






点击蓝字 关注我们
“肠-X”轴

iMeta主页:http://www.imeta.science
综 述
● 原文: iMeta (IF 23.8)
● 原文链接DOI:
https://onlinelibrary.wiley.com/doi/10.1002/imt2.270
● 2025年2月26日,南方医科大学陈鹏、王高峰、黄莉萍,南京医科大学/南方科技大学刘星吟,北京大学心血管研究所郑乐民,上海交通大学医学院刘宁宁,上海交通大学张晨虹,上海市东方医院南院吴文娟,中国医学科学院药物研究所王琰等团队在iMeta在线联合发表了题为“Gut-X Axis”的文章。
● 本文应用“肠-X轴”的概念来分别描述肠道和肝脏、心脏、大脑、肺、肾脏、骨骼、皮肤、生殖和内分泌系统之间的联系并介绍了相关的调节机制和临床干预策略方面的最新研究进展。
● 第一作者:林旭、于祖祥、刘洋、李长洲、胡绘、胡家淳、刘勉
● 通讯作者:陈鹏(perchen@smu.edu.cn)、郑乐民(zhengl@bjmu.edu.cn)、刘星吟(xingyinliu@njmu.edu.cn)、张晨虹(zhangchenhong@sjtu.edu.cn)、刘宁宁(fenghu704@163.com)、吴文娟(wwj1210@126.com)、黄莉萍(lphuang2006@126.com)、王琰(wangyan@imm.ac.cn)、王高峰(wanggaofeng@smu.edu.cn)
● 合作作者:杨琴、顾鹏、李嘉欣、Kutty Selva Nandakumar、胡高飞、张琪、陈兴宇、马慧慧、黄文烨
● 主要单位:南方医科大学、北京大学、南京医科大学、同济大学附属东方医院南院、中国医学科学院药物研究所、瑞典卡罗林斯卡学院、上海交通大学医学院、上海交通大学、南方科技大学
亮 点
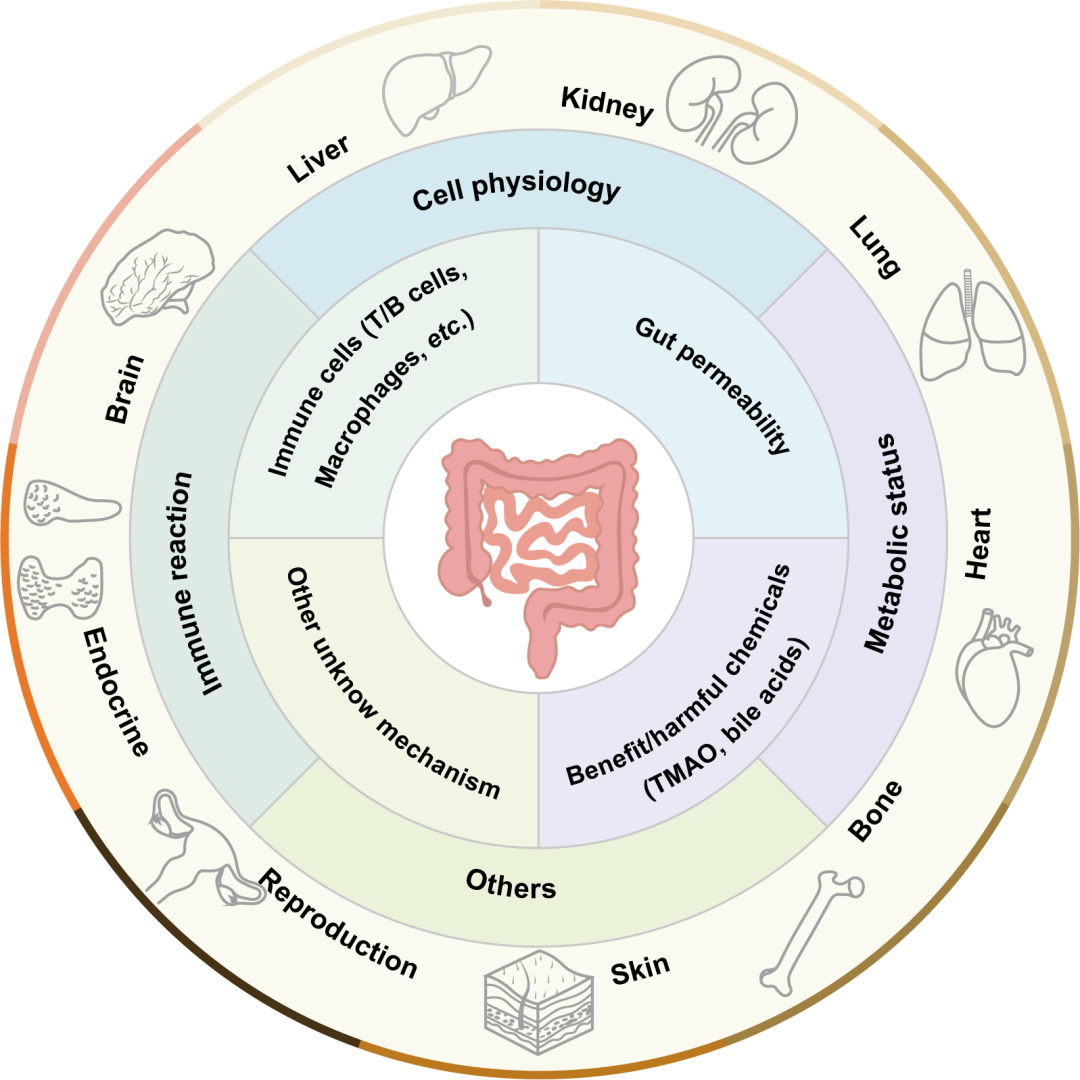
● 肠道与肠道微生物调节肠外器官的稳态;
● 肠道微生物产生的生物活性物质调节肠外器官疾病的发展;
● 肠道微生物靶向治疗是肠外器官疾病的一项有前景的治疗策略。
摘 要
近年来,关于肠道及肠道微生物在人体疾病中的调节功能的认识,不断取得进展,使我们更加关注肠道对于许多肠外器官(包括肝脏、心脏、大脑、肺、肾脏、骨骼、皮肤、生殖和内分泌器官)病理生理改变的作用。在本综述中,我们应用了“肠- X 轴”概念来描述肠道与其他器官之间的联系并讨论了与“肠- X 轴”相关的潜在调节机制和临床干预策略的最新研究发现。
视频解读
Bilibili:https://www.bilibili.com/video/BV1Br9cYGEMH/
Youtube:https://youtu.be/nwQIKqYLWEQ
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
肠道是推动多种肠外器官疾病发生、发展的关键器官。例如,肠道微生物产生的有毒物质三甲胺-N-氧化物(TMAO)是公认的代谢与心血管疾病的诱因。此外,肠道微生物生成的有益代谢产物,例如短链脂肪酸(SCFAs)中的乙酸、丙酸、丁酸,以及吲哚衍生物,能保护多个器官免受损伤。而某器官的病理变化也可能影响肠道微生物的组成与功能。因此,肠道与肠外器官之间的相互作用是复杂的。本综述将介绍当前关于“肠道-肠外某器官”轴(Gut-X axis)的研究现状并讨论潜在的临床干预策略。
肠-肝轴
概述
由于肠道和肝脏在生理和病理上有密切联系,“肠-肝轴”这一概念已被广泛接受。肠道与肝脏之间的连接包含多个重要组成部分。胆汁酸参与肠肝循环,而胆汁(包括初级胆汁酸和胆红素)可以被肠道微生物代谢。这些反应的产物能够调节肠道和肝脏的功能。此外,肠道微生物产生的代谢物,包括生物活性代谢物、微生物相关分子模式(MAMPs)、外膜囊泡和细菌残骸,可进入门静脉并在疾病发展过程中影响肝脏的病理生理。此外,来自肠道的免疫细胞可转移至肝脏,调节局部免疫微环境。最后,肠道和肝脏均能影响全身免疫和代谢稳态并在疾病进展中间接影响彼此(图1)。本节将讨论在特定肝脏疾病背景下,肠-肝轴的最新研究成果以及未来的展望。
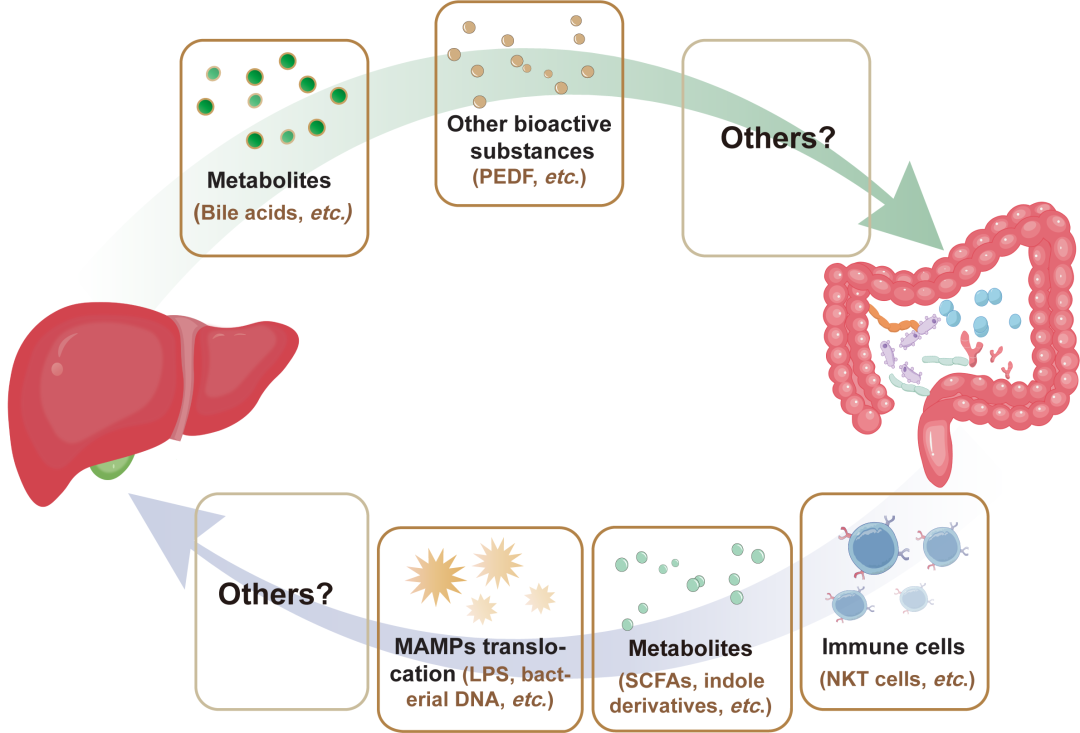
图1. 肝脏与肠道之间的双向联系
肠道产生的物质,包括微生物相关分子模式(MAMPs)与微生物代谢产物,能够直接影响肝脏;而肝脏产生的物质,包括初级胆汁酸和蛋白质,能够影响肠道的稳态。
脂肪肝
脂肪肝包括酒精性肝病(ALD)和非酒精性脂肪性肝病(NAFLD),是目前全球影响肝脏的主要疾病。已有充分的研究证明,肠道在酒精性肝病和非酒精性脂肪性肝病的进展中发挥着重要作用。
长期酗酒会导致肠道微生物过度生长、肠道微生物失调和肠道屏障破坏,从而促进MAMPs通过门静脉转移至肝脏。某些细菌物种的过度生长,例如肠球菌,会激活肝脏巨噬细胞中的Toll样受体(TLR)-2并加剧ALD中的肝脏炎症和损伤。此外,肠道抗菌肽的缺乏,会增加肝脏炎症并促进 ALD进展。肝脏巨噬细胞在细菌清除中起着关键作用,而乙醇减少了巨噬细胞中免疫球蛋白超家族补体受体的表达,从而降低了它们清除细菌的能力,促进了肝脏损伤。免疫球蛋白A(IgA)和聚合免疫球蛋白受体的异常表达或分布促进了细菌的转位,加剧了ALD。
与肠道微生物失调相关的肠道炎症是导致肠道通透性升高的主要原因之一。ALD相关的菌群失调具有多种特征,例如,许多细菌来源的毒素的量更大。由Enterococcus faecalis产生的外毒素直接参与饮酒引起的慢性肝脏损伤。有趣的是,特异性靶向肠球菌的溶细胞素的噬菌体,能显著改善小鼠的ALD;而由Candida albicans产生的外毒素则损害肝细胞并加重酒精性肝炎。除了细菌和真菌外,肠道中的病毒也可能参与ALD的发生;然而,由于病毒在培养中的传代较为困难,需要在未来解决这一技术限制后,才能进一步开展相关研究。
肠道本身参与了ALD的进展。肠道免疫细胞,包括自然杀伤T细胞(NKT)和Th17细胞,在酒精摄入期间,迁移到肝脏并对肝细胞产生有害影响。肠道免疫细胞产生的IL-22对维持肠道稳态至关重要,而ALD的进展与IL-22的分泌减少相关。重要的是,恢复IL-22浓度能够改善小鼠的ALD。肠道芳香烃受体(AhR)能够识别包括吲哚衍生物在内的多种生物活性物质,被认为有助于预防ALD,而生成AhR激动剂的工程菌能够缓解ALD。因此,肠道及肠道微生物对于ALD的进展有重要影响。然而,涉及的机制有待进一步阐明。
非酒精性脂肪性肝病/非酒精性脂肪性肝炎(NAFLD/NASH,又称代谢相关性脂肪性肝病/代谢相关性脂肪性肝炎,MASLD/MASH)是一种慢性肝脏疾病,病理机制复杂,涉及肠道及肠道微生物。与ALD类似,NAFLD患者也出现了肠道微生物失调和肠道屏障破坏。目前研究认为,肠道屏障功能障碍可发生在NASH的早期阶段,而维持肠道屏障功能有助于改善NASH相关的细菌转位和肝脏损伤。NAFLD/NASH的发展和严重程度与肠道微生物的组成和功能变化密切相关。例如,与促进炎症的肠道微生物产物相关的基因,在NAFLD/NASH的儿童患者中,表达明显升高。NAFLD患者肠道菌微生物的代谢功能存在差异。此外,肠道微生物的宏基因组特征可用于NAFLD相关肝纤维化的无创性诊断。值得注意的是,NAFLD/NASH患者中具有异常丰度和功能的肠道细菌,在不同的研究队列报道中有所不同,这可能与种族差异和疾病严重程度有关。
许多菌群及其产物会影响NAFLD。例如,Odoribacteraceae可诱导肝脏中特定亚型巨噬细胞的增生并参与NASH的进展。经典益生菌,例如Akkermansia和Bifidobacterium能防止小鼠发生NAFLD。此外,肠道细菌产生的次级胆汁酸,例如氢脱氧胆酸(HDCA),通过增加Parabacteroides distasonis的丰度和脂质代谢而改善NAFLD。一种新型胆汁酸——3-琥珀酰化胆酸,最近被发现能增加Akkermansia muciniphila的丰度并改善NAFLD。此外,肠道微生物在肠道中能降解尼古丁并缓解尼古丁相关的NAFLD的进展。除了细菌,肠道本身也参与了NAFLD的发生。例如,肠道过氧化物酶体增殖物激活受体(PPAR)α影响膳食脂肪的吸收并促进NASH的进展。由于脂肪肝与心血管疾病密切相关,因此,肠道对NAFLD的调节可能也会影响代谢和心血管异常。
毒素诱导的急性肝损伤
肝脏是最重要的解毒器官,在日常生活中,它会暴露于许多内源性和外源性的毒素,这些毒素可能导致肝脏损伤。四氯化碳(CCl4)毒素是最广泛用于建立化学性肝毒性动物模型的毒素。CCl4破坏肠道稳态,而靶向肠道微生物可能是缓解CCl4引起的急性肝损伤的有效手段。这可能是由于CCl4通过迅速破坏肝脏功能,影响肝脏与肠道之间的相互作用。维持肠道的健康菌群的平衡可能有助于支持肝脏的抗炎和抗氧化反应,从而保护肝脏免受急性肝毒性的损害。
对乙酰氨基酚(APAP,扑热息痛)过量使用,已被公认为发达国家急性肝衰竭的主要原因。损伤相关分子模式(DAMPs)长期以来被认为是肝脏炎症及随后的肝细胞损伤的主要原因。无论是MAMP还是DAMP,相关的免疫反应都会引起肝脏炎症。然而,最近的研究发现,APAP介导了肠道屏障的迅速破坏。肠道微生物也参与了APAP引起的急性肝衰竭。例如,A. muciniphila维持 肠道细菌功能,减少肝脏谷胱甘肽的耗竭和肝脏炎症,从而保护肝脏免受APAP引起的损伤。此外,研究已证实,细菌产生的苯丙烯酸决定了小鼠对于APAP引起的急性肝损伤的易感性,可能是通过影响细胞色素P450 2E1(CYP2E1)的表达。我们最近发现,Bifidobacterium衍生的吲哚-3-羧酸(I3C)能直接靶向CYP2E1并抑制其活性,这也能保护肝脏免受APAP引起的急性肝损伤。因此,菌群产物可能对APAP引起的肝脏病理生理具有重要影响。然而,其他药物引起的肝损伤也可能受到肠道微生物的影响。例如,胆碱酯酶抑制剂他克林(Tacrine)的去葡萄糖酸作用受到肠道微生物的影响,肠道中较高水平的去葡萄糖酸作用导致Tacrine在体内的暴露增加,从而增加肠肝循环并加重大鼠的肝损伤。
黄曲霉毒素B1(AFB1)是一种真菌代谢物和食品污染物,具有高度的肝毒性。AFB1破坏肠道法尼醇X受体(FXR)信号通路;一些有益的干预措施,例如褪黑素治疗,能增加肠道FXR的表达并减轻AFB1暴露后的肠道和肝脏的异常。此外,益生菌能够改善AFB1在人体中的毒性作用,但其潜在机制仍需进一步研究。这些发现表明,肠道及肠道微生物在影响毒素引起的急性肝损伤中起着重要作用。
胆汁淤积性肝病
胆汁淤积可以由许多肝脏损伤引起。自身免疫反应是胆汁淤积性肝病(CLDs)的主要病因之一。原发性胆汁性胆管炎(PBC)和原发性硬化性胆管炎(PSC)是由自身免疫功能失调引起的胆汁淤积性肝病的主要类型。目前已知,PBC患者的整体肠道微生物组成异常,特别是某些类型的PBC患者的肠道中,Faecalibacterium的丰度显著减少。熊去氧胆酸作为一种功能性的次级胆汁酸,是治疗CLD最有效的药物,它部分恢复了PBC特征性的不良菌群状态。PSC患者也表现出肠道微生物失调,涉及微生物组成和代谢产物的变化。这些研究表明,肠道微生物在PBC和PSC的发病机制中发挥着关键作用。
粪菌移植实验显示,CLD患者的菌群失调可以独立引起肝损伤,因此,正常菌群的破坏可能推动CLD的进展。与PBC相关的菌群失调,促进了NOD样受体吡啶结构域3(NLRP3)介导的肝脏炎症,但其中涉及的机制非常复杂。此外,来自肠道的Lactobacillus gasseri可诱导肝脏和全身炎症,表现为γδ-TCR+细胞和IL-17产生的增加,从而引起严重的肝损伤和纤维化。此外,在小鼠中,肠道内的Enterococcus faecalis对肝脏有害,促进CLD进展,而Lachnospiraceae则能够减轻肝脏炎症和纤维化。这些研究结果展示了肠道微生物与CLD之间相互作用的复杂性。除了PBC和PSC,妊娠期肝内胆汁淤积症(ICP)是临床上常见的另一种CLD。肠道的Bacteroides fragilis通过FXR抑制信号通路,导致胆汁酸合成增加并在小鼠中引发ICP。因此,肠道微生物对于CLD的发展有重要影响,可能是由于细菌介导的胆汁酸代谢或胆汁酸对细菌的繁殖和功能产生影响。此外,肠道微生物还会影响系统性和肝脏免疫反应并决定免疫微环境的状态。
肝纤维化/肝硬化
肝纤维化是慢性或反复肝损伤的结果,最终可能导致肝衰竭或肝癌。理论上讲,导致肝损伤的急性损害也可能影响纤维化的形成。与其他类型的肝病一样,肝纤维化/肝硬化伴随肠道微生物失调。有趣的是,许多口腔细菌在肝硬化患者的更加远端的肠道中被发现,其潜在机制可能很复杂。一种可能性是肝硬化期间,肠道抗菌能力下降,影响口腔中的细菌清除。然而,这一发现的意义仍不明确。此外,肝硬化患者的真菌多样性也异常,Bacteroidetes与Ascomycota的比值可用来预测90天住院风险。
肠道微生物由共生菌组成,无菌小鼠中能观察到更严重的纤维化,这提示我们,共生菌能预防肝纤维化。这可能归因于菌群对宿主免疫系统的刺激。然而,肠道异常可能加重肝纤维化。在纤维化实验中,肠道FXR信号通路受损,进而加剧了肠-血管屏障功能障碍和细菌易位,促进了肝脏损伤。肝星状细胞(HSCs)在细胞外基质积累中发挥着关键作用,肠道微生物可能直接或间接调节HSC的活性。例如,菌群代谢产物10-羟基-顺-12-十八烯酸抑制HSC中的Smad3信号通路并抑制其纤维化活性。相反,3-吲哚丙酸通过ROS依赖的MAPK信号通路激活HSC。因此,肠道微生物对肝纤维化的影响是复杂的。
