱vinorine。后者属于含有五环系统的sarpagan类生物碱,这是ajmaline生物合成过程中的最终环合反应(图1)。Vinorine合成酶将两种不同类型的生物碱在生物合成上连接起来,并在Rauvolfia属植物的生物碱代谢中扮演着核心角色。
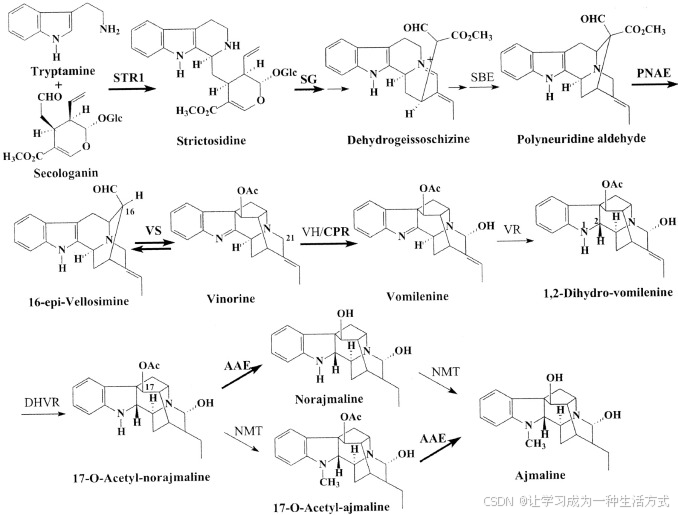
图1. 从严格可溶碱(strictosidine)到抗心律失常单萜吲哚生物碱ajmaline的生物合成途径,发生在药用植物R. serpentina的细胞悬浮培养物中。Sarpagan结构(16-epi-vellosimine)通过vinorine合成酶转化为ajmalan系统(vinorine)。该步骤依赖于酰基-CoA,并且在CoA存在下是可逆的。该图展示了合成酶在整个途径中的核心作用。已通过功能表达的酶催化的反应用粗体字表示。STR1,严格可溶碱合成酶;SG,严格可溶碱葡萄糖苷酶;SBE,Sarpagan桥接酶;PNAE,多神经甙醛酯酶;VH,呕吐宁羟化酶;CPR,细胞色素P450还原酶;VR,呕吐宁还原酶;DHVR,二氢呕吐宁还原酶;AAE,乙酰-ajmalan酯酶;NMT,N-甲基转移酶。
Vinorine合成酶(VS)已在R. serpentina的去分化细胞悬浮培养物中被鉴定并初步表征(13)。最近,它在大肠杆菌中被功能表达并纯化至均一性(14, 15)。该合成酶是一个单体酶,分子质量为46.8 kDa。通过了解该酶的初级结构,进行了序列比对研究,将VS归类为BAHD家族的新成员(15)。这一分类基于共识序列HXXXD和DFGWG。与其他BAHD家族成员的整体序列相似性较低(25-34%),可能表明该家族是从一个祖先基因分化演化而来(1)。通过在植物酶上进行定点突变实验,已证明这两个基序的某些功能重要性(9),同时也开展了针对vinorine合成酶的更详细的突变研究(15)。然而,结果表明,更好地理解催化过程及保守残基的功能,最好通过三维结构分析来实现。由于该酶家族的成员没有结构信息可用,我们已从R. serpentina中结晶了vinorine合成酶(16, 17),并在2.6 Å分辨率下解决了X射线晶体结构。结构分析结合先前报道的生物化学和突变研究,使我们能够提出VS催化的模型,并为BAHD超级家族中保守基序的功能提供了新的见解。
实验方法
VS的过表达、纯化和结晶
—VS被亚克隆到pQE-2载体中,并在大肠杆菌中过表达。可溶性蛋白通过镍-氮三乙酸亲和层析、阴离子交换和凝胶过滤层析纯化,如前所述。结晶时去除了N端的His标签(15, 16)。VS的晶体通过悬滴蒸发扩散法在32°C下获得。储液溶液包含0.1 m Tris-HCl,pH 8.7,2 m硫酸铵和2%的聚乙二醇400。酶(2–3 mg/ml)溶解在含有20 mM Tris-HCl,pH 7.5,10 mMβ-巯基乙醇,1 mM EDTA和0.5 mM乙酰-CoA的缓冲液中。SeMet VS通过抑制甲硫氨酸生物合成途径(18)获得,使用相同的表达载体和大肠杆菌菌株与原始VS的表达相同。SeMet VS的纯化和结晶按照类似于野生型VS的协议进行(17)。
数据采集和处理
—SeMet VS和野生型VS晶体通过向沉淀缓冲液中添加20–25%的甘油进行冷保护,然后在冷氮气流中闪冷至100 K。使用同步辐射在德国汉堡的德国电子同步辐射装置(DORIS存储环)的欧洲分子生物学实验室BW7A光束线采集了SeMet VS晶体在三种不同波长下围绕硒吸收边缘的多波长异常衍射数据。SeMet晶体衍射到3.24 Å,而野生型晶体衍射到2.60 Å。衍射数据使用HKL程序包(19)处理。晶体属于空间群P212121,且结晶非对称单元中有两个分子。数据采集和处理的统计数据汇总在表I中。
Table I. Crystallographic data and refinement statistics of VS
| Data set | Native | Peak | SeMet inflection | Remote |