靶向肠道微生物群可能是治疗肝纤维化的有效方法。Lactobacillus rhamnosus GG(LGG)能够恢复肠道FXR信号通路并降低肝脏中的胆汁酸浓度,从而改善胆汁淤积性纤维化。此外,粪菌移植(FMT)是缓解肝纤维化的有前景的方法。例如,来自肝硬化患者的粪便可能加重神经炎症,而来自健康供体的粪便则改善患有肝性脑病患者的认知功能。然而,其长期安全性和有效性仍需进一步的仔细分析。
肝癌
肝癌最常见的类型是肝细胞癌(HCC)和肝内胆管癌(ICC)。肠道微生物对HCC和ICC的发生、发展及治疗均有影响。脂肪肝相关的HCC与肠道微生物失调密切相关,其特征是益生菌的减少。重要的是,恢复肠道微生物的平衡可以预防HCC,这表明菌群失调是HCC的诱因。菌群失调可能导致全身性炎症,而且HCC患者的血浆中IL-8、IL-13和CCL3的浓度较高。类似地,随着ICC病情的进展,患者肠道中某些菌群(例如Ruminococcaceae)的丰度增加,菌群代谢异常,细胞因子表达改变。因此,评估肠道微生物组成可能成为一种早期检测HCC的无创诊断方法。
肠道微生物也参与肝癌的发病机制。肠道微生物来源的次级胆汁酸影响肝脏CXCL16的表达和NKT细胞的积聚,从而抑制肝癌的进展。此外,革兰氏阳性细菌成分——脂磷壁酸,通过COX2/PGE2轴抑制抗肿瘤免疫并加速HCC的进展。另外,Lactobacillus reuteri来源的色氨酸代谢物通过AhR/SREBP2轴抑制肝脏肿瘤的发生。肠道细菌还可以影响骨髓来源抑制细胞(MDSCs)的丰度和肝癌的进展;例如,革兰氏阴性细菌可能导致MDSCs的积聚并促进肝癌的发生。然而,革兰氏阴性细菌A. muciniphila则能减少MDSCs的丰度。因此,并非所有革兰氏阴性细菌对于MDSCs的作用都相同。肠道的Enterococcus faecalis通过影响TLR4信号通路而加速HCC的进展。此外,菌群生成的乙酸引起高-O-甘氨酸化状态并加速HCC的发展。然而,另一项研究表明,乙酸通过与GPR43结合,抑制IL-6/JAK1/STAT3信号通路,从而防止HCC的进展。这些矛盾的研究表明,肠道微生物及其代谢产物对肝癌的贡献复杂。因此,需要进一步了解肝癌的病因机制。
通过靶向肠道微生物,可能获得治疗肝癌的新方法。例如,Prohep(一种益生菌混合物)有益于肠道微生物并减缓HCC的进展。Bifidobacterium longum加速术后患者肝功能的恢复,而A. muciniphila则参与HCC患者对抗PD-1免疫疗法的反应。因此,恢复这些细菌物种的丰度可能提高免疫疗法的疗效。实际上,丁酸盐与索拉非尼协同改善其效果。然而,不当调控肠道微生物,例如使用过量的益生元(可溶性纤维菊粉),可能导致HCC的易感性。因此,需要进一步研究候选治疗方法,以优化肝癌患者的治疗效果。
肠-肾轴
概述
肠道微生物与宿主之间的相互作用还可以影响其他器官的功能。“肠-肾轴”即是指肠道微生态系统与肾脏之间的双向调节机制。这种串扰沟通机制通过宿主和微生物及其代谢物产生。肠道微生物通过产生胆汁酸、SCFAs、神经递质、尿毒素(UTs)和炎症因子等物质,影响肾脏功能。与此同时,肾脏可以排泄肠道来源的毒素和合成激素以调节肠道稳态(图2)。肾病患者的肠道微生态系统会出现紊乱,UTs产生增多,进一步损伤肾功能;而受损肾脏排泄毒素的能力减弱,毒素堆积损伤肠屏障,引起炎症,最终形成肠、肾之间的恶性循环。
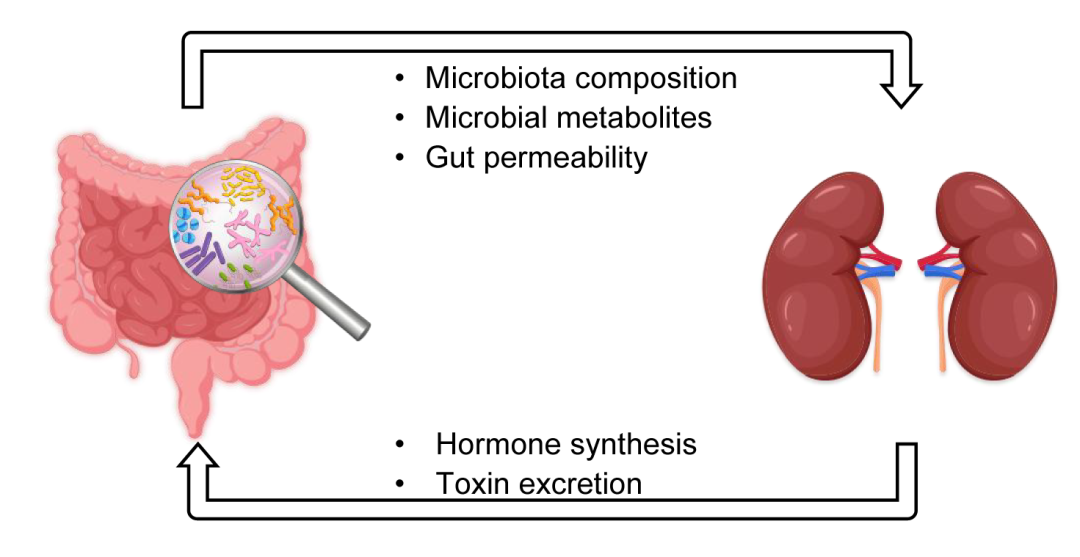
图2. 肠-肾轴互作机制
肠道稳态可以通过多种途径影响肾脏,包括菌群组成、肠道屏障通透性以及肠道微生物产生的功能性代谢物(如胆汁酸、短链脂肪酸、神经递质、尿毒素和炎症因子等)。与此同时,肾脏可以排泄肠道来源的毒素和合成激素以调节肠道稳态,从而形成双向互作。
肠道微生物与肾脏疾病
慢性肾脏病
慢性肾脏病(CKD)是全球主要的公共卫生问题之一,影响着全球10%−15%的人口。CKD患者的肠道菌特征主要表现为多样性降低,例如Bifidobacteriaceae、Lactobacillus、Blautia和Roseburia等SCFA产生菌减少,这表明高纤维饮食可能有助于CKD的治疗;蛋白质分解菌,例如Enterobacteriaceae、Enterococcaceae和Klebsiella增多,这些细菌能产生对甲酚硫酸盐(PCS)和吲哚硫酸盐(IS)等UTs,这些物质会损害肠屏障和肾脏功能。这表明高蛋白饮食可能会损害肾脏功能并增加终末期肾病(ESRD)的风险。此外,胆碱衍生的肠道微生物代谢物TMAO能够通过激活NLRP3炎症小体,加剧CKD患者的纤维化。此外,高水平的TMAO可使CKD患者的死亡率增加2.8倍并预示了治疗预后不良。
由于UTs无法通过透析清除,在体内的积累将加速终末期肾病(ESRD)的发生。与CKD患者不同,ESRD患者的血清次级胆汁酸水平显著升高,Eggerthella lenta和Fusobacterium nucleatum等次级胆汁酸合成细菌的丰度增加,同时,肠道中Faecalibacterium prausnitzii的丰度显著减少。这说明随着CKD的进展,肠道微生物的组成和功能也会发生变化,从而加速CKD向ESRD转化。总之,肠道微生态的变化增加了CKD的进展风险,是CKD患者需要重点监测的指标之一,也为基于肠道菌群的CKD发病机制和治疗策略做出重要提示,调节肠道菌群有望成为透析的补充疗法,值得深入研究。
糖尿病肾病
糖尿病肾病(DN)是糖尿病微血管病变最常见的并发症之一,也是导致ESRD的重要原因。肠道群失衡也被证实是DN的关键诱因之一,主要表现为Escherichia、Citrobacter和Klebsiella水平的增加,以及Roseburia的减少。不仅是肠道细菌,DN患者的肠道病毒组也发生了变化。病毒的丰度和多样性显著降低,尤其是裂解宿主细菌的噬菌体功能。这些研究表明,联合肠道病毒和细菌标志物或许是DN良好的诊断方法。肠道微生态组成的改变势必会带来功能代谢物的变化。研究发现,DN患者体内的乙酸盐可上调G蛋白偶联受体43(GPR43)的表达,增加肾小管上皮细胞内胆固醇的积累,破坏胆固醇稳态,导致蛋白尿和肾小管间质损伤。这也提示了SCFAs在不同疾病中的不同作用。与此同时,芳香族氨基酸除了促进CKD的发生外,也是DN的危险因素。例如,酪氨酸在肠道微生物产生的酪氨酸裂解酶等一系列酶的作用下转化为苯基硫酸盐(PS),促进蛋白尿的发生。外泌体作为细胞间通讯工具,在肠道环境紊乱中也可能成为致病因素。例如,DN大鼠中,肠道微生物来源的外膜外泌体增加,损害了肠道血管屏障,激活了半胱天冬酶-11/1通路,引发肾脏炎症。这些工作都为肠道菌紊乱与DN发病之间的因果关系提供了新见解,重塑肠道菌群有可能成为DN新的治疗方式。
免疫相关性肾病
由于肠道是机体重要的免疫器官,肠道微生物与自身免疫性疾病之间的关系也引起了广泛关注。狼疮性肾炎(LN)是系统性红斑狼疮(SLE)最常见的并发症,临床上尚无有效治疗方法。LN患者的肠道微生物失调,微生物的多样性和复杂性显著改变。其中,Ruminococcus gnavus(Rg)的增加最为明显。Rg特异性表达细胞膜脂聚糖,诱导宿主自发产生高水平的抗原特异性IgG抗体,这些抗体是SLE的致病因素之一。同时,低纤维饮食和其他破坏肠道稳态的饮食习惯,也通过增加肠道通透性和激活适应性免疫,加速LN的发病。
IgA肾病(IgAN)是最常见的原发性肾小球肾炎,属于自身免疫性疾病的一种。肠道稳态失衡可能在IgAN的发病中起到促进作用。在IgAN患者中,Escherichia、Veillonaceae、Akkermansia和Bacteroides的丰度显著上调,而Dialister的丰度显著下调。在机制上,上调的Akkermansia通过去糖基化IgA,暴露出新的位点供血清IgG结合,导致免疫复合物的形成和IgAN的发生。此外,肠道黏膜免疫可能也参与了IgAN的发病。例如,B细胞中γ-氨基丁酸(GABA)转运蛋白-2(GAT-2)表达水平较低,改变了肠道IgA+ B细胞的反应和肠道微生物组成,激活了GABA-哺乳动物雷帕霉素靶蛋白复合物1(mTORC1)通路,促进了生发中心B细胞的分化并诱导结肠SIgA的产生。总之,这些研究为深入理解肠道微生物与自身免疫性肾病之间的关系提供了新的视角,也为探索肠道与免疫疾病背后的机制提供了新的思路。
其他肾脏疾病
除了慢性疾病外,急性肾损伤(如肾结石和缺血再灌注)后肠道细菌组成的变化也有相关报道。有些研究还指出,肠道微生物会影响肾细胞癌患者对于免疫检查点抑制剂的反应。然而,大多数研究停留在相关性分析的层面,缺乏对于深层因果机制的阐述。即便如此,这仍为后续关于肠-肾轴的研究提供了新的方向。
肠道微生物与肾脏疾病的治疗
药物
肾脏、肠道和菌群之间相互作用研究有助于解释肾脏疾病背后的潜在机制,以肠道菌为靶点的药物也逐渐成为肾病治疗的潜在策略之一。中药(TCM),尤其是口服难吸收的天然药物,不可避免地会与菌群发生相互作用。因此,中药活性成分通过调控肠道菌来发挥肾保护作用已成为新的研究热点。小檗碱(BBR)是近年来我国研究最多的原创天然药物之一,其通过肠道微生物改善动脉粥样硬化和糖尿病等代谢性疾病已被多项研究证实。近期有研究首次发现BBR改善CKD的药效,阐明了其通过直接抑制梭状芽孢杆菌(对甲酚产生菌)丰度并同时抑制酪氨酸-对甲酚途径中关键酶TyrB的活性,降低肾毒素硫酸对甲酚水平,进而改善CKD的分子机制;同时,BBR还能增加产丁酸菌丰度,上调丁酸水平,协同发挥药效作用。这项研究被评为中国2023年度重要医学进展(药学领域),为中药单体通过肠-肾轴发挥药效的机理研究提供了新证据。黄酮也是肠道微生物领域的研究热点。异槲皮苷可通过调节肠道细菌菌的电子传递链,抑制氢质子势建立,从而抑制色氨酸传输,减少吲哚的生物合成。除了抑制肠道微生物来源的UTs外,丹酚酸B镁被发现能够改变肠道胆汁酸代谢谱,进而延缓DN的进程。不仅中药小分子能够与肠菌互作,大分子多糖与肠道菌也存在密切联系。牡丹多糖通过增加SCFAs和减少支链氨基酸(BCAAs),从而抑制炎症并改善肾脏损伤。除了中药,一些化学药物也被发现通过肠-肾轴为肾脏保护做出了贡献。小鼠口服广谱抗生素清除肠道菌后,TMAO水平显著下降,从而抑制了急性肾损伤向慢性肾病的转变,以此来看,靶向消除特定致病菌群或许可以成为临床中扼制肾病发展的重要思路。钠-葡萄糖共转运蛋白-2(SGLT2)抑制剂是新型降糖药物,最新研究也发现其也可以通过减少代谢苯丙氨酸和色氨酸的肠道微生物丰度来降低循环中的UTs水平。
然而,除了治疗作用外,一些药物还可能通过肠道微生物引发肾毒性。例如,长期服用中药泽泻会导致肠道微生物组成紊乱,引起肾脏氨基酸和磷脂代谢的失衡;中药苦参的活性成分苦参碱在口服后,会显著降低肠源性代谢物马尿酸的水平,进而引发肾脏能量代谢的失衡,造成毒性;化疗药物顺铂在使用时会加剧肠屏障的损坏,导致菌源的LPS更多地进入循环,形成微炎症环境,加重其肾损伤作用。这也提示在药物治疗肾病的临床实践中充分考虑肠道菌的作用,将有利于药物的合理使用。
益生菌
益生菌作为一种调节肠道微生物的新型干预手段,在CKD的治疗中受到了越来越多的关注。F. prausnitzii能够产生丁酸并作用于G蛋白偶联受体43(GPR43)发挥肾脏保护作用。预防性补充Lactobacillus casei Zhang可提高SCFAs和烟酰胺的水平并减少肾脏巨噬细胞和肾小管上皮细胞中的炎症反应。Bacteroides fragilis可以上调肾脏葡萄糖转运蛋白SGLT2的表达,增加5-脱水葡萄糖的重吸收,最终激活胆汁酸受体TGR5,抑制氧化应激。Lactobacillus paracasei HII01通过改善肾脏有机阴离子转运蛋白3(OAT3)的功能,促进肠毒素的排泄。益生菌不仅对CKD具有良好的治疗效果,对其他肾脏疾病也同样有效。乳杆菌属可降低肠道中的IL-6水平,同时增加IL-10,进而使得肾脏的Treg-Th17平衡偏向于Treg,从而改善LN。口服Oxalobacter formigenes可以预防大鼠肾脏中草酸钙晶体沉积,减少肾结石的形成。益生菌疗法更新了以往探讨疾病发生发展的分子机制的理念,为临床治疗拓展了视野。而菌群疗法的长期干预是进一步研究的方向。
饮食
除了药物干预外,肾病患者还能通过改变饮食习惯,延缓甚至改善肾病。膳食蛋白质中含硫氨基酸可以产生硫化物通过翻译后修饰来影响蛋白质功能。例如,对肠道菌中色氨酸酶(TnaA)的巯基化修饰抑制了其生成吲哚的能力,从而减少了UTs的产生。说明饮食能通过翻译后修饰的机制,改变肠菌功能,为研究饮食-菌群互作带来新启示。另外,模拟禁食饮食、限制80%或100%含硫氨基酸的饮食(SR80/100)和热量限制饮食等饮食习惯也可以通过调节半胱氨酸的氧化和硫化氢依赖性分解代谢发挥肾保护作用。这些结果是通过菌群干预影响宿主机体状态的又一例证,对于研究肠道菌群-宿主互作、开发缓解肾病的饮食干预手段等都具有参考价值。
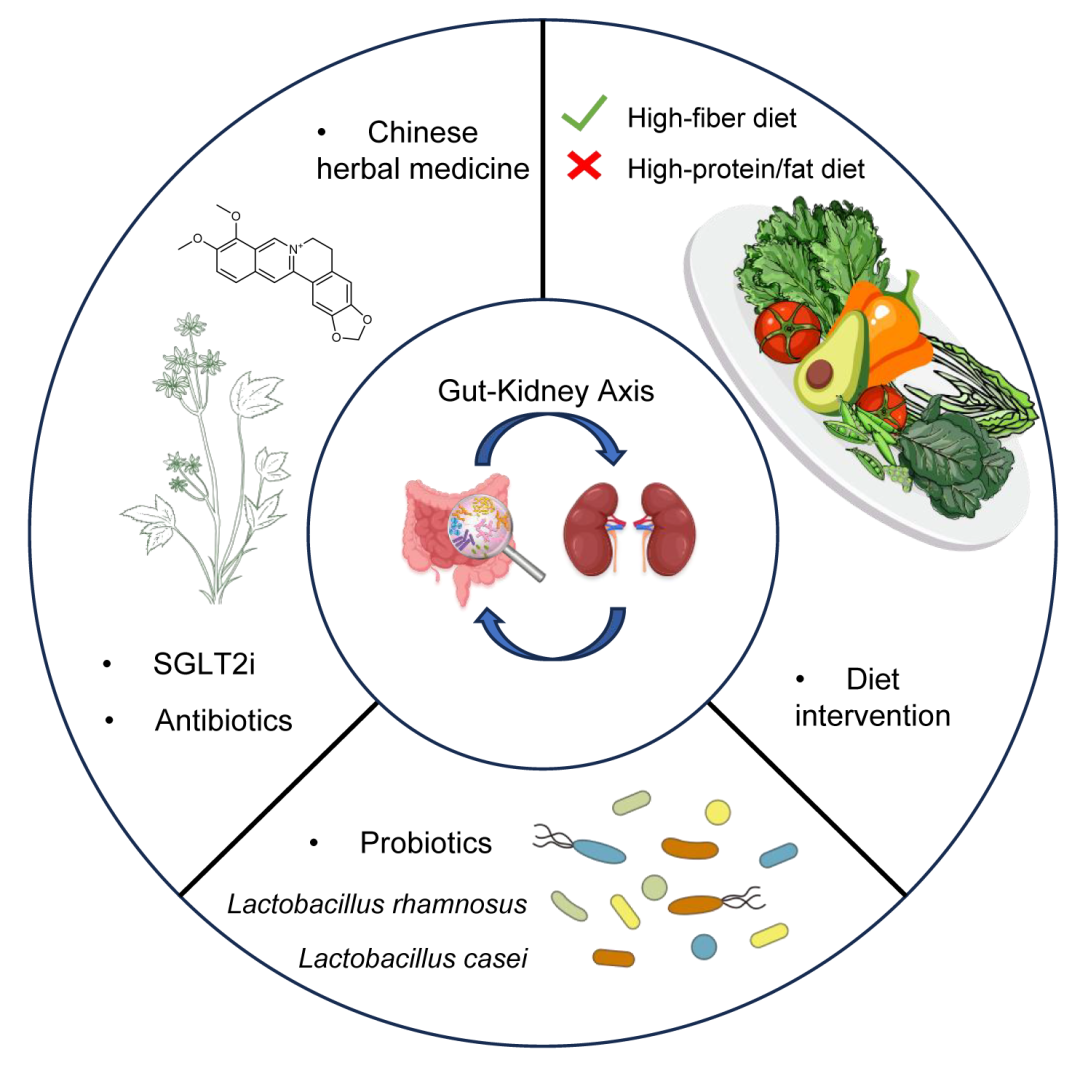
图3. 基于肠-肾轴的干预策略
通过肠道微生物干预肾脏疾病的主要方式有三种,包括饮食、益生菌和药物。高蛋白/高脂饮食通过破坏肠道微生物而加重肾脏疾病,高纤维饮食则可以恢复肠道微生物的稳态,有助于保护肾脏。益生菌,如乳杆菌属,通过产生有益代谢物来缓解肾脏疾病。药物治疗,以中草药和抗生素为代表,主要通过抑制尿毒素的生成以及抗炎和抗氧化机制发挥肾脏保护作用。
肠-肺轴
概述
肠-肺轴正受到越来越多的关注,因为肠道微生态的稳定(即肠道微生物的稳态)在维持肺部健康中起着至关重要的作用。肠道微生物失调可以通过肠-肺轴引发多种肺部疾病,包括病毒性肺炎、哮喘、结核病和慢性阻塞性肺疾病(COPD)。临床证据已确认肺部与大肠之间存在病理生理学的联系。肺部与肠道之间的结构同源性构成了“肠-肺轴”的基础,肺、气管、呼吸道上皮和肠道在胚胎发育过程中均来源于内胚层。肺的下叶与横结肠相邻并通过隔膜相连。在生理学上,肺与大肠也有功能上的联系。肺主要负责吸入氧气和呼出二氧化碳,而大肠则主要负责水分和电解质的吸收与排泄。因此,它们的功能互为补充,共同维持机体的正常生理功能。这包括典型的黏膜结构、分泌型IgA、细胞因子、固有淋巴细胞、B淋巴细胞及其他免疫细胞。胃肠道和呼吸道的黏膜是共同黏膜免疫系统的组成部分(图4)。淋巴细胞向黏膜的归巢是选择性的,是黏膜免疫的前提。当黏膜部位受到影响时,通过黏膜免疫通路激活局部免疫反应,进而引发广泛的免疫反应并在不同黏膜部位产生不同程度的反应性。例如,肠道微生物可以诱导产生2型和3型固有淋巴细胞(ILCs),这些细胞通过淋巴和循环系统迁移到呼吸道,增强呼吸系统的免疫活动。这种相互作用在疾病中也可能存在,例如,分段丝状细菌(segmented filamentous bacteria,SFB)特异性的Th17细胞,具有双重T细胞抗原受体,一个针对SFB,另一个针对自身抗原,可以迁移到呼吸道并导致肺损伤。反过来,呼吸道的微生物感染可以减少SFB的数量并增加肠道中大肠埃希菌的数量,从而刺激肠道上皮细胞中IL-15的表达,促进Th17细胞的极化,并可能导致肠道免疫损伤。此外,肠-肺轴的内分泌通路也至关重要。肠道和肺部的通讯主要通过可溶性微生物成分和代谢物的循环运输进行。肠道微生物成分,如病原相关分子模式(PAMPs),与固有免疫系统的模式识别受体(如Toll样受体TLRs)相互作用,触发炎症反应,从而激活免疫系统。肠道TLR信号的激活触发肺部免疫系统。此外,肠道黏膜的固有免疫细胞可以通过外周循环直接迁移到呼吸道,参与维持呼吸道免疫稳态。定植在黏膜表面的ILCs可以增强免疫反应,维持黏膜完整性并保护组织稳态。IL-25或由蠕虫引发的炎症ILC2s可以依赖鞘氨醇-1-磷酸(S1P)迁移到淋巴和血液循环,最终聚集在肺部以对抗蠕虫感染并促进组织修复。
肠道微生物不仅调节胃肠道的免疫反应,还影响远距离的器官(例如呼吸系统)的健康与疾病。通过调节肠道微生物,肺部和肠道在生理上相互联系并在病理上相互影响。本节将讨论肠道微生物与呼吸系统疾病的关系、肠-肺轴的作用机制以及临床应用。
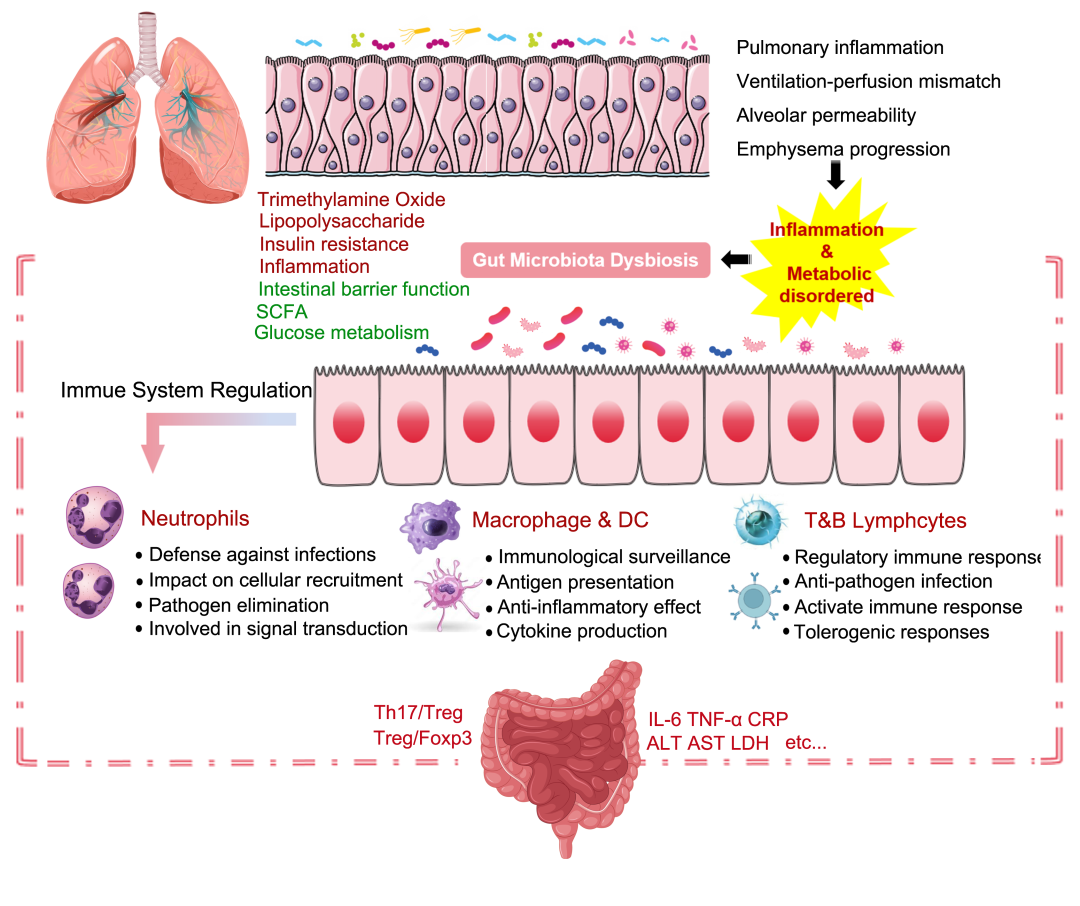
图4. 肠-肺轴通过免疫和内分泌通路介导
肠-肺轴通过免疫细胞迁移和肠道微环境变化而影响肺部炎症、通气-灌注失衡、肺泡通透性和肺气肿。ILC2和ILC3细胞可以损伤气道上皮细胞,增加黏液分泌并加剧炎症。相反,树突状细胞和单核细胞可以减少肺部炎症。肠道微环境的变化,释放细胞因子如IL-17和TNF-α,这些细胞因子通过血液循环到达肺部,降低感染风险。肠道微生物刺激B细胞产生IgA,增强肺部免疫功能并减缓菌群失调。这能够增强中性粒细胞反应,减少细胞因子产生,降低肥大细胞活性并促进淋巴细胞的耐受性。
肠-肺轴在呼吸系统疾病发病机制中的作用
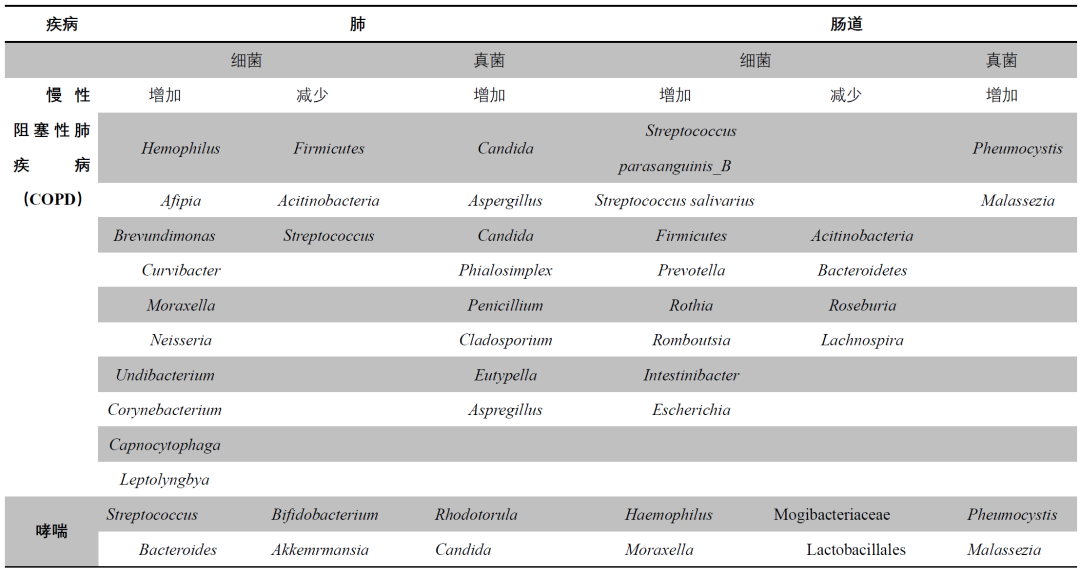
COPD患者的疾病进展与肠道微生物密切相关。COPD患者的肠道微生物多样性和组成与健康对照组相比,存在显著差异(表1),这种差异主要体现在门水平上,包括Firmicutes、Bacteroidetes、Proteobacteria、Actinobacteria和Verrucomicrobia等。肺部菌群由Prevotella、Veillonella、Pseudomonas、Acinetobacter、Fusobacterium、Sphingomonas、Rothia、Staphylococcus和Streptococcus等属构成,这些属构成了区分COPD患者与正常人群的“核心菌群”。此外,Achromobacter、Gemella和Capnocytophaga等属也已在COPD患者的下呼吸道中被发现。进一步的研究发现,COPD患者肺功能的下降与多个属之间存在相关性,包括Streptococcus、Streptomyces和Veillonella等。
肠道微生物失调、微生物转位和SCFAs减少与大鼠肺气肿的严重程度相关。高纤维饮食显著增加了Bacteroidetes的丰度,Bacteroidetes是SCFAs的主要生产者,SCFAs是肠道微生物的主要代谢产物。SCFAs在抗炎机制中发挥着重要作用,包括通过亚油酸途径介导的机制,这些机制在阻止COPD的进展、缓解炎症和肺泡损伤方面具有重要作用。近期一项研究使用非靶向粪便宏基因组学和代谢组学,报告了COPD患者与健康对照组之间肠道微生物和代谢组的组成存在显著差异,COPD患者中Streptococcus parasanguinis_B的丰度增加。在关于慢性阻塞性肺疾病急性加重(AECOPD)的早期研究中,发现粪便菌群中S. parasanguinis_B和Streptococcus salivarius的丰度增加,而在痰液中仅S. parasanguinis_B的丰度增加。此外,在一项比较肠道和肺部菌群的研究中,采集了15名COPD加重并接受14天抗生素和类固醇治疗的患者的样本,观察到两种样本类型之间某些门的丰度存在差异;然而,核心菌群的多样性没有差异。这些研究共同表明,肠-肺轴的潜在相互作用可能在COPD的发病机制中发挥着至关重要的作用。
哮喘
支气管哮喘是一种慢性气道炎症性疾病,主要特征是以气道高反应性作为主要的病理生理改变。多种免疫细胞,包括肥大细胞、嗜酸性粒细胞和T淋巴细胞,参与过敏性哮喘的发病机制。微生物群是免疫、代谢和细胞功能的关键调节因子,能够响应与哮喘相关的炎症信号并可能在哮喘的易感性、严重程度和表型中起到介导作用。
Strachan的“卫生假说”强调了肠道微生物在哮喘发病机制中的不可忽视的作用(表1),而Rook等人的“老朋友理论”则认为,恢复健康的微生物环境可以降低过敏性疾病的风险。肠道微生物多样性的减少与特应性疾病的风险增加相关,而发育延迟则可能触发遗传易感性,导致哮喘的发生,表现为微生物多样性的下降和机会性病原体的过度生长。Stiemsma等人观察到,已被诊断为哮喘的儿童,其肠道微生物失调在三个月时已显现。一月龄的婴儿在口咽部定植如Haemophilus influenzae、Streptococcus pneumoniae或Moraxella catarrhalis等细菌,与儿童哮喘的风险显著相关。一月龄婴儿定植的Clostridium difficile,在六岁之前与哮喘和喘息的发生风险存在关联。对于有哮喘风险的婴儿,Veillonella、Rothia、Bacteroides和Faecalibacterium等属在出生后头100天内通常会减少。上呼吸道早期定植Moraxella genus与呼吸道感染相关,在6至17岁患有哮喘的儿童中,鼻分泌物表现出显著的嗜酸性粒细胞活化,尤其是在Moraxella定植的情况下。哮喘患者可能由于类固醇的干扰而出现粘膜免疫的变化,导致有益细菌,如Bifidobacterium和Lactobacillus减少以及病原菌的过度生长。早期肠道群通过调节Th1/Th2平衡可能预防哮喘气道炎症;小鼠研究表明,Lactobacillus、Bifidobacterium和Bacteroides等共生菌可以通过诱导Treg细胞的产生、减少Th1促炎因子的释放、增强Th2和Th17细胞因子的产生,从而缓解包括气道在内的某些组织炎症反应。补充有益的肠道微生物也可以相应改善肺部疾病。此外,给孕鼠喂食高纤维或乙酸盐可以通过Foxp3/Treg通路保护其后代免受过敏性气道疾病的影响。
呼吸道感染
呼吸道感染一直对全球人类健康构成重大威胁并导致了巨大的医疗费用。无菌小鼠对流感病毒的反应较弱。SFB缺乏的小鼠与携带这种细菌的小鼠相比,表现出更高的细菌负荷、更严重的肺部炎症和更高的死亡率。这些研究结果共同表明,肠道微生物在抵抗呼吸道感染中发挥着重要作用。
抗生素的滥用导致肠道菌群失调,可能促进潜在病原体从肠道转移到口咽部,从而引发呼吸道感染。一项关于肠道微生物失调与严重急性呼吸综合症冠状病毒2型(SARS-CoV-2)感染关系的研究发现,COVID-19患儿的肠道微生物的均匀度显著高于对照组(表1),并且Bacteroidetes和Firmicutes的丰度增加,而Proteobacteria的丰度减少。与健康对照组相比,COVID-19患儿有相对丰度较高的条件致病菌和环境细菌且细菌多样性显著降低,益生共生菌的相对丰度较低。与整体多样性变化不同,H1N1流感病毒感染的患者表现出肠道细菌群落组成的改变,Bacteroidetes的丰度增加,同时Firmicutes的丰度相应减少。Grayson等人发现,摄入不吸收的链霉素显著降低了肠道微生物的多样性,但对肺部菌群没有显著影响;然而,肠道微生物多样性的减少与呼吸道病毒感染的死亡率显著增加相关。给大鼠口服硫酸甲硝唑后,导致厌氧菌失衡,随后感染流感病毒,提示肠道厌氧菌失衡加剧了流感病毒引起的病理损伤。此外,感染H7N9流感病毒的患者与健康个体之间的微生物群体差异显著,Eubacterium、Ruminococcus、Bifidobacterium和Lactobacillus的丰度显著减少。
此外,平衡的肠道微生物能够增强肺结核患者的抗感染能力并减少对Mycobacterium tuberculosis(MTB)的易感性。目前认为,这可能与患者食欲显著下降,导致总体膳食纤维摄入量减少,从而选择能够更有效利用宿主粘多糖的Bacteroides并减少肠道中Prevotella的数量有关。Prevotella的相对丰度与激活的CD4+和CD8+细胞数量呈正相关,表明肠道中丰富的Prevotella反映了机体免疫功能的水平。Prevotella可以利用丙酮酸作为底物,通过乙酰辅酶A途径合成SCFAs,这些短链脂肪酸在胃肠道中被处理并通过循环运输到肝脏代谢。最终的代谢产物部分通过肠-肺轴与外周循环,运输到肺部,影响肺组织中免疫细胞的分化和成熟,参与肺部炎症的调节。胡等人使用宏基因组测序,分析了肺结核患者肠道微生物的特征并鉴定了39条与生物合成相关的途径,确认了肠道微生物与代谢功能的显著变化。在肺结核患者中,产生短链脂肪酸的细菌,如Prevotella显著减少,并且与此相关的5条途径显著缺失。同时,这些细菌丰度的下降反映了全身炎症的加重和宿主免疫功能的损害。然而,这些机制仍需进一步深入研究。
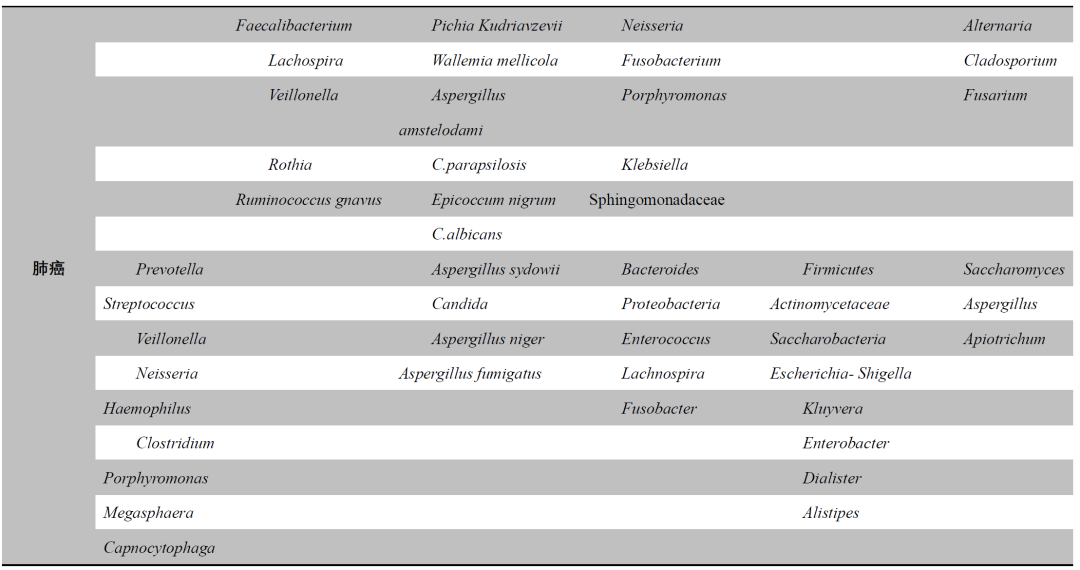
表1. 肺部和肠道的微生物失调会影响慢性阻塞性肺疾病(COPD)、哮喘和肺癌
肺癌
研究表明,肠道微生物在肺癌的发病机制、进展以及调节治疗的免疫应答中,发挥着关键作用。与健康人群相比,肺癌患者的肠道微生物组成明显不同,Firmicutes、Actinobacteria和Bacteroidetes的丰度显著降低而其他门类的丰度显著增加,包括Proteobacteria。多项研究已证实,肺癌患者与健康个体之间肠道微生物存在显著差异,提示肠道微生物有可能成为精准肿瘤学中诊断和治疗的生物标志物。在肺癌患者中,常见的癌症相关细菌包括Clostridium、Escherichia coli、Fusobacterium、Porphyromonas和Bacteroides,其中Clostridium的过度生长被认为可能激活致癌物并与肿瘤发生相关。肺癌患者显示出肠道微生物失调,表现为Actinobacteria和Bifidobacterium的相对丰度减少,以及Enterococci的相对丰度增加,代谢组学分析揭示这些患者的正常肠道微生物功能出现下降。
此外,不同病理类型的肺癌患者的肠道微生物组成有所不同,非小细胞肺癌(NSCLC)患者的肠道中的Rikenellaceae、Prevotella、Streptococcus、Lactobacillus、Bacteroides、Treponema和Enterobacteriaceae的相对丰度高于健康对照组。NSCLC患者还表现出产丁酸盐细菌的减少,而这些有益的肠道微生物生成的丁酸盐可以影响肠道微血管系统中的内皮血管生成,抑制肿瘤细胞的生长。肺腺癌和鳞状细胞癌具有独特的微生物标志物,其中,腺癌患者的肠道中富集了Gemmiger和Erysipelatoclostridium,而鳞状细胞癌患者则富集了Enterococcus、Veillonella和Eubacterium eligens。另一项研究已识别并验证了13个预测肺癌的生物标志物,建立了一个特定的肠道微生物标志物曲线,用于预测早期肺癌(曲线下面积AUC = 97.6%)。Aspergillus sydowii通过分泌IL-1β促进髓源抑制性细胞(MDSC)的增殖和激活,从而促进肺腺癌的进展。定植在肠道中的SFB可以在肺部真菌感染中调节CD4+ T淋巴细胞的极化并增强肺部Th17细胞的抗真菌反应。此外,越来越多的研究认为,针对肠道微生物的治疗策略在癌症预防和治疗改进中有效,Bacteroides ovatus和Bacteroides xylanisolvens的丰度增加可以增强靶向药物厄洛替尼(erlotinib)在肺癌小鼠模型中的疗效。肠道微生物还可以调节免疫检查点抑制剂在肿瘤患者中的疗效。Bifidobacterium还可以招募和激活抗肿瘤T细胞,促进干扰素和促炎细胞因子的产生并促进树突状细胞的成熟,从而增强抗PD-L1单克隆抗体治疗的疗效。