|---|---|---|---|---|
| Data collection | ||||
| Wavelength (Å) | 0.9714 | 0.9787 | 0.9790 | 0.9537 |
| Unit cell (Å) | a = 82.72 | a = 82.25 | a = 82.27 | a = 82.29 |
| b = 90.46 | b = 90.28 | b = 90.30 | b = 90.34 | |
| c = 136.97 | c = 136.53 | c = 136.50 | c = 136.54 | |
| Total reflections | 108491 | 97554 | 99876 | 97730 |
| Unique reflections | 31740 | 15465a | 15607a | 15466a |
| Mosaicity | 0.24 | 0.58 | 0.51 | 0.57 |
| Resolution (Å) | 20-2.60 | 30-3.24 | 30-3.24 | 30-3.24 |
| Completeness (%) | 98.6 (99.1)b | 98.7 (97.1) | 99.6 (95.0) | 98.5 (90.9) |
| I/σ(I) | 29.5 (7.2) | 17.6 (3.4) | 18.0 (3.0) | 15.6 (3.0) |
| Rmerge (%)c | 2.8 (21.8) | 6.8 (42.6) | 7.0 (43.9) | 7.2 (39.2) |
| Refinement | ||||
| Resolution (Å) | 20-2.60 | |||
| Rcryst/Rfree (%)d | 21.1/27.2 | |||
| Average B (Å) for protein | 28.0 | |||
| Average B (Å) for water | 70.0 | |||
| No. of atoms | ||||
| Non-hydrogen | 6572 | |||
| Water | 145 | |||
| r.m.s.d. | ||||
| Bond (Å) | 0.013 | |||
| Angles (°) | 1.60 |
-
a
To compare with native data set, Bijvoet pairs are counted once.
-
b
The values in parentheses correspond to the last resolution shell.
-
c
Rmerge = ΣhklΣi|Ii (hkl) - 〈I(hkl)〉|/ΣhklΣ i〈I(hkl)〉, where 〈I(hkl)〉 is the average intensity over symmetry equivalent reflections.
-
d
Rcryst (Rfree) = Σhkl||Fo(hkl)| - |Fc(hkl)||/Σhkl |Fo(hkl)|, where Fo and Fc are observed and calculated structure factors, respectively.
结构解析、模型构建和优化
该结构通过使用欧洲分子生物学实验室汉堡自动化晶体结构解析平台的三波长多波长异常衍射方法解决(20)。在平台中,使用程序SHELXD(21)确定了异常散射原子的位点,并使用MLPHARE(22)对18个硒位点进行了进一步优化,以生成初始相位。通过密度修正进行相位改进,使用程序DM(23)。平台提供了正确的硒位点和可解释的电子密度图,部分α-螺旋模型包含了842个残基中的167个。部分模型由平台中的程序ESSENS(24)生成。