靶向肠-肺轴的治疗策略
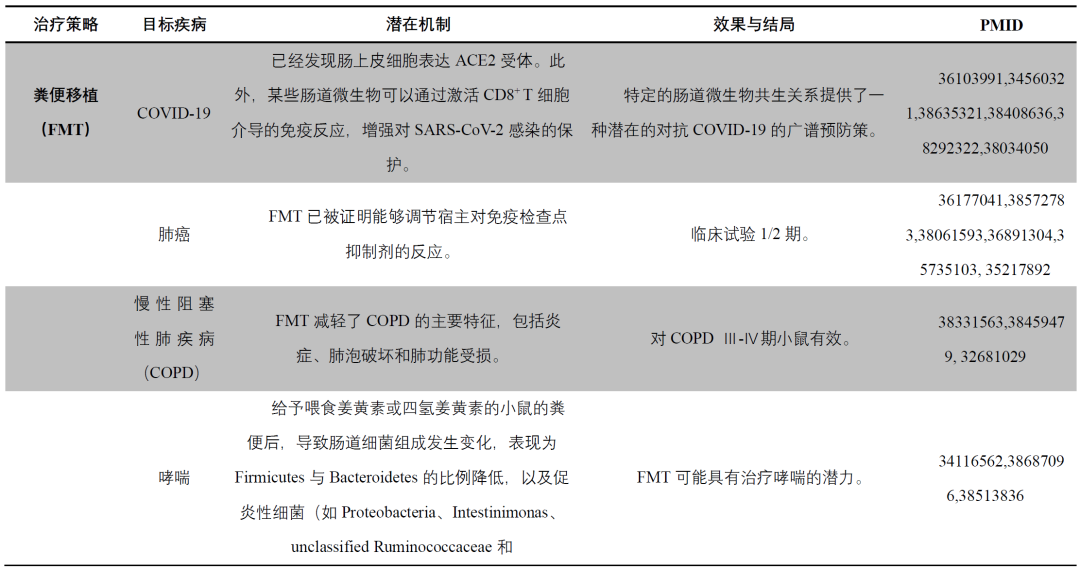
粪菌移植(FMT)
粪菌移植(FMT)对调节受试者的肠道微生物具有良好的前景,已在炎症性肠病(IBD)、肠易激综合症(IBS)或非酒精性脂肪性肝炎的治疗中显示出积极的影响。值得注意的是,在抗生素治疗后,进行FMT处理的小鼠表现出肺动脉高压症状的减轻。有趣的是,胶囊化FMT被认为是治疗肺动脉高压的一种有前途的方法。然而,FMT存在严重感染的风险,特别是在免疫抑制患者中,已有研究表明,FMT后出现了E. coli感染,且有一例导致死亡。因此,在不同疾病背景下,必须对供体进行严格筛查和检测,并对FMT材料进行精细处理和保存(图5)。因此,FMT在治疗疾病时需要谨慎考虑。
饮食
饮食模式是调节肠道微生物的一个关键因素(表2)。临床和实验研究都强调了饮食对于肠道微生物多样性的深远影响。肠道微生物对饮食变化的迅速反应尤为显著,个体在一天之内经历动物性和植物性饮食转变后,微生物组成会发生明显变化。虽然饮食习惯已知会影响肠道微生物的β多样性,但α多样性似乎受到的影响较小,并且在以动物食品为主的人群中呈现出显著的个体差异。流行病学数据表明,富含促炎成分的饮食常与增加的吸烟习惯、慢性阻塞性肺疾病(COPD)和肺功能损害相关。饮食成分可影响肠道微生物的组成,例如,奶酪可能增加Bifidobacteria,减少Bacteroides和Clostridia;人工甜味剂增加Proteobacteria、E. coli并减少Bacteroides、Clostridia和总需氧菌;富含多酚的食物增加Bifidobacteria、Lactobacillus、F. prausnitzii、Roseburia、Bacteroides vulgatus、A. muciniphila并减少E. coli和Enterobacter cloacae;高脂饮食增加Firmicutes并减少Bacteroidetes。高热量饮食通过破坏肠道微生物和Th17/Treg平衡,与脂多糖(LPS)诱导性肺炎加重相关。相反,来自水果、谷物和麦片的膳食纤维显示出对肺癌的潜在保护作用,全谷物也表现出类似的关联。预计未来的预防和支持性疾病管理可能通过调整特定食物的摄入(如未加工乳制品或富含纤维的食物)或改变生活方式(如减少久坐、肥胖和吸烟)来进行饮食调整(图5)。
益生菌
新兴研究强调了肠道微生物改变在管理和缓解肺部疾病中的重要性(表2)。研究揭示了益生菌与抗PD-1治疗联合使用时,对于进展期或复发期的非小细胞肺癌的潜在益处。益生菌能够刺激免疫细胞(巨噬细胞、树突状细胞、中性粒细胞和自然杀伤细胞),增强吞噬作用、细胞因子释放以及Th1/Th17的极化,特别是在肠道黏膜区域。相反,某些益生菌菌株通过上调调节性T细胞(Tregs)、IL-10、TGF-β,促进免疫球蛋白A(IgA)产生,通过调节树突状细胞来增强肠道屏障功能。未来的研究将重点关注特定益生菌菌株的补充周期、递送方式、剂量及随访时间。
表2. 针对肠-肺轴的治疗策略


治疗药物的生物累积
许多药物倾向于在特定的肠道细菌物种中积聚,这些微生物对药物的化学性质不做改变。这种生物累积可能会降低药物效力,改变肠道微生物的组成和功能,调节宿主的生理状态及代谢。药物在肠道细菌中的生物累积通过两种主要方式影响治疗效果:首先,降低药物的生物利用度;其次,改变代谢产物谱。此外,越来越多的证据表明,肠道微生物影响了宿主对于化疗和其他癌症治疗的反应,从而带来三种核心临床意义:增强药物效果、抵消抗癌效果以及诱发药物相关的毒性。肠道微生物与多种化疗药物(如5-氟尿嘧啶、环磷酰胺和甲氨蝶呤)以及新兴免疫疗法(如抗PD-L1和抗CTLA-4治疗)的药理作用之间表现出强相关性。临床医生和转化研究人员可能将在这一领域取得重大进展,将这些见解融入未来的临床试验中。
小结
肠道与呼吸系统之间存在着重要而复杂的相互作用,肠道微生物失调被认为与哮喘、COPD、肺癌和呼吸道感染等常见呼吸系统疾病的发生和发展密切相关(图6)。
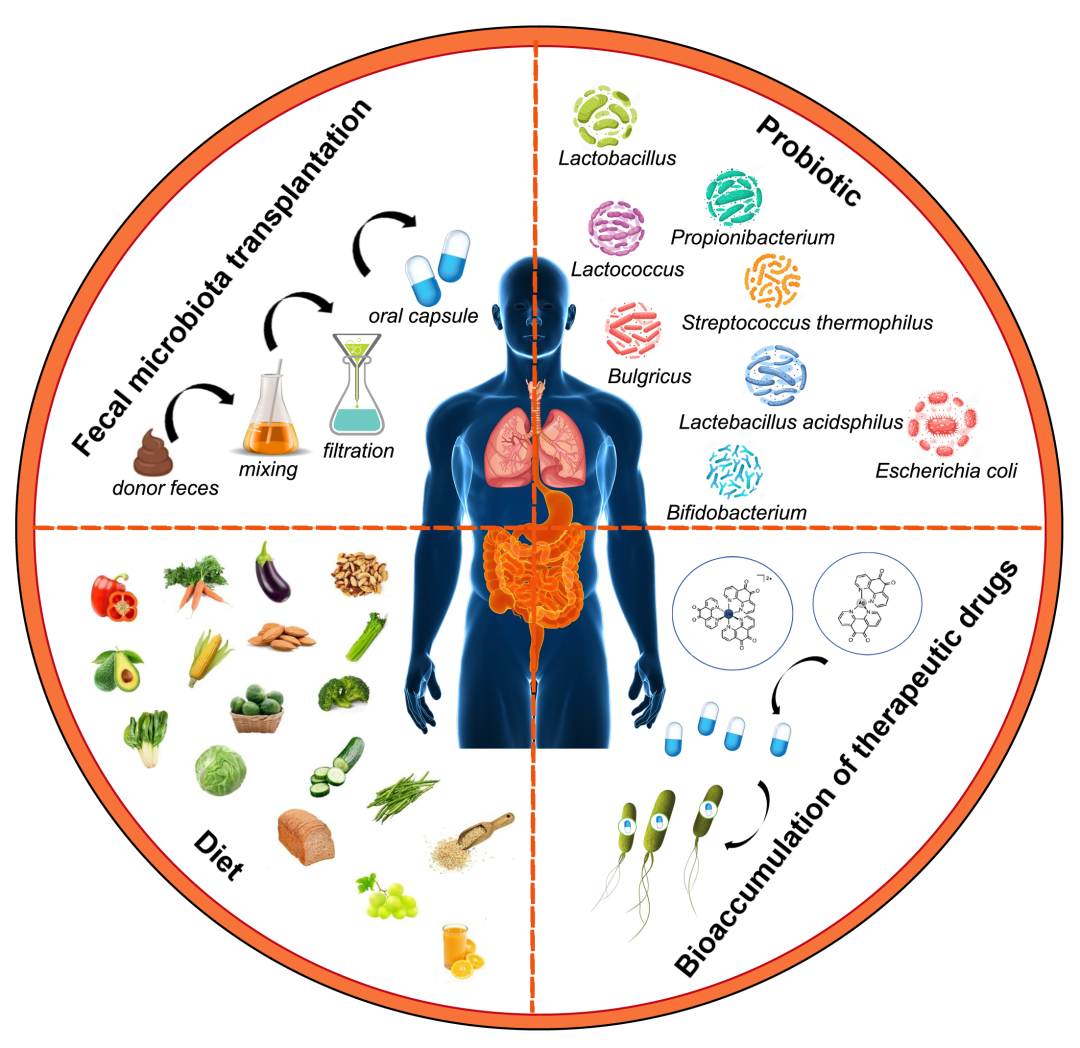
图5. 靶向肠-肺轴的治疗策略
具有前景的靶向肠-肺轴治疗策略包括粪菌移植(FMT)、益生菌、饮食和治疗药物的生物累积。
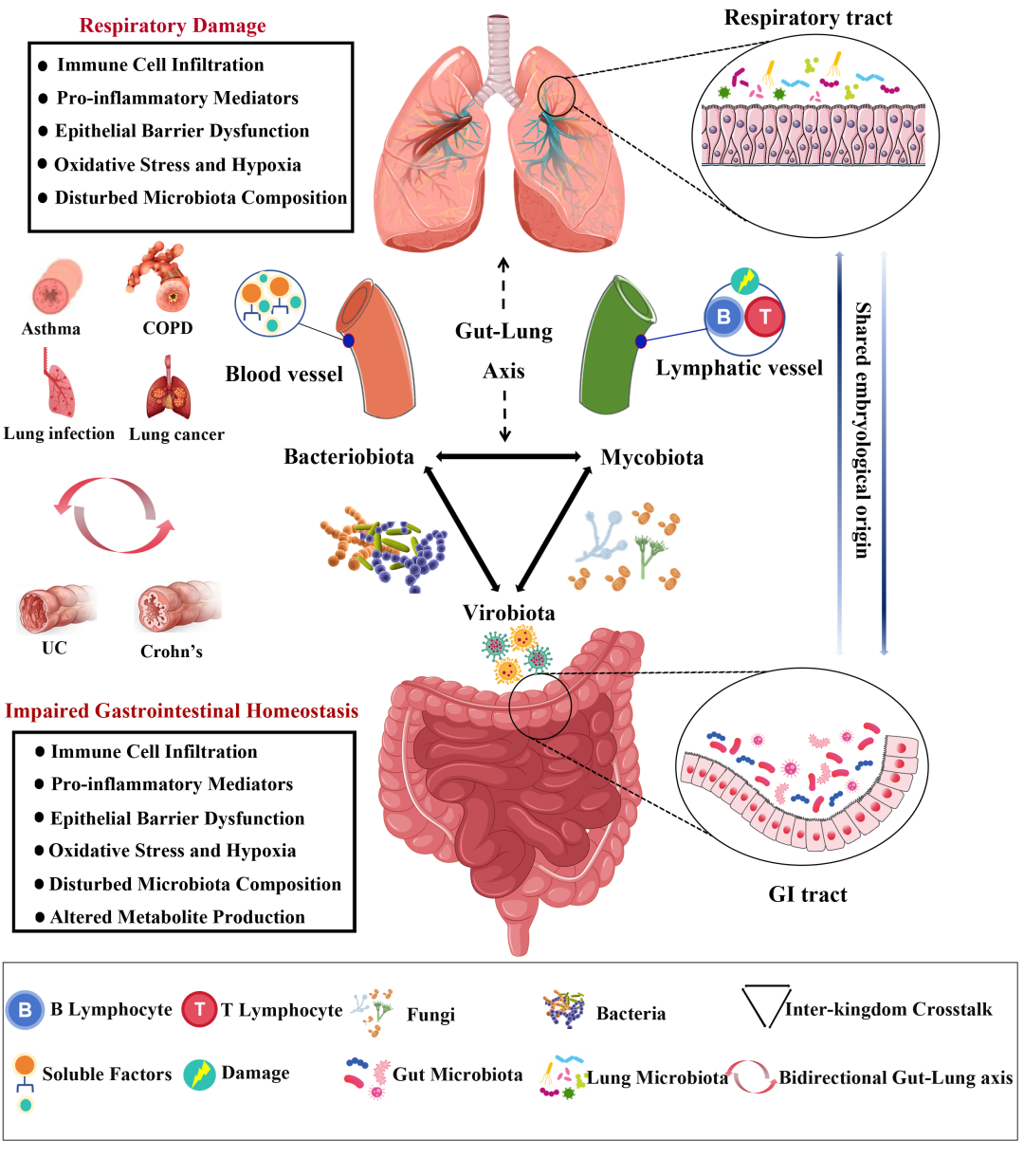
图6. 肠-肺轴:健康与疾病中的双向作用
肠-肺轴涉及细菌、真菌和病毒在各个器官之间的复杂相互作用。肠道与肺部之间的联系在慢性阻塞性肺疾病(COPD)、哮喘、肺部感染和肺癌中尤为明显。COPD表现为肺部炎症、免疫细胞浸润以及促炎介质(如TNF-α和IL-6)水平升高,导致氧化应激和低氧。这些促炎介质和烟草烟雾可以影响肠道,导致免疫细胞浸润和上皮损伤加剧。慢性肠道炎症改变了微生物群,减少了有益菌群。肠道功能受损时,促炎介质水平升高,营养吸收和抗氧化能力下降,病原防御减弱,从而加重肺部疾病。
肠-心轴
概述
心血管疾病(CVD)是全球威胁人类健康的最主要疾病之一。心血管疾病主要包括动脉粥样硬化、高血压、心力衰竭和心肌病。其发病受多种因素的影响,如生活方式、遗传和年龄。
近年来,肠道微生物与心血管疾病之间的联系日益显著。与心血管疾病类似,肠道微生物也受到饮食、生活方式和年龄等因素的影响。这些影响可以改变肠道细菌的组成及其代谢产物,从而影响心血管系统。Hazen等人通过代谢组学识别了一种肠道细菌代谢产物——TMAO,该物质可以预测心血管疾病的风险。当分析老年小鼠和年轻小鼠的肠道微生物变化时,发现肠道微生物的组成随着年龄的增长而发生变化。这些改变的菌群通过TMAO损害心血管功能并加重心血管疾病。TMAO的循环水平与年龄相关,高水平的TMAO会引发氧化应激失衡,导致内皮细胞衰老。
因此,肠-心轴是肠道微生物影响CVD的一个重要途径。该轴受到年龄、饮食以及其他影响心血管疾病的因素的影响,是心血管疾病研究的重要方向。本文将讨论最新的研究,阐明肠道微生物及其代谢产物对心血管疾病发展、机制和治疗方法的影响。
肠-心轴在各种心血管疾病中的作用
菌群代谢产物,例如TMAO、苯乙酰谷氨酰胺(PAGIn)和SCFAs,与CVD密切相关。我们在图7中总结了这些代谢产物(包括N,N,N-三甲基-5-氨基戊酸(TMAVA))通过何种机制影响心血管疾病,列举了与肠-心轴相关的动脉粥样硬化(AS)、心力衰竭、高血压和心肌病的研究。
动脉粥样硬化(AS)
与健康个体相比,动脉粥样硬化(AS)患者的肠道微生物发生了显著变化。肠道微生物是动脉粥样硬化的重要调节因子。肠道微生物代谢产物,如TMAO、PAGIn和SCFA,是心血管效应的主要调节因子。TMAO水平主要受饮食和遗传因素的影响,与动脉粥样硬化的病程呈强的正相关,参与巨噬细胞胆固醇积累和泡沫细胞形成。最近,采用代谢组学方法筛选了与心血管疾病相关的三种磷脂酰胆碱代谢产物(TMAO、胆碱和甜菜碱)。膳食补充的磷脂酰胆碱,通过肠道微生物转化为三甲胺(TMA),TMA是TMAO的前体。TMA随后在肝脏中通过黄嘌呤单加氧酶(FMO1和FMO3)氧化生成TMAO,而TMAO水平的升高会加速动脉粥样硬化的发展。产TMA的L. saccharolyticum会导致ApoE-/-小鼠血清中TMAO水平升高,动脉粥样硬化症状加重。TMAO通过多种机制促进动脉粥样硬化的发展。它刺激血管内皮中的活性氧(ROS)产生,引发血管炎症,抑制超氧化物歧化酶(SOD2)并激活NLRP3炎症小体,从而加剧内皮炎症,促进动脉粥样硬化的发展。此外,TMAO的促动脉粥样硬化作用,与琥珀酸脱氢酶复合物B亚基(SDHb)上调导致ROS水平升高、引发血管内皮细胞焦亡有关(图7)。TMAO还影响胆汁酸的合成,抑制肠道胆固醇的逆向转运,导致胆固醇在巨噬细胞中积累。这与动脉粥样硬化的加重有关。脯氨酸/丝氨酸丰富卷曲螺旋蛋白1(PSRC1)是与免疫反应调节和冠状动脉疾病(CAD)发展相关的关键因子。PSRC1的缺乏,改变了肠道TMA生成菌的丰度和肝脏FMO3的表达,导致血浆TMAO水平升高,加重动脉粥样硬化。然而,抗生素治疗清除了肠道菌群,减轻了这一效应,提示存在依赖肠道菌群的调节途径。除了与TMA生成相关的菌群外,某些微生物,如Enterobacter aerogenes ZDY01,利用TMA作为营养物质,降低血浆TMAO水平。它还促进了胆汁酸的吸收并抑制了肠道FXR-FGF15轴。这促进了肝脏细胞色素P450家族7亚家族A成员1(Cyp7a1)基因的表达,加速了胆固醇向胆汁酸的转化,促进了胆固醇逆向转运。因此,E. aerogenes ZDY01改变了血浆TMAO和胆固醇水平,减轻了动脉粥样硬化的程度。
除了TMAO在肠道微生物与AS之间的作用外,还有其他一些肠道微生物代谢产物也参与了这一过程。PAGln与动脉粥样硬化密切相关,其生成与肠道微生物对苯丙氨酸的代谢有关,肠道微生物通过苯丙酮酸脱羧酶(PPDC)和苯丙酮酸:铁氧还蛋白还原酶(PPFOR)途径将苯丙氨酸转化为苯基乙酸(PAA),然后进入肝脏与谷氨酸结合形成PAGln。PAGln通过肾上腺素受体促进血小板反应性和血栓形成,参与了动脉粥样硬化等不良心血管事件的发展。与PAGIn类似,TMAO也可以导致血栓形成,从而影响动脉粥样硬化。由肠道微生物代谢产生的SCFAs也是肠-心轴中的重要信号分子。丙酸(PA)通过增加IL-10的表达并减少小肠胆固醇转运关键蛋白(NPC1L1)的表达,在动脉粥样硬化中起到缓解作用,从而降低血浆胆固醇水平。同样,产生丁酸的细菌在动脉粥样硬化中也具有保护作用。因此,肠道微生物代谢的变化,可以通过肠-心轴改善动脉粥样硬化的发展。
心肌病
心肌病是一种由多因素引起的心肌损伤,主要表现为心肌肥厚和心室扩张,是心力衰竭的重要风险因素。缺血性心肌病和扩张型心肌病患者的肠道微生物与健康个体有显著不同,这表明心肌病与肠道微生物之间存在明确的联系。这两种心脏疾病都是由不同原因引起的心肌损伤。在一项研究中,16S rRNA基因测序发现,缺血性心肌病和扩张型心肌病患者的肠道微生物中,Proteobacteria显著增加。相反,先前的研究显示,Proteobacteria与包括AS在内的多种心血管疾病密切相关。目前的研究尚未阐明这两种心肌病的主要致病菌和致病机制,这仍是一个值得进一步研究的问题。在糖尿病性心肌病(DCM)小鼠中,发现肠道微生物多样性下降,同时,与支链氨基酸(BCAAs)生成相关的Lachnospiraceae和Clostridiales的丰度增加。研究表明,在1型糖尿病小鼠的糖尿病性心肌病中,肠道中增加的BCAA,通过肠-肝-心轴增加了心脏BCAA转运蛋白LAT1的表达。这导致心脏中BCAA的过剩,进而通过mTOR信号通路引发线粒体损伤和心肌细胞凋亡。心肌缺血/再灌注(I/R)损伤也是导致心脏功能障碍的风险因素。心肌I/R损伤会导致肠道屏障功能受损,增加细菌移位的可能性,从而引发更强的炎症反应,而抗生素治疗后,心肌I/R损伤得到减轻。这些证据表明,肠道微生物与心肌损伤之间存在密切关系。
心力衰竭
心力衰竭(HF)主要表现为心脏泵血功能下降,可以由多种心肌损伤因素引起。心力衰竭的发展与肠道微生物密切相关。心力衰竭导致心脏泵血减少,进而引起肠道缺血。肠道血流减少会影响营养吸收并导致肠道功能障碍。通过16S rRNA基因测序发现,在小鼠心脏负荷压力下,肠道SCFA和色氨酸生成细菌的数量减少。肠道微生物的失调,通过T细胞依赖的方式引起心脏重构和功能障碍,而色氨酸和心脏AhR(芳香烃受体)则表现出显著的心脏保护作用。
菌群代谢物与心力衰竭的进展同样密切相关。TMAO水平与心力衰竭的预后密切相关。在小鼠模型中,TMAO通过TGF-β1/Smad3通路诱导心肌纤维化和心脏肥厚,这表明TMAO在心力衰竭中发挥作用。心肌肥厚是心力衰竭的重要危险因素。在心力衰竭患者中,肠道上皮分泌因子FGF19的水平升高,促使心肌肥厚,进而引起心力衰竭。TMAVA(N-甲基-3-甲基-酪氨酸)也与心力衰竭患者的预后相关。TMAVA抑制肉碱合成和脂肪酸氧化(FAO),导致心肌细胞的代谢重编程,进而引起心肌能量代谢受损(图7)。与TMAVA类似,TMAO也可以导致线粒体功能障碍和心肌能量代谢受损,促进心力衰竭的发展。射血分数保留型心力衰竭(HFpEF)是心力衰竭的一种类型。利用代谢组学分析发现,HFpEF小鼠模型中的吲哚-3-丙酸(IPA)水平下降,提示IPA在HFpEF中可能具有保护作用。IPA的心力衰竭保护作用主要通过AhR(芳香烃受体)的刺激,促进SIRT3表达并抑制NNMT(烟酰胺N-甲基转移酶),从而增加NAD+水平。此外,肠道微生物产生的代谢物犬尿氨酸(kynurenine)也与心力衰竭相关,也通过AhR激活促进心肌纤维化的进展。心肌梗死后的心肌纤维化是心力衰竭的一个危险因素,肠道中的产丁酸细菌在这一过程中起着重要作用。丁酸通过抑制组蛋白去乙酰化酶(HDAC)活性,促进心肌梗死后的组织修复。然而,产丁酸细菌的减少会促进心肌梗死后的心肌纤维化和心脏功能障碍。
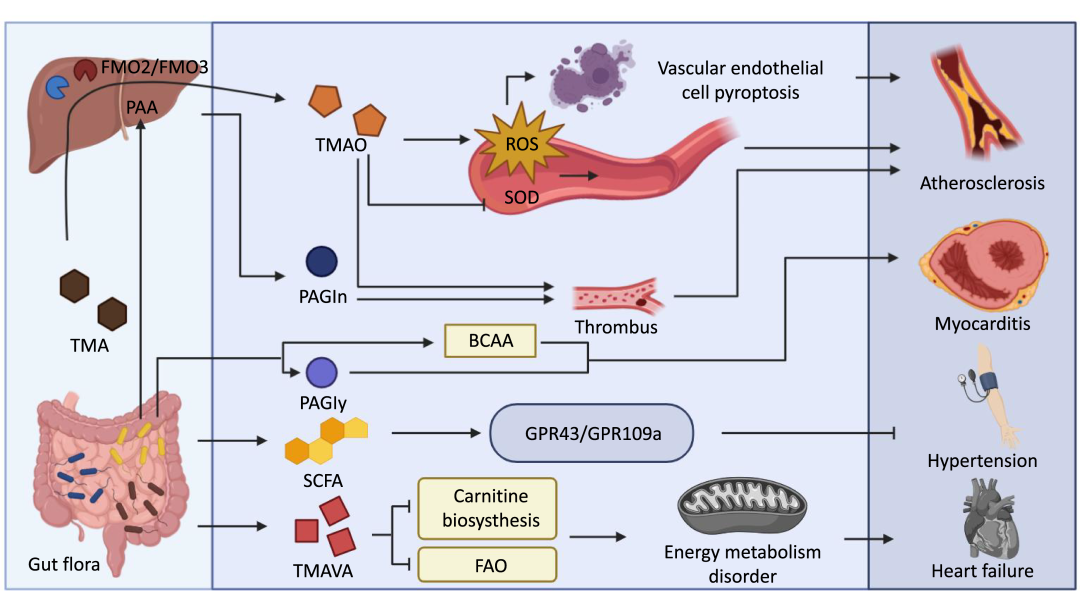
图7. 肠道微生物组代谢产物(TMAO、PAGIn、SCFA和TMAVA)通过多种机制影响心血管疾病的发展
肠道微生物产生的TMA在肝脏中转化为TMAO,TMAO通过诱导氧化应激和NLRP3的产生,促进血管内皮细胞的焦亡,从而加重动脉粥样硬化(AS)。PAGIn是另一种肠道代谢产物,通过加速血栓形成,从而加重动脉粥样硬化。PAGIy和支链氨基酸(BCAA)是与心肌病密切相关的两种细菌代谢产物。此外,肠道微生物产生的短链脂肪酸(SCFA)被发现通过GPR43/GPR109a影响高血压的进展。最后,细菌代谢产物TMAVA可以导致能量代谢障碍,从而影响心力衰竭。(本图由BioRender.com制作)
高血压
高血压是一种多因素心血管疾病。最近的一项研究,比较了41名健康对照者、56名高血压前期患者和99名原发性高血压患者的肠道微生物组成,结果显示,高血压前期和高血压患者的肠道中产生SCFA的细菌,如F. prausnitzii和 Roseburia 的丰度减少。将高血压患者的粪便移植给无菌小鼠后,观察到小鼠血压升高,这表明肠道微生物在高血压发展中起着重要作用。高盐饮食是高血压的危险因素之一。在高盐饮食下,Lactobacillus murinus的丰度在人体和小鼠中都减少。Bacteroides fragilis通过其代谢物花生四烯酸抑制高盐饮食引起的肠源性皮质酮的产生,从而减轻高血压症状。调节类固醇激素可能是肠道微生物影响高血压的一种方式。此外,其他肠道微生物代谢物,如TMAO和SCFA,也会影响高血压症状。高血压患者的血浆TMAO水平升高,TMAO通过促进血管收缩,参与Ang II诱导的高血压。SCFA是肠道微生物发酵纤维产生的产物,它们与心血管系统密切相关。SCFA的心脏保护作用通过其受体GPR43/GPR109a介导,SCFA的减少会削弱这一信号通路,从而导致高血压。因此,粪便中的SCFA、进入血液循环的SCFA与心血管疾病之间的关系值得进一步阐明。此外,肠道微生物对心血管系统的调节还涉及免疫反应的调控。乳糜泻(CeD)是一种自身炎症性肠病,会导致肠道细胞释放IL-17并激活NLRP3炎症小体。这些炎症因子进入血液循环后,会引发一系列问题,包括增加心血管风险和动脉高血压。
基于肠-心轴的心血管疾病治疗策略
目前针对肠-心轴的治疗手段主要通过饮食调节、益生菌、药物以及其他影响肠道微生物及其代谢物的因素来实现治疗心血管疾病的目的。饮食调节是影响肠道微生物组成和代谢物产生的因素之一。众所周知,高脂肪和高盐饮食是心血管疾病的重要危险因素。通过改善饮食治疗心血管疾病具有很大的前景,尤其是地中海饮食的心脏保护特性。摄入高纤维食品与肠道微生物的代谢作用相结合,能够促进多种具有心脏保护作用的SCFAs产生。在高纤维食品喂养后,小鼠肠道中产生乙酸的细菌丰度增加,心脏肥厚和高血压症状得到了改善。此外,补充乙酸也显著改善了小鼠的心脏肥厚和高血压症状。类似地,外源性补充丙酸显著改善了主动脉瘤的发展。除了改变摄入的食物外,禁食也被证明具有心血管保护作用。间歇性禁食(IF)通过改变肠道微生物而降低血压,其效果主要通过胆汁酸信号通路介导。禁食的效果还包括对Th1细胞和树突状细胞的免疫调节作用,进而发挥降血压作用。目前,阻止高血压的饮食方案(DASH饮食法)是国际公认的治疗高血压的饮食方法。当与禁食结合时,DASH饮食具有更好的降压效果。
益生菌通过改变肠道微生物的组成来产生心血管保护作用。Lactobacillus rhamnosus和Bifidobacterium lactis是两种具有降血压作用的益生菌。它们通过影响脂质代谢、血管平滑肌收缩和类固醇激素合成来发挥降血压作用。用Lactobacillus治疗高血压大鼠,显著降低了血压。有趣的是,通过粪便组织学研究发现,F. prausnitzii与CVD显著相关并且具有显著的抗动脉粥样硬化作用。因此,补充F. prausnitzii是一种有前景的动脉粥样硬化益生菌治疗策略。Akkermansia muciniphila在患有主动脉瘤的小鼠中显著减少,口服补充能够减缓主动脉瘤的发展。其主要机制包括抑制炎症、恢复肠道微生物的结构多样性以及调节Lactobacillus的功能。口服Saccharomyces boulardii能改善心力衰竭患者的射血功能。
肠-骨轴
概述
骨骼稳态依赖于破骨细胞的骨吸收和成骨细胞的骨形成之间的平衡,这一过程被称为“骨重塑”。多种因素导致骨质疏松症,其中之一就是肠道微生物。肠道微生物通过影响多个与骨骼相关的因素来调节骨代谢,即“肠-骨”轴(图8)。
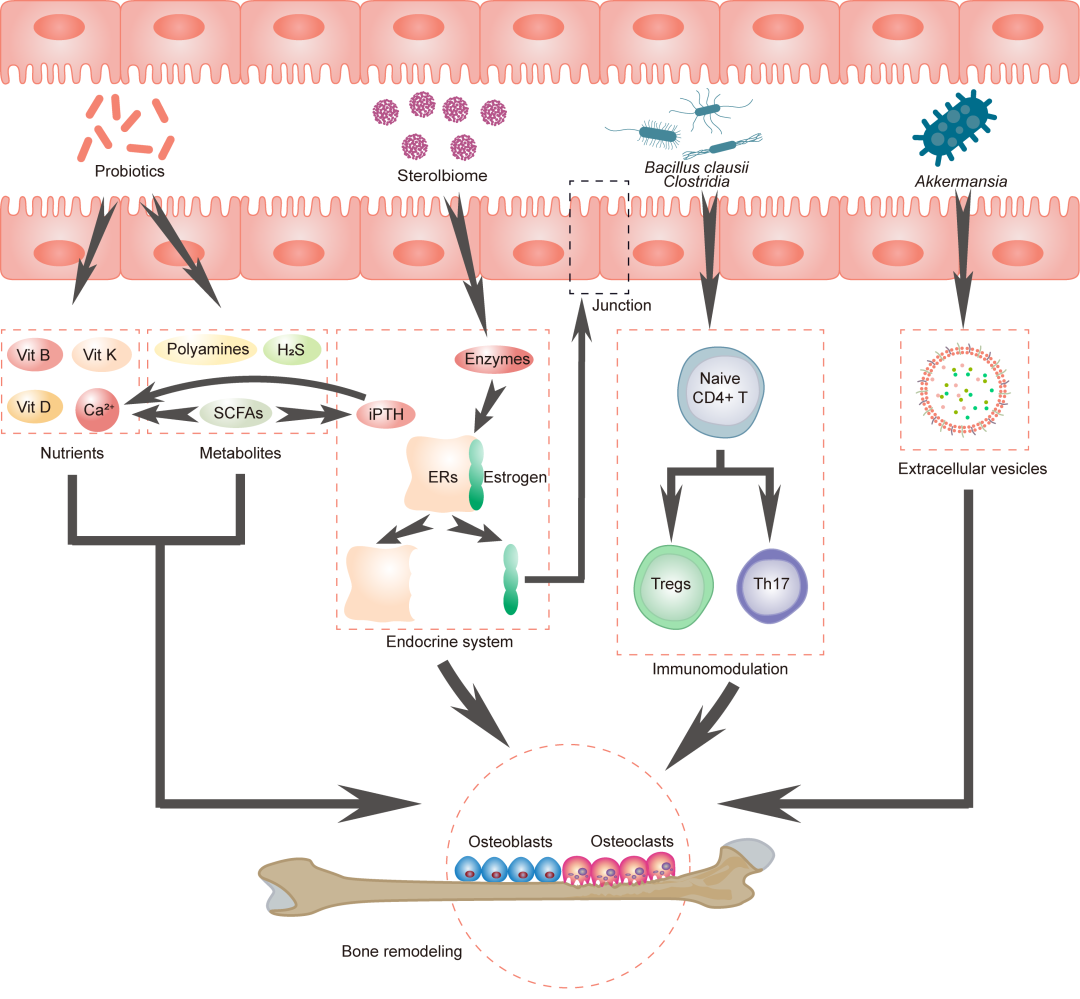
图8 肠道微生物调节骨代谢的潜在机制
肠道微生物,特别是益生菌,促进营养物质(如钙、维生素D、B和K)的产生和/或吸收,维持骨骼健康。此外,肠道微生物产生短链脂肪酸(SCFAs)、多胺、硫化氢(H2S)和细胞外囊泡,影响Tregs和Th17细胞之间的平衡,通过甾醇生物酶从雌激素受体(ERs)中释放雌激素,通过SCFAs介导间歇性甲状旁腺激素(iPTH)的作用。所有这些机制通过影响成骨细胞和破骨细胞前体的分化,最终调节骨重塑,进而调控骨代谢。
肠道微生物与骨骼密切相关
十多年来的研究发现,肠道微生物与骨骼密切相关。无菌小鼠的骨量较高且破骨细胞较少,相比之下,常规饲养的小鼠骨量较低;引入普通小鼠的肠道菌群,可使无菌小鼠升高的骨量变为普通水平。此外,去卵巢(OVX)小鼠去除了肠道菌群后,通过G蛋白偶联胆汁酸受体介导,改善了骨量、骨微结构和骨强度。在中国多民族研究队列中,“海拔-菌群-定量超声指数”分析揭示了高海拔暴露、肠道微生物与骨密度(BMD)之间的负相关关系;此外,研究还发现了Catonibacterium在这一相关性中的中介作用。通过使用来自TwinsUK、LifeLines-DEEP和UK Biobank队列的数据进行孟德尔随机化分析,发现肠道微生物与骨骼发育之间存在因果关系。其他综述文章也探讨了肠道微生物与骨骼之间的相关性,此处不再详细阐述。
肠道微生物调节骨代谢的潜在机制
营养吸收
肠道微生物的丰度、多样性和组成的变化会影响膳食摄入,特别是营养吸收,进而通过肠-骨轴影响骨代谢。例如,钙在维持人体骨骼健康方面起着至关重要的作用。补充益生菌,特别是Lactobacillus菌株,增强了小鼠钙的运输和吸收。此外,一项针对绝经后女性的临床试验显示,食用Lactobacillus helveticus发酵奶后,血清钙水平有所提高。补充半乳寡糖(益生元)则增加了钙和镁的吸收,并改善了BMD。益生菌和益生元增强钙吸收和生物利用度的机制包括上调钙转运蛋白的表达,增加细胞密度、肠隐窝深度和血流量以及改变肠道微生物的组成和完整性。
肠道微生物还调节维生素D的代谢,而维生素D代谢反过来影响肠道中钙和磷的吸收以及骨代谢。无菌小鼠的维生素D代谢受损,但在肠道微生物定植后,1-25(OH)2D和钙的水平得到了恢复。口服补充益生菌Lactobacillus reuteri可以提高循环中的25(OH)D水平。维生素D受体在胃肠道中显著表达,维生素D激活其受体以维持肠上皮屏障并促进肠道微生物的稳态。
肠道微生物对于合成维生素B和K也至关重要,而这两种维生素对维持骨骼健康非常重要。维生素K通过促进组织特异性维生素K依赖蛋白(如骨钙素)的γ-羧化作用,刺激成骨细胞上的外源性受体,进而影响骨重塑和骨矿化过程,最终增强骨骼强度。在髋部骨折患者中,已观察到维生素B的摄入不足;多项观察性研究还表明,某些维生素B(如B2、B6、叶酸或B12)与降低骨质疏松症或髋部骨折风险之间存在相关性。
代谢物
肠道微生物通过产生被称为“后生元”(postbiotics)的代谢物,有益地影响远端器官。与肠道微生物相关的代谢物的变化,与骨质疏松症的病情进展密切相关。例如,肠道微生物发酵膳食中不可消化的碳水化合物,在肠腔中生成SCFAs(例如丙酸、丁酸和戊酸)。SCFAs能够降低肠道中的pH值,从而增加钙、磷和镁等矿物质的溶解度并促进其吸收。丙酸和丁酸通过减少破骨细胞相关基因TRAF6和NFATc1的表达,抑制破骨细胞前体分化和骨吸收,显著提高卵巢切除小鼠的骨量。此外,戊酸通过抑制NF-κB信号通路来阻止破骨细胞前体分化,起到防止绝经后骨质流失的保护作用。
多胺是另一种代谢物,主要通过肠道微生物的氨基酸转氨基反应合成,特别是精氨酸,在催化酶的帮助下完成。多胺增强成骨基因(如ALP、RUNX2、OCN)的表达,从而促进人类骨髓来源间充质干细胞(BMSCs)的细胞外基质矿化和成骨作用。此外,多胺还作为破骨细胞生成的抑制剂;例如,定期食用富含多胺的酿酒酵母Saccharomyces cerevisiae S631可防止OVX小鼠的破骨细胞激活和骨质流失。精氨酸作为多胺合成的前体之一,与“肠-骨”轴相关。Lachnospiraceae通过产生L-瓜氨酸并将其转化为L-精氨酸,有助于小鼠的骨机械适应。L-精氨酸通过激活包括一氧化氮和钙在内的正反馈环路,增强骨机械适应作用。
硫化氢(H2S)是另一种参与肠道微生物介导的骨重塑的重要代谢物。它通过硫酸盐还原菌分解半胱氨酸生成。H2S对骨形成和出生后骨骼的发育至关重要,因为它通过Wnt/β-catenin信号通路支持BMSCs的自我更新和成骨分化。H2S不足会导致持续的骨质疏松,表现为BMSCs功能受损和骨形成受阻。
免疫调节
肠道微生物与宿主免疫系统之间的相互作用始于婴儿期。根据“骨免疫学”理论,Tregs和Th17细胞之间的平衡对于维持骨骼稳态至关重要。Treg细胞通过抑制单核细胞向破骨细胞分化,诱导CD8+T细胞生成Wnt配体(Wnt10b),从而促进成骨细胞的生成。相反,Th17细胞诱导促炎细胞因子的产生并抑制抗炎因子的作用,这促进了破骨细胞的形成,从而增加骨质流失。
肠道微生物在影响CD4+ T细胞分化为Tregs或Th17细胞,从而调节骨代谢方面,发挥着关键作用。例如,Bacillus clausii显著增加CD4+ Foxp3+ Treg细胞的数量,同时减少骨髓中CD4+ Rorγt+ Th17细胞的比例,从而防止小鼠卵巢切除后引起的骨质流失。另一方面,SFB促进了Th17细胞的增殖和肠道中IL17的产生,而Clostridia能增强Treg细胞群体。此外,其他细菌种类也有调节Treg和/或Th17细胞的潜力,相关内容已在其他已发表的综述中提到。
内分泌系统
“类固醇组群(sterolbiome)”一词描述了一个肠道微生物特殊群体,它们通过改变胆固醇衍生化合物,直接参与调节宿主性激素水平。类固醇组群包含一些酶,如β-葡萄糖苷酸酶、β-葡萄糖苷酶、羟类固醇脱氢酶和硫酸酯酶,这些酶能够从雌激素受体中释放雌激素并增强其在肠道中的再吸收;因此,肠道微生物能调节雌激素代谢并影响局部和全身的雌激素水平。雌激素能直接和间接地调节骨代谢和功能。缺乏雌激素会导致肠道微生物驱动的肠道紧密连接蛋白的表达减少、肠道通透性和炎症增加。
甲状旁腺激素(PTH)对于出生后的骨骼发育也至关重要,因为它调节钙平衡。持续的甲状旁腺激素作用(cPTH)是导致骨质疏松和骨折的常见因素,而且通过与肠道微生物的相互作用而介导该效应。SFB使cPTH进一步放大炎症效应,导致肠道中TNF+ T细胞和Th17细胞水平升高。这些细胞随后从肠道迁移到骨髓,最终促进骨质流失。相反,间歇性甲状旁腺激素作用(iPTH)则诱导骨生成效应,这一过程依赖于肠道微生物产生的丁酸。通过与树突状细胞上的GPR43结合,丁酸促进Treg细胞的分化;随后,Treg细胞促进骨髓中CD8+ T细胞表达Wnt10b,从而促进骨形成。
细胞外囊泡
由细菌释放的细胞外囊泡(EVs)作为一个物种间通讯的形式,有其特殊性。肠道微生物通过这些囊泡作用于骨骼。例如,来自儿童的肠道攻群或A. muciniphila可以防止小鼠卵巢切除去引起的骨质疏松。这一保护作用是通过分泌细胞外囊泡实现的,这些囊泡能够渗透并聚集在骨组织中,抑制破骨细胞生成并刺激成骨作用。类似地,来自Proteus mirabilis的细胞外囊泡通过促进破骨细胞中的线粒体依赖性凋亡途径来保护骨骼免受骨质流失。
益生菌与骨质疏松症治疗
当摄入足够的益生菌时,能带来积极的健康效益。许多动物模型研究已证明益生菌有治疗骨质疏松症的潜力。例如,经过Lactobacillus发酵的酸奶和脱脂奶,能对骨骼健康产生积极影响。Bifidobacterium通过阻止破骨前体细胞的分化以及加速骨折小鼠的骨痂生长,防止卵巢切除引起的骨质流失。
目前只有少数研究报道了益生菌可以直接或间接地影响人体骨代谢。例如,补充钙和短链果糖寡糖(一种益生元)已被证明能够降低绝经后女性中Ⅰ型胶原C端肽(CTX-I)的水平;CTX-I是骨代谢的标志物,反映骨吸收情况。研究发现,Lactobacillus reuteri能够减少低骨密度的老年女性的骨质流失。此外,在牛奶中补充Lactobacillus casei Shirota可以改善桡骨远端骨折老年患者的骨折相关症状(如握力、疼痛和活动范围)。多种益生菌的补充对于骨生物标志物有利,导致患有骨质疏松的绝经后女性中,骨特异性碱性磷酸酶、CTX-I、TNFα和PTH的水平降低。
肠-皮肤轴
概述
定植在肠道和皮肤的微生物在维持屏障稳态方面发挥着关键作用,而肠道微生物失调会影响皮肤健康。在本文中,我们讨论了肠道微生物与皮肤之间的关系,并总结了肠道微生物对不同皮肤疾病的影响,包括特应性皮炎(AD)、牛皮癣(Ps)、痤疮、玫瑰痤疮、头皮屑和脂溢性皮炎、化脓性汗腺炎以及皮肤癌。我们回顾了肠道如何通过不同机制影响皮肤,以增进对肠-皮肤轴的理解。最后,我们基于现有研究结果,提供了有关如何通过调节肠道微生物改善皮肤健康的展望,这将为未来皮肤疾病的治疗提供启示。