一旦电子密度图被认为是可解释的,就通过程序MAID(25)、RESOLVE(26)和XTALVIEW/XFIT(27)使用半自动过程构建了50%的多丙氨酸模型。之后,通过密度修正和2倍非晶体对称性平均,将相位扩展到2.6 Å,并使用XTALVIEW/XFIT继续手动模型构建,将相对完整的多丙氨酸模型构建到改进后的电子密度中。利用硒位点作为标记,在电子密度中放置正确的侧链,并成功地为单个分子构建了大部分侧链。然后,使用非晶体对称操作符生成了第二个分子。在此阶段,结构优化开始使用模拟退火法进行,随后在CNS(28)中进行位置优化和限制B因子优化。通过模拟退火和扭转角动力学协议以及手动构建,在CNS中迭代改进模型的几何结构和完整性。在最后阶段,使用非晶体对称性约束、大体溶剂修正和各向异性缩放进行优化,并在使用程序REFMAC5(29, 30)进行各向异性建模时,将每个单体的每个结构域定义为TLS组。优化过程中,使用3.2%唯一反射计算的自由R因子进行监控。共421个残基中,链A的8个残基和链B的9个残基未在电子密度图中显示(见“结果”部分),可能是无序的。由于乙酰-CoA未能在电子密度中找到,因此这些晶体被称为野生型晶体。野生型结构的优化统计数据如表I所示。
使用PRO-CHECK(31)评估了模型的整体几何质量。87.7%的VS氨基酸残基位于Ramachandran图的最有利区域,没有残基位于不允许的区域。所有图形均使用MOLSCRIPT(32)、PyMOL(33)和RASTER3D(34)生成。
CoA的建模
由于VS与CoA依赖性酰基转移酶家族的结构相似性,我们将CoA模型化到VS的活性位点。由于二氢硫辛酰转乙酰酶(蛋白质数据银行代码1EAD)的结构已解决并与CoA复合(35),因此该蛋白质被用作VS中CoA分子建模的基础。二氢硫辛酰转乙酰酶三聚体在溶剂通道中结合CoA,该通道位于两个亚基的界面。二氢硫辛酰转乙酰酶的两个亚基与VS结构中的单个单体相似,后者包含两个结构域。为了为VS·CoA复合物提供起始模型,两个结构进行了重叠。为了提高CoA定位的准确性,使用程序CONTACT(22)选择了与CoA分子距离在4 Å内的所有残基,并通过程序LSQKAB(22)以1.8 Å的r.m.s.d.将这些残基与VS中的相应残基进行重叠。此矩阵被应用于蛋白质数据银行代码1EAD中的CoA分子,以将CoA转移到VS结构的溶剂通道中。然后,对包含拟合CoA的单一VS分子模型进行了在CNS(28)中的模型优化,排除了X射线项。得到的模型随后用于进一步分析。
结果
VS的结构解析
VS的结构通过使用硒代美氨酸替代的VS的多波长异常衍射方法解决。晶体在空间群P212121中形成,非对称单元中包含两个分子。该模型经过优化,最终结晶学R值为21.1%(Rfree = 27.2%),使用的数据分辨率为20.0–2.6 Å。所呈现的VS原子模型显示了非对称单元中两个分子的所有残基,除了N端残基1–3和一个表面环(链A的235–239残基和链B的235–240残基)。结晶学信息总结于表I。晶体中非晶体对称性相关二聚体之间的接触通常较弱且为亲水性质。结构观察与生物化学数据一致,表明VS作为单体活性,之前通过尺寸排除色谱法已确认这一点(14)。
VS的整体结构
VS的结构包含14个β链(β1-β14)和13个螺旋(α1-α13),并由两个大致相等大小的结构域组成。两个结构域通过一个跨度约为36 Å的大交叉环(残基201–213)连接。结构域1包含一个混合的6条链β-折叠(β1-β2, β5-β7, β12),两侧被7个螺旋(α1–α7)覆盖(图2)。β链12(残基370–372)从结构域2突出,形成结构域1中的一个反向平行折叠。结构域1还包含一对β链(β3和β4),位于蛋白质表面,位于中心β-折叠的一端。结构域2包含6个螺旋和一个混合的6条链β-折叠(β8-β11, β13-β14)。结构域2中的一个环从β链9和β链10之间延伸到结构域1,并与α6接触。结构域1和结构域2共享非常相似的多肽骨架折叠,但它们的拓扑结构不同。它们的骨架可以在85个氨基酸位置上对齐,r.m.s.d为3.1 Å。两个结构域中的对应二级结构包括6条链的β-折叠和两个α-螺旋(结构域1中的α2和结构域2中的α9)。这些对齐位置之间的序列相似性较低,只有七对相同的残基(8.2%)
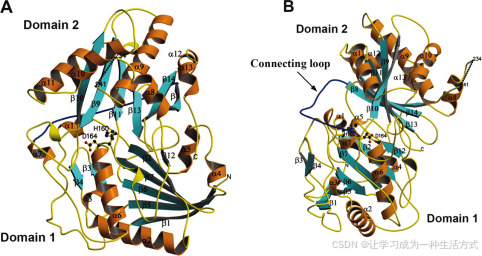
图2. Vinorine合成酶的结构 A和B分别表示VS结构的正交视图,如带状表示。N和C表示VS的两个末端。二级结构元素(α1–α13和β1–β14)被标注,结构域1和结构域2也有所指示。α-螺旋用橙色表示,β-链用青色表示。连接两个结构域的大交叉环(氨基酸201–213)用蓝色标出。虚线表示无序区域。保守的催化残基His160和Asp164以球棍模型显示。
溶剂通道的结构和活性位点的位置
一个溶剂通道贯穿整个VS分子(图3),由两个环组成,位于两个结构域之间,这两个环从结构域2突出,接触到结构域1(图2)。第一个环位于结构域2的两个平行链β11和β13之间,并包括结构域1的链β12。第二个环位于β9和β10之间。第一个和第二个环中的DFGWG和GN基序在BAHD超级家族中是完全保守的(图4)。VS结构中的活性位点HXXXD序列基序位于两个结构域之间的界面,且该基序的催化残基His160可以从通道的两侧接触到(图3)。
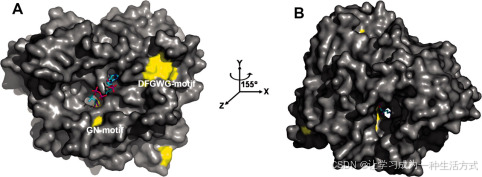
图3. Vinorine合成酶的表面表示,CoA建模于溶剂可接近通道中 A,VS的表面表示(正面);BAHD家族的保守残基以黄色高亮显示,CoA以球棍模型表示。DFGWG和GN基序在表面上标出。 B,按照A中描述的表面表示,但绕y轴旋转155°,显示通道的对面入口(背面)。B中清晰地可见贯穿整个酶的溶剂通道。
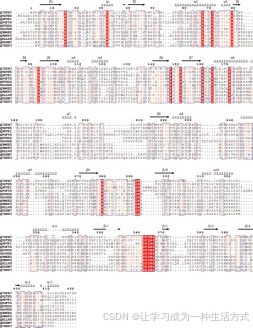
图4. 基于结构的BAHD家族蛋白质序列比对 R. serpentina的VS(Swiss-Prot登录号Q70PR7)与BAHD家族一些代表性成员的结构基础序列比对(草莓(Fragaria ananassa)的醇酰基转移酶、罂粟(Papaver somniferum)的salutaridinol 7-O-乙酰转移酶、长春花(Catharanthus roseus)的去乙酰维多林4-O-乙酰转移酶、天竺葵(Salvia splendens)的花青素5-O-葡萄糖苷-4′′′-O-马隆酰基转移酶、克拉克(Clarkia breweri)的苯甲醇乙酰转移酶、紫杉(Taxus cuspidata)的10-去乙酰巴卡亭III 10-O-乙酰转移酶、烟草(Nicotiana tabacum)的羟基肉桂酰转移酶、加拿大杉(T. canadensis)的3′-N-去苯甲酰-2′-去氧紫杉醇N-苯甲酰转移酶、燕麦(Avena sativa)的羟基蒽酸羟基肉桂酰转移酶、以及紫杉(T. cuspidata)的Taxadienol乙酰转移酶)。BAHD家族的代表性成员的Swiss-Prot登录号分别为Q9FVF1、Q94FT4、Q9ZTK5、Q6TXD2、O64988、Q9M6E2、Q8GSM7、Q8LL69、Q7XXP3和Q9M6F0。这些酶的序列相似性在25%–34%之间。序列上方的水平螺旋段表示α-螺旋(标记为α1–α13);水平箭头表示β-链(标记为β1–β14)。序列编号按VS显示。
VS结构代表CoA依赖性酰基转移酶家族的成员