皮肤与微生物
皮肤是生物体中最大的器官,作为与外部环境的重要界面,涵盖了生物体的整个外部表面,通过口腔和肛门的粘膜与消化系统建立联系。皮肤主要由表皮和真皮组成。表皮由五层角质形成细胞构成:基底层、棘层、颗粒层、透明层和角质层。真皮主要是由纤维-胶原-弹性组织组成。皮肤是抵御感染和损伤的第一道防线。皮肤具有强大的过滤作用,其表面被共生菌群定殖,从而抑制病原体的入侵。皮肤及其附属结构的完整性是维持皮肤屏障稳态和防止感染的关键。作为身体内部环境与外部环境之间的屏障,皮肤受到空气污染、紫外线、卫生条件等外部因素的影响,同时也受到炎症、创伤、衰老和全身性疾病等内部因素的影响。
皮肤共生微生物随时间而变化,初生婴儿的皮肤微生物在出生后的前六个月内迅速变化,12个月后变化速度减缓。在青春期,由于性激素的作用,皮肤微生物发生显著变化,直至成年期稳定下来。成年人的皮肤微生物相对稳定,但不同部位之间的变化很大。不同体质指数(BMI)和饮食习惯的个体之间,皮肤微生物也存在显著差异,这可能影响皮肤疾病的治疗效果。因此,肠道对皮肤微生物稳态的影响可能为皮肤疾病的治疗提供新的临床解决方案。
肠道和皮肤都富含血管和神经组织,这促进了身体与外部环境之间的沟通。在临床实践中,胃肠道疾病常常与皮肤疾病同时发生。类似地,像牛皮癣这样的疾病,可能与肥胖或炎症性肠病(IBD)等共患病存在相关性。肠道屏障受损会导致致病微生物的定植,而它们的代谢产物通过循环影响皮肤的表皮屏障。肠道微生物与皮肤之间这种相互联系和紧密关系被称为“肠-皮肤轴”(图9和图10)。
肠道微生物由细菌、真菌、病毒和原生动物组成。内部和外部因素都会影响个体的肠道微生物的平衡,从而影响皮肤的健康。例如,饮食通过影响某些肠道微生物的丰度来影响宿主的皮肤微生物。补充外源性益生菌可以改善因肠道微生物失调,产生过量酚类物质而导致的皮肤屏障损伤。高脂饮食或酒精摄入会破坏肠道微生物的平衡并导致皮肤创伤愈合受损。此外,Bifidobacterium longum调节色氨酸代谢并减轻特应性皮炎(AD)症状;它还促进神经酰胺的合成并改善衰老小鼠的皮肤屏障损伤。
皮肤微生物包括细菌、真菌、病毒和噬菌体。每个人的皮肤上都有独特的微生态,这种微生态影响着皮肤的结构和功能,但不会渗透到表皮。近年来,越来越多的关注点集中在皮肤微生物在维持皮肤屏障中的作用上,而且,一些皮肤疾病可能会因肠道微生物的不平衡而加重。然而,需要进一步的研究以理解肠道和皮肤微生物群之间复杂的相互作用。
AD患者表现出肠道微生物多样性和物种的组成发生改变,特征是益生菌的减少和致病菌的增加。补充外源性Bifidobacterium longum可以增强吲哚-3-醛的释放,从而激活AhR受体。这种激活抑制了异常的Th2免疫反应并缓解了AD症状。AhR受体的激活还会引起氧化应激并调节Tregs。靶向AhR受体激活是治疗AD的一种策略。此外,过量摄入脂肪酸以及减少水果和蔬菜的摄入会影响AD的进展,进而影响Treg细胞和Th2型免疫反应。AD患者肠道中Coprococcus的丰度减少导致丁酸盐的产生及其抗炎作用的降低。抗生素引起的肠道微生物群失调,通过减少SCFAs的生成而加重了AD,导致Treg和ILC3的浸润减少。相反,长链饱和脂肪酸(LCSFA)会在肠道中诱导炎症性ILC3的增加,这些细胞迁移到皮肤并触发AD。此外,肠道衍生的SCFAs增强角质形成细胞的线粒体代谢,从而在实验性AD中保护皮肤屏障。
银屑病是一种以皮肤任何部位的鳞屑和增厚为特征的系统性疾病,属于Th17介导的疾病。除了皮肤损伤外,其他器官也会发生炎症。例如,银屑病患者通常表现为较高的BMI并且常伴有高血压、2型糖尿病或IBD。肠道微生物失调可能会触发免疫反应,导致银屑病的易感性。银屑病患者肠道中Firmicutes的丰度增加,而Bacteroidetes的丰度减少。银屑病患者Akkermansia和Faecalibacterium的丰度降低,导致SCFAs生成减少,而SCFAs作为抗炎因子,维持上皮屏障的完整性。补充Leuconostoc mesenteroides可降低IL-17的产生并通过增加血清脱氧胆酸缓解银屑病。摄入Bifidobacterium或Lactobacillus可产生SCFAs并抑制IL-23/Th17轴相关的炎症因子。此外,银屑病还会减少小鼠肠道中的B细胞,导致肠道失调并加重肠炎。
痤疮是一种多因素疾病,与饮食、宿主免疫状态和皮肤微生物组的稳态有关。痤疮的发生与饮食因素高度相关。痤疮患者的肠道微生物多样性和物种丰度减少。中度严重的痤疮与高脂和高糖饮食密切相关。这种关联可能是由于高脂和高糖引发了类固醇调节元件结合蛋白1(SREBP-1)的激活,从而导致皮脂中脂肪酸合成的增加。因此,饮食因素引起的肠道微生物失调会影响痤疮的进展。
玫瑰痤疮是一种慢性炎症性皮肤疾病,其特征为神经血管失调和免疫功能受损,其主要原因是异常炎症因子和抗微生物肽的释放。玫瑰痤疮与肠道微生物之间存在一定的关联,因为玫瑰痤疮患者对幽门螺杆菌感染更为敏感,而幽门螺杆菌感染又促进了胃肠道疾病的发展。此外,玫瑰痤疮患者的肠道微生物丰度降低,这可能通过硫代谢、钴胺素利用和碳水化合物运输而影响疾病的进展。特定的食物和饮料会加重玫瑰痤疮的症状,这可能与瞬时受体潜在的阳离子通道的激活有关,从而导致神经源性血管扩张。
头皮屑、脂溢性皮炎与肠道微生物失调之间的关联仍存在争议,一些证据表明,高脂和高糖饮食可能会加重头皮状况。此外,补充益生菌可以缓解头皮屑和脂溢性皮炎。饮食诱导的肠道微生物失衡与脓肿性汗腺炎的进展相关,而这可能导致IBD的加重。由Bifidobacterium longum产生的神经酰胺通过肠-皮肤轴增强老年小鼠的皮肤屏障完整性。此外,饮食调节和益生菌补充对于黑色素瘤免疫治疗有积极影响。有趣的是,咖啡与皮肤基底细胞癌的恶化风险呈负相关。肠道微生物在这些过程中的作用值得进一步研究。
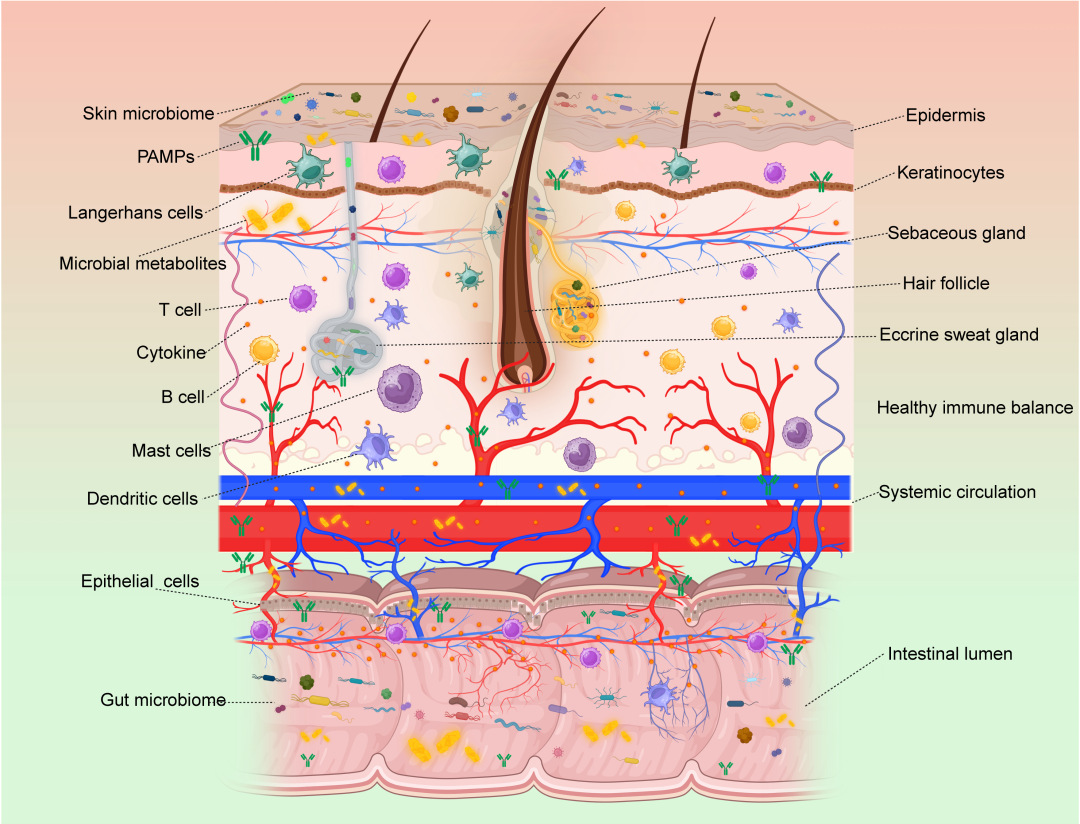
图9. 肠道与皮肤之间的相互作用
肠道和皮肤的表皮屏障通过全身循环相连。肠道(肠道微生物和肠道炎症)和皮肤(免疫失衡、皮肤微生物群和角质形成细胞增殖)的失调是相互关联和相互影响的。
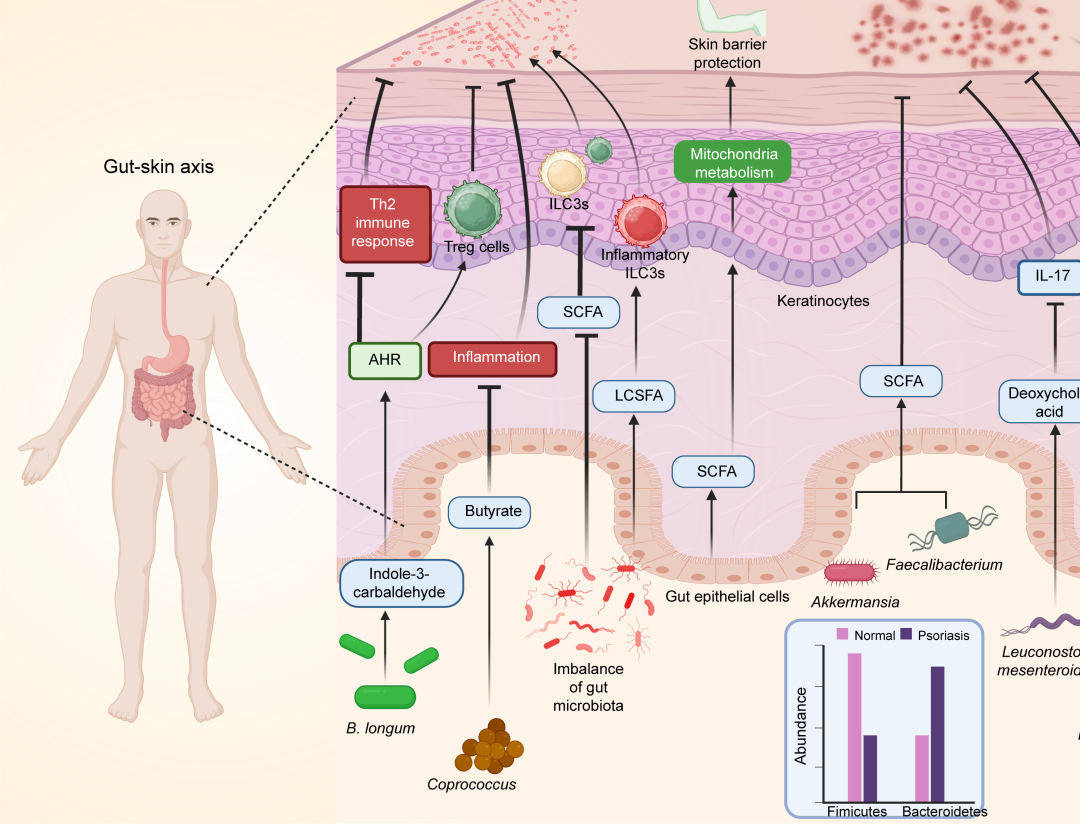
图10. 皮肤疾病与肠道系统之间的联系
不同疾病(如特应性皮炎、牛皮癣、痤疮、玫瑰痤疮及其他皮肤屏障损伤)的详细描述详见正文。
临床应用
改善肠道微生物可以调节皮肤健康,队列研究中已证实,摄入益生菌和益生元能有效修复皮肤屏障。此外,健康的饮食对于皮肤疾病也是有益的。最新的临床研究表明,粪菌移植(FMT)后,AD的症状显著缓解,而喂养益生菌则使小鼠的毛发质量改善。因此,针对肠道微生物的调节成为了管理皮肤问题的一种新方法。
肠-皮肤轴从新的视角强调了肠道微生物在皮肤健康中的关键作用。未来,在小鼠皮肤中采用特定的微生物代谢物标记技术,将得到关于肠道微生物直接影响皮肤生理的有力证据。同时,已识别的益生菌和益生元在日常护肤产品中的应用,具有不可估量的临床转化价值。例如,各种肠道微生物的代谢物目前正被添加到化妆品中。进一步明确肠道微生物在皮肤屏障中的特定角色,为皮肤健康管理制定个性化的治疗措施,不久后将具有临床应用前景。
肠-生殖轴
概述
肠道微生物群的稳态通过与雌激素等性激素相互作用,在女性不同阶段的生殖健康调控过程中发挥着关键作用。微生物群的组成和功能异常会导致许多生殖疾病的发生和发展,包括多囊卵巢综合症、早发性卵巢功能不全、子宫内膜异位症以及妊娠相关并发症。尽管许多研究已经证明了肠道微生物群失调与生殖障碍之间的关联性,但肠道微生物群失调与生殖功能障碍之间的因果关系仍然不清晰。未来需要进行更多研究,以揭示肠道微生物群失调的确切病理效应及其与生殖疾病之间的分子机制,这将极大促进新型治疗手段与预防策略的开发。
人类肠道-性激素轴
人类肠道-雌激素轴
肠道微生物对宿主激素水平的调节至关重要,尤其是雌激素。抗生素的使用,证明了肠道微生物与雌激素水平之间的紧密联系,即抗生素显著降低了女性体内循环的雌激素水平。此外,肠道微生物通过“雌激素组(estrobolome)”参与雌激素水平的调节,该组包含了编码肠道微生物中雌激素代谢酶的基因。肠道微生物分泌的β-葡萄糖醛酸酶可以将结合型雌激素代谢为非结合型雌激素,从而影响血液中的活性雌激素水平。肠道微生物失调和肠道微生物多样性的减少会降低β-葡萄糖醛酸酶的活性,进而导致系统性雌激素水平的改变,导致雌激素依赖性疾病的发生,例如心血管疾病和骨代谢相关疾病。当β-葡萄糖醛酸酶活性升高时,外周血液中的雌激素水平增加。这一系列事件参与了子宫内膜异位症和雌激素依赖性肿瘤的发展。此外,肠道微生物还通过合成SCFAs参与雌激素代谢。丁酸,作为肠道微生物合成的主要短链脂肪酸之一,能够调节猪卵巢颗粒细胞中的雌激素合成。综上所述,这些结果表明“肠道-雌激素轴”在调节女性生殖健康和生理功能中有重要作用。因此,肠道微生物失调在女性生殖疾病发展中的作用受到了广泛关注。
人类肠道-孕激素轴
四氢孕酮(THP)是孕激素之一,在女性的许多生理过程中发挥关键作用(特别是在月经周期和妊娠期间),并且可以通过宿主胆汁中丰富的糖皮质激素(四氢脱氧皮质酮,THDOC)转化生成;这一转化过程是在人类肠道微生物,如Gordonibacter pamelaeae和Eggerthella lenta作用下,通过21-脱羟基而实现。此外,人类肠道微生物,如E. coli Nissle 1917(EcN)产生的氢气也能促进THDOC的21-脱羟基过程。与非孕妇对照组相比,孕妇的粪便中THP水平显著升高,同时,G. pamelaeae和E. lenta的丰度也有所增加。上述结果表明肠道微生物在孕激素代谢中的重要作用,确认了肠道-孕激素轴对女性生殖功能的关键影响。
肠道微生物与多囊卵巢综合症(PCOS)
PCOS是最常见的生殖内分泌和代谢性疾病之一,全球约有6%–18%的育龄女性受到该病的影响。PCOS表现为高雄激素(如多毛症和/或高雄激素血症)以及卵巢功能障碍(如无排卵、少排卵和/或多囊卵巢),在排除其他特定诊断(如高泌乳素血症和非典型先天性肾上腺增生)后可确诊。PCOS患者常伴有全身代谢综合征,如胰岛素抵抗、肥胖、全身性炎症状态,以及其他心血管危险因素,是排卵性不孕的主要原因。PCOS还是早发型2型糖尿病和精神疾病发展的重要促成因素。PCOS的发病机制复杂,临床表现有显著的异质性。PCOS是一种由多基因、表观遗传学和环境因素共同作用的疾病,其发病机制尚未完全阐明。肠道微生物失调在代谢性和内分泌相关疾病中的作用引发了越来越多的研究,旨在探讨肠道微生物在PCOS发病中的潜在作用。
相比于健康对照组,PCOS患者的粪便微生物表现出较低的微生物多样性和不同的群落谱系。健康对照组与PCOS患者的粪便微生物对比显示,PCOS患者的微生物的β-多样性显著降低,且Bacteroides vulgatus的丰度显著升高。荟萃分析显示,PCOS患者的肠道微生物在均匀性和群落谱系丰度上显著降低,相比之下,健康对照组没有显著变化;而多样性指数则普遍保持不变。进一步的分析反映了PCOS患者与健康对照组之间肠道微生物的差异性。PCOS患者中,Lachnospira和Prevotella的丰度减少,而Bacteroides、Parabacteroides、Lactobacillus、Fusobacterium和Escherichia/Shigella的丰度则升高;这些变化表现为短链脂肪酸产生菌的减少。短链脂肪酸具有抗炎作用,其减少有助于PCOS患者建立一种促炎状态。
为了进一步验证肠道微生物变化与PCOS之间的关联性,一些研究者通过来曲唑(letrozole)诱导了PCOS大鼠模型,发现Lactobacillus、Ruminococcus和Clostridium的丰度较低,而Prevotella的丰度则升高。此外,将健康大鼠的粪便移植到诱导PCOS的大鼠体内后,后者的发情周期恢复正常并表现出卵巢功能恢复的特征,包括卵巢形态的显著改善和雄激素合成的减弱。另外,仅仅移植Lactobacillus就显著改善了大鼠的发情周期,提示单一菌属可能在PCOS的发病机制中发挥重要作用。来自PCOS女性的粪便微生物或B. vulgatus移植到受体小鼠体内后,导致了PCOS的临床表型,包括胰岛素抵抗、胆汁酸代谢改变、IL-22分泌减少和不孕症,这些变化主要是通过B. vulgatus代谢物——精氨酸的作用介导的。精氨酸通过激活肠上皮细胞的FXR信号通路并以非胆汁酸依赖的方式抑制GLP-1的分泌。这些研究揭示了“肠道-卵巢轴”在PCOS发病机制中的关键作用并强调了肠道微生物,特别是单一菌属,在PCOS发展的重要性。因此,针对肠道微生物失调的干预,可能成为改善症状和阻止PCOS进展的治疗新靶点。
肠道微生物与早发型卵巢功能不全(POI)