可以通过使用整个VS分子或单独的结构域作为搜索模型,从二级结构匹配(www.ebi.ac.uk/msd-srv/ssm/cgi-bin/ssmserver)服务器中检索与VS结构相关的蛋白质。与VS最接近的结构是来自结核分枝杆菌的多酮合成酶相关蛋白5(Pap5;蛋白质数据银行代码1Q9J),它与VS的277个氨基酸对齐,r.m.s.d为2.58 Å,序列相似性为14%。可以与VS结构对齐的其他蛋白质包括嗜盐性维生素合成酶(VibH;蛋白质数据银行代码1L5A,r.m.s.d为3.5 Å,262个氨基酸,序列相似性为7%)、大鼠胆碱乙酰转移酶(蛋白质数据银行代码1Q6X,r.m.s.d为3.7 Å,244个氨基酸,序列相似性为10%)、人类肉碱乙酰转移酶(蛋白质数据银行代码1NM8,r.m.s.d为3.6 Å,236个氨基酸,序列相似性为10%)。VS的两个结构域也与氯霉素乙酰转移酶(蛋白质数据银行代码1CLA,r.m.s.d为2.75 Å,对齐VS结构域1,r.m.s.d为3.32 Å,对齐VS结构域2)和二氢硫辛酰转乙酰酶(蛋白质数据银行代码1EAD,r.m.s.d为3.75 Å,对齐VS结构域1,r.m.s.d为3.29 Å,对齐VS结构域2)显示结构相似性。所有这些对齐的蛋白质都是CoA依赖性酰基转移酶,且在活性位点中包含保守的HXXXD基序。在所有这些酰基转移酶中,除了VibH,HXXXD基序中的His残基在CoA依赖性酰基转移反应机制中起着至关重要的作用(35,36,37,38,39)。VibH也包含HXXXD基序,并且保守的His位于活性位点中,但将此His突变为Ala或Glu对催化几乎没有影响,表明VibH中的HXXXD基序在酰基转移催化中不发挥等效作用(40)。根据结构对齐和蛋白质中存在的序列基序,显然VS是CoA依赖性酰基转移酶家族的一个新成员。
讨论
VS的活性位点及拟议的反应机制
HXXXD基序在BAHD基因家族和其他一些酰基转移酶中高度保守。我们之前的生物化学和突变研究表明,VS中的His160对乙酰转移酶活性至关重要(15)。此处展示的VS结构解释了该残基的功能重要性。His160位于螺旋5和链7之间的一个环中,直接位于溶剂通道的中心。这种结构安排使得配体(乙酰-CoA)和底物16-epi-vellosimine可以独立地从酶的前面(CoA结合)和背面(底物结合)接近活性位点(图3)。事实上,根据之前从Rauvolfia细胞中富集的VS制备获得的动力学数据,提出了一种反应机制(13),其中涉及酶、底物和辅基的三元复合物。结论是底物和辅基在VS的活性位点独立结合,这与所描述的结构一致。有趣的是,催化位点的His160侧链采用了一种相当不寻常的构象(χ1 = –140°,χ2 = –31°),形成了一个内部氢键(2.9 Å),该氢键连接了咪唑氮Nδ1和相同氨基酸的羰基氧。此外,His160的Nδ1也与Ala163的羰基氧(2.9 Å)以及Asn293的侧链(3.0 Å)形成氢键。已解决的几种相关CoA依赖性酰基转移酶的结构与辅因子和底物复合物(如Azotobacter vinelandii二氢硫辛酰转乙酰酶(蛋白质数据银行代码1EAD和1EAB)与CoA和底物脂肪酰胺、以及小鼠肉碱乙酰转移酶(蛋白质数据银行代码1NDB和1NDI)与底物肉碱和CoA(35,36))已经建立。通过将二氢硫辛酰转乙酰酶单体与VS结构域1进行重叠,我们能够将CoA和脂肪酰胺的结合位点映射到VS上。在此模型中,His160位于与二氢硫辛酰转乙酰酶中的催化残基His610相同的位置。根据我们的VS-CoA模型,可以看到CoA从分子的前面进入溶剂通道(图3),位于β-链11和β-链13之间。脂肪酰胺的结合位点可以映射到CoA结合位点的对面(数据未显示)。基于生物化学结果(13,15)和我们的结构分析,我们提出了以下VS的乙酰转移机制。活性位点中的His残基作为一般碱,从17-去乙酰vinorine的17-羟基或CoA的巯基中提取质子,具体取决于可逆反应的方向。激活的羟基或巯基随后可以直接攻击乙酰-CoA或vinorine中的羰基碳,反应在不形成乙酰酶中间体的情况下进行(图5)。
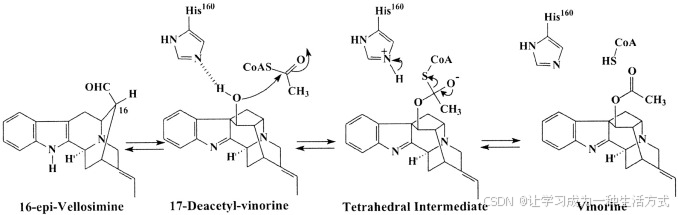
图5. Vinorine合成酶的拟议催化机制 提出了His160作为一般碱催化剂在底物去质子化中的作用。随后,建议形成一个假定的四面体中间体。
BAHD超级家族酶中的保守残基作用
多重蛋白质序列比对显示,BAHD家族蛋白质具有25%–34%的显著序列相似性(图4),并包含19个完全保守的氨基酸残基(图4)。有趣的是,所有这些保守残基完全属于结构域1(图6)。未来对该酶家族其他成员的结构分析可能表明,严格的结构域定位保守性是该超级家族的一个共同特征。

图6. BAHD家族中属于结构域1的残基的保守性 VS的带状图(颜色方案如图2所示),其中CoA(黑色)被建模到溶剂可接近的通道中,显示了与图3A中相似的方向。共有19个严格保守且完全属于结构域1的残基,这些残基在Cα位置用黑色球表示,并标有相应的残基名称。
在HXXXD基序中,His160与两个主链羰基氧通过氢键相连,此外还与Asn293的侧链形成氢键。Asp164远离His160和活性位点。尽管将Asp164突变为Ala导致完全失去活性(15),但Asp164的侧链取向并未参与与His160的氢键作用。因此,这两个残基不太可能像在人体肉碱乙酰转移酶中所提出的那样,作为催化二聚体发挥作用(37)。Asp164更可能参与与保守的Arg279形成盐桥,这对于维持活性位点的几何结构至关重要。因此,Asp164似乎在催化中不具有直接作用,更可能在结构上起作用,正如在其他一些酰基转移酶中所讨论的那样(38,39)。通过化学修饰和突变实验,已经证明了HXXXD共识序列中His和Asp在BAHD家族其他成员中的重要性(1,9,15)。因此,BAHD家族酶可能具有相似的催化His构象,并采用类似于VS提出的反应机制。
BAHD酰基转移酶中的另一个高度保守的区域是靠近C端的DFGWG基序。这个基序是BAHD酶家族特有的,已被建议对CoA的催化或结合至关重要(1,9,15)。然而,VS的结构分析揭示,DFGWG基序远离活性位点,因此不太可能在底物结合或催化中起直接作用。CoA建模到VS结合口袋中也显示,该特定的转折与CoA的泛酸部分或腺苷部分都没有接触(图6)。因此,这个保守的序列似乎通过维持酶结构的构象完整性而发挥重要的结构作用,而不是参与催化功能。Asp在DFGWG基序中的重要性已通过两项先前的突变实验确定。其突变为Ala导致了花青素5-O-葡萄糖苷-6′′′-O-马隆酰基转移酶的完全失活(9),并使VS的催化活性下降了65%(15)。DFGWG基序位于β11和β12之间的转折处(图6)。Asp362是该转折的一部分,氢键与Trp365和Gly366主链的酰胺氮形成氢键。因此,Asp362的取向在维持该转折结构中起着至关重要的作用。正如在肉碱乙酰转移酶和二氢硫辛酰转乙酰酶中观察到的那样,结构域2中的β11和β13在前面展开(图6),这可能为CoA的结合创造了开口。DFGWG基序也可能在维持CoA结合口袋的完整性方面起作用。CoA建模到结合通道中显示,几个残基可能与CoA接触。然而,除了His160之外,在该区域没有其他严格保守的残基。
为了更详细地了解结合口袋的性质和该酶的反应机制,仍需要获得配体和底物结合的VS晶体结构,目前这项工作正在进行中。
未来展望
如图1所示,ajmaline的生物合成是自然产物生物合成领域中最为复杂的途径之一。这也是现代蛋白质组学研究中最为著名的例子之一,其中不仅为直接参与该途径的所有酶提供了实验性证据,还包括催化旁路的酶(41)。这些工作共同为我们提供了Rauvolfia属植物中生物碱代谢的全面酶学知识。此外,约一半参与ajmaline生物合成的蛋白质已在大肠杆菌中实现了功能过表达。除了VS之外,另外两种酶(严格可溶碱合成酶和严格可溶碱葡萄糖苷酶)也已成功结晶,并且近期已进行初步的X射线分析(42,43)。因此,ajmaline的生物合成提供了一个独特的机会,使我们能够在不久的将来以结构水平深入研究生物碱的形成。这种研究将有助于更好地理解参与酶的极高底物特异性,还将帮助寻找一个特定的吲哚结合位点,并提供关于进化起源的信息,以及关于Rauvolfia酶单个结构域或整个酶及自然产物生物合成途径之间关系的更多信息。



















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