卵巢衰老通常被认为是卵巢功能随着年龄的增长而逐渐下降的过程,其特征是卵母细胞数量减少和质量下降,伴随不规则月经、不孕症和最终的停经。更年期是卵巢自然衰老的标志,通常发生在49至52岁之间。与自然更年期不同,部分女性在40岁之前,卵巢功能便显著下降,这种情况被称为POI。POI的主要临床特征是月经异常(闭经或月经稀少/频繁)、促性腺激素水平升高(促卵泡激素 [FSH] > 25 U/L)以及40岁之前的雌激素水平下降。POI的全球患病率约为3.7%,其中,30岁以下女性的患病率为0.1%。由于POI患者不仅面临严重的生育障碍,还显著增加了心血管疾病、认知障碍、骨质疏松症和预期寿命减少的风险,引起了越来越多的关注。POI也是一种高度异质的疾病,具有复杂的发病机制。越来越多的研究发现,肠道微生物-卵巢轴在卵巢功能中起着重要作用,肠道微生物失调可能在POI的发生和进展中发挥关键作用。
与健康对照组相比,POI女性中的短链脂肪酸产生菌,如Blautia、Clostridium、Faecalibacterium、Roseburia和Ruminococcus的丰度显著下降。短链脂肪酸通过调节免疫细胞发挥抗炎和免疫调节功能,而短链脂肪酸产生菌的减少,可能通过影响短链脂肪酸的合成来影响POI的进展。激素替代疗法(HRT)是POI患者常用且有效的临床治疗方法。HRT治疗逆转了POI患者的菌群β-多样性,Eggerthella的丰度也发生了变化,表明性激素在POI患者肠道微生物组成中起着关键作用。这些结果表明,肠道微生物失调和微生物代谢产物可能是POI的核心因素;然而,具体的功能和作用机制仍需要进一步研究。
肠道微生物与子宫内膜异位症
子宫内膜异位症影响大约10%的育龄女性,定义为子宫内膜腺体和间质组织出现在子宫腔外。子宫内膜异位症的主要临床表现为慢性盆腔痛、痛经和不孕症。子宫内膜异位症的发病机制复杂,涉及遗传、免疫和环境因素;然而,子宫内膜异位症的确切病因尚未完全阐明。由于肠道微生物在雌激素代谢、炎症和免疫功能中起着关键作用,肠道微生物在子宫内膜异位症的发生和进展中的作用已引起了近期的关注。早期研究发现,与健康对照组相比,III期或IV期子宫内膜异位症患者的Shigella/Escherichia丰度显著升高;然而,其他研究者则未在月经周期的增生期和分泌期观察到子宫内膜异位症患者和健康对照组之间的肠道微生物的差异。与健康对照组相比,子宫内膜异位症患者的粪便样本中的α-多样性和β-多样性显著降低。在子宫内膜异位症患者中,来自Bacteroidia的两种细菌属(Bacteroides和Parabacteroides)以及来自Clostridia的两种细菌属(Oscillospira和Coprococcus)的丰度较高;而来自Bacteroidia的两种细菌属(Paraprevotella和一种尚未鉴定的细菌)以及来自Clostridia的两种细菌属(Lachnospira和一种尚未鉴定的细菌)的丰度则在子宫内膜异位症患者的粪便样本中较低。然而,仍需更大规模的研究来验证子宫内膜异位症患者与健康对照组之间肠道微生物的确切差异。
此外,与对照组相比,子宫内膜异位症小鼠的粪便样本显示肠道微生物的多样性和丰度减少。进一步研究发现,子宫内膜异位症小鼠的粪便样本中Firmicutes/Bacteroidetes比例升高。肠道微生物的失调促进了子宫内膜异位症小鼠病灶的生长,且粪便中的SCFAs水平(特别是丁酸)显著降低,而补充丁酸显著减小了体内异位灶的大小。这些结果表明,肠道微生物失衡和微生物产生的SCFAs在子宫内膜异位症的发展中发挥着重要作用。然而,肠道微生物与子宫内膜异位症发生和发展之间的因果关系,在人类中仍不明确,肠道代谢产物失衡在子宫内膜异位症发展中的具体作用需要进一步分析。
妊娠与肠道微生物失调
妊娠是女性生命中的一个重要生殖阶段,伴有激素水平、免疫状态和新陈代谢的显著生理变化,以支持胎儿发育。妊娠相关的并发症,如妊娠糖尿病(GDM)和先兆子痫(PE),是母婴健康的重大风险。在怀孕期间,肠道微生物(包括细菌和真菌)发生显著变化,主要受到孕妇生理过程调节的影响,特别是激素的变化。肠道微生物的变化有助于调节孕妇的内分泌、免疫反应和代谢活动,支持怀孕。孕妇的肠道微生物失调会促进多种妊娠相关并发症的发生和发展,如GDM和PE。GDM通常定义为在怀孕期间诊断或识别出的异常葡萄糖耐量,但未达到显性糖尿病的诊断标准。尽管GDM被视为怀孕期间的暂时性高血糖,但它与母婴的围产期和长期健康风险密切相关。越来越多的证据表明,GDM患者的肠道微生物发生显著紊乱。尽管关于GDM患者肠道微生物失调的研究结果仍有差异,但普遍存在Enterobacteriaceae、Desulfovibrio spp.、Aspergillus farinaceus、Ruminococcaceae、Prevotella spp.和Collinsella spp.的丰度增加,而Alistipes、E. faecalis和Bifidobacterium spp.的丰度减少。此外,肠道微生物失调通过减少微生物产生的SCFAs促进GDM的发生。PE是另一种妊娠特有的疾病,定义为妊娠20周后出现的新发高血压并伴随至少一种并发症,如蛋白尿或母体器官功能障碍,这是威胁母婴健康的严重并发症。大量研究已证实PE患者的肠道微生物与健康孕妇相比存在异常。我们的研究发现,PE患者肠道微生物的α-多样性显著降低,同时伴随着致病菌的显著升高,如Clostridium difficile(以及一些有益菌群,如Clostridium butyricum)、Dialister、Veillonella和Fusobacterium;同时,益生菌属(如Lachnospira、Akkermansia和Faecalibacterium)的丰度显著减少。这些微生物的变化与PE的临床标志物,如血压和肝肾功能的生物标志物,密切相关。PE患者的肠道微生物在怀孕大鼠中诱发了类似先兆子痫的症状,与肠道屏障功能受损和Th17/Treg比例失调相关,导致全身炎症增加和胎盘着床受损。PE患者的肠道微生物特异性地缺失了短链脂肪酸产生菌,导致胎盘中丙酸和丁酸的局部水平下降,而通过补充A. muciniphila、丙酸或丁酸,可以逆转小鼠的PE样症状,促进胎盘巨噬细胞自噬并抑制M1表型转化。此外,补充的A. muciniphila通过分泌外膜囊泡(OMVs)显著改善了先兆子痫小鼠的临床表型,提示OMVs是通过与胎盘靶细胞相互作用,启动和促进PE发展的关键因素。父方肠道微生物的失调通过干扰睾丸代谢,增加了后代的低出生体重、严重生长受限和早产死亡的概率;这影响了精子中的小RNA组成,最终导致胎盘发育受损。研究强调了肠道微生物对成功妊娠的重要性并进一步指出了父方肠道微生物失调对胎盘功能和妊娠结局的影响;这些指标对环境因素敏感,需要进一步的研究。
肠-内分泌轴
概述
肠道-内分泌轴涉及肠道微生物、肠内分泌细胞和内分泌器官之间的复杂相互作用,在人体健康和疾病中发挥着不可或缺的作用。肠道微生物产生多种代谢产物,如SCFAs、胆汁酸代谢物和吲哚衍生物,这些产物通过血液循环影响全身内分泌器官。肠内分泌细胞是肠上皮的一部分,分布在整个消化道中。它们分泌多种激素,如胰高血糖素样肽-1(GLP-1)、多肽YY(PYY)和胃抑制肽(GIP),这些激素可调节食欲、胰岛素分泌和胃肠运动,它们的功能受肠道微生物的影响。迷走神经连接着肠道和大脑,将肠道状态的信息传递给中枢神经系统,从而影响内分泌系统的功能。被称为“第二大脑”的肠神经系统独立于中枢神经系统调节肠道功能并通过复杂的神经网络与中枢神经系统进行沟通。肠道微生物和肠内分泌细胞分泌的激素和代谢产物影响下丘脑-垂体-肾上腺(HPA)轴,调节压力反应和皮质醇分泌。肠道微生物通过代谢产物和神经递质还可能影响下丘脑-垂体-甲状腺(HPT)轴,调节甲状腺激素分泌。此外,肠道微生物和肠内分泌细胞分泌的激素和代谢产物还会影响下丘脑-垂体-性腺(HPG)轴,调节性激素分泌。因此,肠道-内分泌轴是一个复杂的生物学网络,涉及肠道微生物、肠内分泌细胞、神经系统和内分泌器官之间的相互作用,对各种内分泌疾病的发展产生重大影响。下一节将具体阐明肠道微生物对糖尿病发展的影响。
糖尿病
许多研究表明,2型糖尿病(T2D)患者与健康个体之间的肠道微生物的组成存在显著差异。与健康个体相比,T2D患者的丁酸盐生成菌(如Roseburia intestinals、F. prausnitzii和Eubacterium rectale)的丰度减少,而机会性病原菌(如Bacteroides caccae、Clostridiales、E. coli)以及还原硫酸盐的细菌(如Desulfovibrio)的丰度增加。丁酸盐生成菌的减少也是糖尿病前期个体的肠道微生物的一个特征。功能上,T2D患者的肠道微生物表现出与糖运输、支链氨基酸(BCAA)运输、硫酸盐还原和氧化应激反应相关的基因增加,而与丁酸盐生物合成相关的基因减少。T2D患者肠道微生物的代谢特征也与健康个体不同,包括SCFAs水平降低、胆汁酸组成和浓度变化以及BCAA水平升高。此外,T2D患者的肠道微生物成的代谢产物,如咪唑丙酸、苯酚类化合物(如3-苯基丙酸)和3-吲哚乳酸,其浓度在T2D患者中高于健康个体。
肠道微生物影响T2D发展的主要机制包括其对能量吸收和能量平衡的影响。肠道微生物通过调节宿主食欲激素的分泌,影响能量摄入。SCFAs是肠道微生物发酵膳食纤维的主要终产物,作用于肠上皮细胞上的GPR41/43受体,促进GLP-1和PYY的分泌。这些激素抑制食欲并增加饱腹感。丁酸盐通过减少下丘脑中促进食欲的神经元的活动,降低食物摄入量。次级胆汁酸和肠道细菌分泌的某些蛋白质,如E. coli分泌的ClpB蛋白和A. muciniphila分泌的P9蛋白,也能刺激GLP-1的分泌。相反,一些代谢产物,如脱氧胆酸和高浓度的乙酸,会抑制GLP-1的分泌或促进胃饥饿素(ghrelin)的分泌,从而导致食欲增加。吲哚是色氨酸代谢物,短期内促进GLP-1的分泌,但随后抑制其分泌。肠道微生物还调节宿主脂肪组织和肝脏的产热活动,促进能量消耗。小鼠口服丁酸盐可促进棕色脂肪组织中与产热相关基因、PPAR-γ共激活因子1α(PGC-1α)和解偶联蛋白1(UCP1)的表达,增强能量消耗和脂肪氧化。乙酸上调肝脏中与脂肪酸氧化和产热相关的基因的表达,抑制脂肪在脂肪组织和肝脏中的积累。琥珀酸和P9蛋白也能促进棕色脂肪组织中的UCP依赖性产热。胆石酸通过与脂肪组织中的TGR5受体结合,促进白色脂肪和棕色脂肪的褐变,刺激产热。另一方面,这一过程调节了糖原的合成和分解,从而影响宿主的能量代谢。肠道葡萄糖异生的激活可减少与葡萄糖稳态相关的肝脏葡萄糖的生成。丁酸盐、丙酸盐和琥珀酸激活了与肠道葡萄糖异生相关的基因的表达。相比之下,硫化氢能刺激啮齿类动物的肝细胞中的糖异生和糖原分解,减少葡萄糖的利用,降低糖原储存,破坏葡萄糖稳态。
此外,肠道微生物及其代谢产物还影响胰岛素分泌和胰岛素敏感性。SCFAs通过影响胰腺β细胞的功能和胰岛素分泌来调节葡萄糖代谢。SCFAs促进GLP-1的分泌,GLP-1结合到胰腺β细胞表面的受体后,可以增强胰岛素的合成和分泌,促进β细胞的生长和增殖。另一方面,SCFAs直接作用于β细胞表面的GPR41/43受体,增强葡萄糖刺激的胰岛素分泌并改善β细胞功能。丙酸盐主要通过抑制β细胞的凋亡来确保β细胞质量和葡萄糖刺激的胰岛素分泌。然而,丙酸盐也可能对宿主的葡萄糖代谢产生不利影响,例如增加餐后血浆胰高血糖素、脂肪酸结合蛋白和去甲肾上腺素水平,这可能导致胰岛素抵抗和代偿性高胰岛素血症,从而增加T2D的风险。肠道微生物介导的蛋白质和氨基酸代谢产物显著影响胰岛素敏感性和β细胞功能。例如,血浆中BCAAs水平升高与胰岛素抵抗和T2D风险增加相关。在胰岛素抵抗患者的肠道微生物中,由Prevotella copri和Bacteroides vulgatus驱动的BCAA合成能力增强,而由Butyrivibrio crossotus和Eubacterium siraeum驱动的BCAA摄取和降解能力则减弱。肠道微生物产生的代谢产物咪唑丙酸盐(组氨酸代谢的产物)通过激活mTORC1信号通路,损害葡萄糖耐受性并干扰胰岛素信号。相反,肠道微生物代谢色氨酸的中间产物3-吲哚丙酸,与胰岛素分泌和敏感性的改善,以及T2D风险的降低相关。此外,肠道微生物发酵蛋白质的产物4-甲基酚,能刺激胰岛素分泌和β细胞增殖。
肠道微生物在肠道屏障功能受损和慢性炎症中的作用显著,这些因素对糖尿病的发展至关重要。肠道屏障的损伤,使得肠腔中的细菌和毒素进入血液,触发局部和全身的炎症反应及胰岛素抵抗,从而导致葡萄糖耐受性受损。粘液层、紧密连接和肠上皮细胞构成了肠道机械屏障的结构基础,而这些都受到肠道微生物的影响。肠道微生物及其代谢产物还调节肠道粘蛋白的合成和分泌。丁酸盐促进杯状细胞(分泌粘蛋白的细胞)的增殖以及粘蛋白-2基因的表达,增加粘蛋白-2的分泌并改善肠道粘液屏障。同样地,A. muciniphila对肠道屏障的有益作用与其促进杯状细胞增殖的能力有关。肠道微生物还调节肠上皮细胞中紧密连接蛋白基因的表达。丁酸盐和吲哚通过上调肠上皮细胞中紧密连接蛋白基因的表达,增强肠道屏障功能。A. muciniphila的膜蛋白Amuc_1100作用于TLR2,调节肠上皮细胞中紧密连接蛋白相关基因的表达,改善肠道屏障。然而,肠道微生物失调会导致有害代谢产物的增加,这些产物(如乙醇胺和反式脂肪酸)对肠道屏障功能不利,它们下调紧密连接蛋白相关基因的表达并破坏肠道屏障。肠道微生物还调节肠上皮细胞的数量。A. muciniphila及其膜蛋白Amuc_1100促进肠上皮细胞的再生和修复,从而维持肠道屏障的完整性。肠道微生物失调和肠道屏障功能受损可能导致细菌或其代谢产物进入宿主循环系统,这是诱发宿主低度慢性炎症的关键机制。革兰氏阴性细菌的细胞壁成分——脂多糖(LPS),一旦进入血液,就会激活先天免疫细胞表面的CD14/TLR4信号复合物,诱导促炎细胞因子的分泌并促进炎症反应。相反,丁酸盐促进抗炎Treg细胞的分化,抑制肠道和外周组织中的炎症反应并可以直接作用于组蛋白去乙酰化酶来缓解肠道炎症。T2D患者存在的低度慢性炎症也可能与其肠道中抗炎细菌(如F. prausnitzii)的减少有关。F. prausnitzii通过阻断NF-κB的激活和IL-8的产生来减轻炎症反应。
干预肠道微生物以缓解糖尿病
直接改变肠道微生物的结构:将健康瘦体重个体的肠道微生物移植到肥胖和代谢综合症患者体内,可以有效改善受体的胰岛素敏感性。最近的研究还发现,粪菌移植(FMT)对糖尿病周围神经病变具有一定的治疗效果。此外,补充一种或多种益生菌的混合物可以增强胰岛素敏感性,减少T2D或肥胖患者的胰岛素抵抗。临床研究进一步表明,益生菌与益生元或药物的联合使用,可能对改善葡萄糖代谢有更好的效果。
间接调节肠道微生物的结构:多个临床试验已证实,高膳食纤维的饮食干预,可以显著改善肥胖和T2D患者的葡萄糖和脂质代谢,减少肠道通透性、全身炎症和胰岛素抵抗。这一效果是通过选择性地丰富肠道中的有益菌群,如SCFA生成菌,从而增加肠道中SCFA的含量并促进GLP-1和PYY等肠道激素的分泌而实现。类似地,富含膳食纤维的Ma-Pi 2饮食,通过丰富SCFA生成菌并抑制肠道中的促炎细菌,能够有效改善T2D患者的葡萄糖和脂质代谢。
本领域挑战
从健康状态到糖尿病前期,再到2型糖尿病的发病,以及各种并发症的发展,是一个涉及代谢、免疫和神经系统的复杂多脏器疾病过程。由于先前的研究方法和策略在微生物研究中的局限性,导致关于肠道微生物与糖尿病的研究仍处于早期阶段。肠道微生物是一个复杂的生态群落,其成员并非无序或孤立存在;相反,它们通过合作和竞争等生态关系所形成的网络系统发挥作用。更重要的是,肠道微生物中的各个物种的功能并不等同。关键物种通过与宿主的相互作用而影响宿主的健康和疾病状态,而且,它们通过调节生态网络中的其他物种,能够决定群落的稳定性。这些关键物种构成了肠道微生物的核心物种。尽管付出了大量努力,但当前仍然面临着阐述核心微生物物种并理解其与宿主相互作用机制的关键科学挑战。
在早期的研究中,肠道微生物与疾病之间的关系主要是相关性,但当前的研究正朝着因果关系和机制性研究方向发展。肠道微生物及其代谢产物被肠道识别和感知,参与调节肠道上皮屏障并改变先天性和适应性免疫信号/细胞功能,可能通过远端器官介导的系统性炎症而产生影响。肠道微生物能够调节肠脑肽,如胆囊收缩素、饥饿素、PYY和GLP-1,这些肽随后可能影响大脑中的神经功能以及肠脑轴的生物反馈系统,从而影响能量稳态。因此,肠道微生物通过与宿主免疫和神经系统的相互作用,影响糖尿病的发生和进展。然而,这些微生物相互作用中的免疫系统效应分子、信号通路以及神经系统功能神经元仍不清楚。鉴于糖尿病是一种多脏器、多阶段的复杂性疾病,阐明关键微生物与宿主相互作用的多维度、多层次的分子机制,仍然是一个重要的挑战。
肠-脑轴
概述
肠道微生物可以生成各种代谢产物,通过神经、免疫和代谢途径与宿主相互作用,影响大脑功能并维持全身稳态。这种双向沟通被称为微生物-肠-大脑轴,它代表了一个紧密相连且复杂的网络,调节代谢、免疫稳态和中枢神经系统功能。许多综述系统地总结了肠-脑轴在抑郁症、帕金森病和阿尔兹海默病中的研究进展。本综述将主要集中于其在母婴微生物传递异常引起的神经发育障碍方面的研究进展。
作为一个重要的生理过程,母婴垂直传播表明,母体微生物可能通过干扰肠-大脑信号传导,以许多出乎意料且引人入胜的方式,发挥着对后代神经发育的关键作用。删除和选择性重建母体肠道微生物,可以影响后代的神经发育。无菌和抗生素处理后的母鼠胚胎大脑显示,与轴突形成相关的基因表达减少,丘脑皮层轴突数量不足,丘脑轴突生长受损。相比之下,以拟杆菌为优势定植菌的母鼠,可以提高胎儿大脑中三甲胺氧化物和咪唑丙酸的水平,防止胎儿丘脑皮层轴突形成缺陷,这表明结构和功能正常的母体微生物对胎儿神经发育至关重要。
大量证据表明,众多复杂的母体和产后因素可以引发后代大脑发育中的结构和功能异常,其影响可能延续到成年。其中一些效应甚至可能通过代际遗传跨越种系传递。前瞻性研究将妊娠期间的母体感染与后代精神障碍的风险增加,联系在一起。孕期的高心理压力与儿童行为问题和精神障碍的风险增加相关。此外,母体的代谢状态、饮食习惯和哺乳期间的行为也会影响后代的神经发育,可能改变他们对焦虑、抑郁和认知障碍等疾病的易感性。尽管存在这些关联,但其大部分潜在机制仍然不明。本文将对现有的关于母体肠道微生物对后代神经发育影响的研究报告进行批判性审视。我们的目标是提供一个全面的框架以探讨母婴微生物相互作用及其对神经发育的影响机制,从而为这一快速发展的领域提供多维度和更深入的视角(图11)。
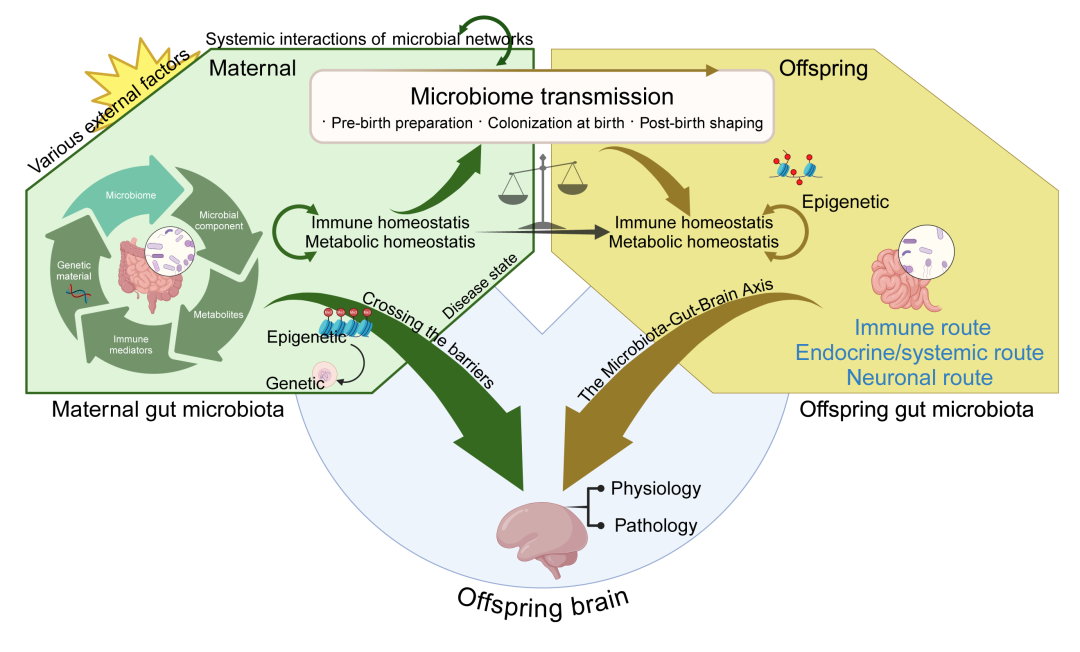
图11. 母体与后代的肠道与大脑的相互作用通路
本图概述了母体肠道微生物及其产生的成分对一系列生理过程的多因素影响,包括代谢、免疫功能、胚胎发育和哺乳。母体微生物及其生成的代谢物在产前和产后期间,精细调控后代的神经发育。这些效应可能独立于或与母婴微生物传播过程共同发生,最终塑造后代的肠道微生物。本图由BioRender.com制作。
微生物生长与大脑发育的变化时期
生命早期是微生物成熟和大脑发育动态变化最大的时期,微生物的定植和繁殖与神经发育同时进行,这两者都发生在一个对外部影响高度敏感的关键窗口期内。在这个关键时期,神经元组织快速变化,包括但不限于神经生成、轴突和树突的生长、重要连接的精炼以及髓鞘的生成。这些过程形成了身体的功能性神经回路,对于正常的认知、运动和情感发展至关重要。与此同时,肠道微生物从不稳定状态开始渐变,这种状态的特点是,从松散的微生态结构和较低的成熟度,过渡到一个更为稳定的组成,后者具有复杂的三维功能路径。大量研究已表明微生物-肠-大脑轴沟通,在结构和功能变化中的关键作用,因此,在这一时期改变或干扰肠道微生物的初始定植或发育,很可能会深刻影响肠-大脑信号传导。因此,改变的微生物可能增加神经发育障碍的风险并影响终生健康。所有这些理论强调了微生物在早期发育中的重要性,依据来自于无菌小鼠的研究中,显示了微生物在早期神经发育中的重要作用。
母体肠道微生物的系统性相互作用,在后代肠道微生物发育中,发挥着不可或缺的作用
许多因素可以影响胎儿早期微生物的定植和组成,包括遗传因素、周围环境、妊娠期间的母体饮食、感染暴露以及分娩方式。值得注意的是,来自人体多个部位的微生物——如肠道、口腔、阴道、子宫和皮肤的微生物——在结构组成和功能特性上,可能有所重叠。早期生命阶段的屏障功能逐步发展,可能为细菌的转位和相互作用提供机会。例如,口腔微生物通过肠道迁移、血行途径和免疫细胞迁移等机制,可能影响肠道微生物的组成(图12)。关于特应性皮炎的研究表明,补体系统、神经内分泌调节和AHR介导的免疫信号可能会影响肠-皮肤轴,进而影响疾病的进展和宿主的平衡(图12)。测序研究显示,早期婴儿粪便中的微生物来自各种母体来源。这些多样化微生物生态系统的集体贡献,可能突显了母婴微生物传递在关键窗口期中的整体性特征,以及它对后代健康和发育的潜在长期影响(图12)。
产前传递和预处理:母体肠道微生物在塑造后代肠道微环境和生理中的作用
目前,由于技术限制,关于微生物是否存在于产前胚胎肠道中的讨论仍存在争议,尚不清楚母体肠道微生物是否能够跨越胎盘屏障,直接定植后代肠道(图13)。在妊娠妇女的胎盘和子宫中发现微生物,挑战了胎儿环境无菌的观念(图12A)。一些研究人员在胎盘和羊水中检测到常见的口腔共生菌和牙周病原菌。女性上、下生殖道的微生态环境显示出连续的微生物分布,子宫内含有来自阴道和输卵管的微生物混合物。在这一环境中,Pseudomonas aeruginosa和Serratia marcescens是主要的微生物,这表明阴道微生物可能通过向上迁移的方式进入子宫(图12B)。与此同时,粘膜组织部位观察到高度的细菌同源性,暗示当肠道粘膜上皮屏障未成熟或受到破坏时,肠道微生物可能通过独特机制侵入子宫内膜。这些发现表明,子宫、口腔、阴道和肠道微生物之间存在关联,表明在出生前,新生儿与母体肠道微生物之间存在紧密的相互作用。这暗示了母体微生物对胎儿在子宫内健康的影响。尽管仍存在不确定性,但显然母体微生物可能通过多种机制,从产前便开始对后代微生物的定植产生影响。众所周知,胎儿经历了从受保护的子宫环境到外部世界的重大环境转变。母体微生物在这一关键生命阶段,能够为新生儿提供长期支持,帮助其应对这一挑战。分离的母婴通过脐带和母乳保持着特殊且亲密的联系,作为母体微生物影响后代发育的桥梁。在子宫内,受微生物影响的母体免疫细胞、IgG、少量的IgE、微生物抗原结构和其他免疫调节因子,通过胎盘传递,它们在胎儿体内发挥效应功能,如中和、吞噬和促进免疫细胞的激活,为胎儿在出生时及出生后进行微生物定植,准备合适的免疫和细胞微环境(图13),并对免疫系统的发育和后代健康产生影响(图14)。例如,选择性胎盘转移母体IgG为新生儿提供被动免疫保护。有研究表明,母体HIV感染可能通过影响母体IgG与胎盘表达的Fc受体FcγRIIa和FcγRIIIa的结合能力,降低病原特异性IgG的胎盘转运效率,从而减少病原的胎盘转运效率。因此,母体肠道微生物可能通过多种方式调节转移到后代的抗体的特征和数量。除了抗体转移外,母体肠道微生物还调节多种免疫细胞、细胞因子和补体成分,这些成分会影响妊娠结局和胎儿发育。例如,在生理条件下,母体肠道微生物诱导Th17分化,促进滋养层细胞在胎儿胎粪中的插入和生长,从而支持后代的健康发育。在妊娠期间消除或改变母体肠道微生物会重新校准后代免疫细胞的分布和功能,增加疾病的发生概率。值得注意的是,母体在孕期暴露于压力、饮食变化和感染等因素,所导致的微生物结构改变,产生过量的免疫产物或免疫调节代谢物,这些物质可能直接或间接介导胎儿大脑损伤。在这些事件中,母体病毒感染与后代神经发育之间的联系已有悠久的历史。例如,根据某些病毒(如风疹、流感、麻疹、腮腺炎、水痘和小儿麻痹症)的生态学和流行病学研究,科学家们已经基本绘制出它们与自闭症和精神分裂症等神经发育障碍流行的关联。使用多核苷酸-多胞苷酸(poly I:C)模拟动物妊娠相关病毒感染的后续研究表明,后代表现出类似人类神经疾病的大脑和行为特征。孕期感染后,IL-17a信号通路在肠道中过度激活,导致胎儿皮层发育畸形并影响原始CD4+ T细胞的染色质,进而导致胎儿神经发育障碍和严重的肠道炎症。此外,母体免疫激活产生的其他信号,影响后代的纹状体、海马体和皮质体积,改变大脑区域(如海马的背腹核和前扣带皮层)的转录水平,导致围神经元基质网络的缺损。此外,母体高脂饮食导致内毒素在胎儿组织中的积累,增加巨噬细胞的Toll样受体4信号通路,诱导小胶质细胞对中缝背核(DRN)中5-HT神经元的过度吞噬,从而增加后代患神经疾病的易感性。
除了免疫信号之外,在胚胎期,各种母体微生物代谢物可以在胎盘中被检测到,通过脐带传递到胎儿(图14)。其中一些代谢物直接到达胎儿大脑或影响轴突形成和神经元连接,而另一些代谢物,如SCFAs,可以到达交感神经系统、肠道和胰腺等其他部位。这些代谢物可能被游离脂肪酸受体GPR41和GPR43捕获,促进神经细胞、肠内分泌细胞和胰腺β细胞的发育,从而影响后代的长期能量代谢平衡和神经发育。因此,妊娠期肠道微生物的动态与宿主血清中的代谢物呈相关关系,相关的属水平的真菌与宿主之间的相互作用可能共同介导不良妊娠结局,如妊娠糖尿病(GDM)和胎儿过度生长。此外,母体高脂饮食引起的肠道微生物变化,提高了母体血浆中犬尿氨酸(kynurenine)的水平并调节胚胎大脑中参与谷氨酸-谷氨酰胺循环的分子水平,损害后代行为。此外,母体感染或孕期微生物结构的变化,可能导致有害代谢物、细菌产物、DNA和致病因子在胎儿环境中逐渐积累。这种积累可能影响胚胎的代谢物水平和必需营养素的生物利用度,从而可能对脆弱的胎儿大脑造成直接损伤。随着母胎界面检测方法的进步,将更好地追踪从早期生命开始的代谢物,推动这一研究领域的发展并解码细菌代谢机制。母体肠道微生物及其代谢物与母体免疫转移,共同为胎儿肠道微生物和生理状态做准备,加速早期微生物定植过程中的胎儿免疫系统的成熟,显著影响后代的神经发育。
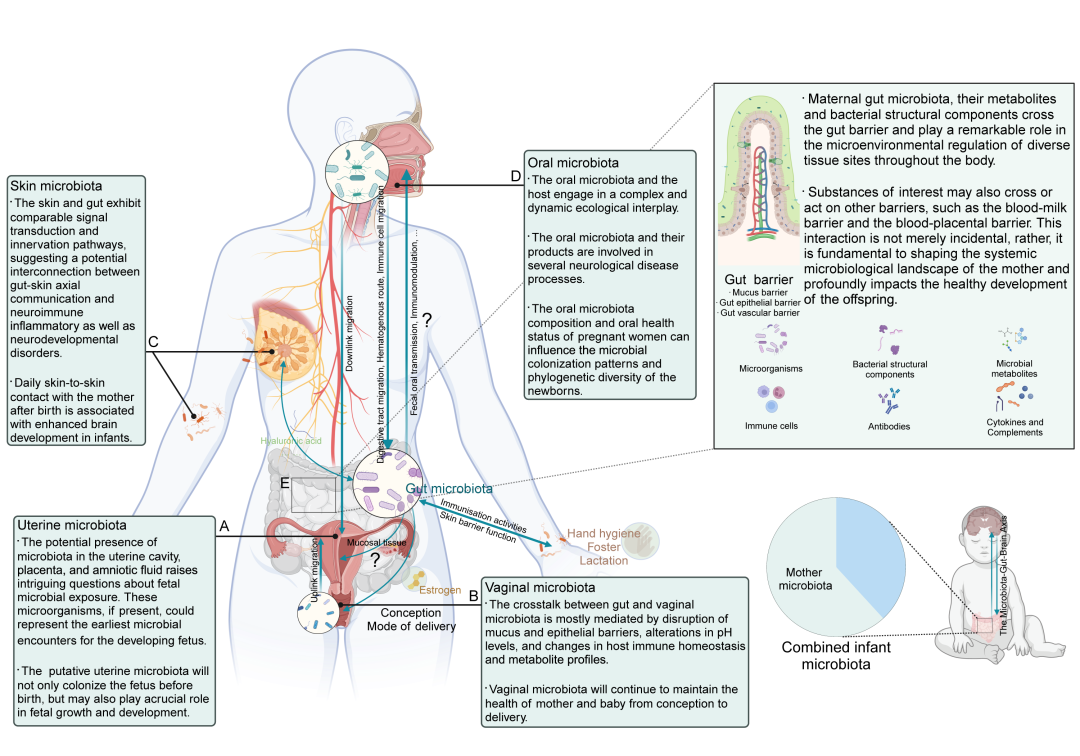
图12. 母体肠道微生物的系统性相互作用影响后代神经发育的机制
人类多个部位(包括肠道、口腔、阴道、子宫和皮肤)之间的微生物表现出显著相关性。这些多样化的微生物群落通常表现出重叠的结构组成和功能特性。母体微生物的这一复杂网络,在生理和病理过程中进行复杂的相互作用和协同作用,可能对后代的神经发育轨迹产生深远影响。此外,母体体内的这些不同部位,在促进微生物从母体到婴儿的垂直传递中,发挥着关键作用。这一微生物转移过程并不限于单一部位,而是涉及多个微生物群的协调作用,如子宫微生物群(A)、阴道微生物群(B)、皮肤微生物群(C)、口腔微生物群(D)以及身体的主要屏障(E),特别是肠道屏障,它是微生物系统性相互作用中的重要环节。本图由BioRender.com制作。
出生时肠道微生物的定植
胎儿肠道是否在出生前就被母体微生物定植,尽管仍存在争议,但大量证据表明,母体微生物通过多个途径在出生时及出生之后,垂直传递给婴儿。母体微生物与婴儿肠道微生物之间的频繁互动和共享,发生于生命早期,母体对后代的贡献最为显著。不同的分娩方式改变了胎儿初始微生物的暴露环境,从而导致不同的定植效应(图13)。通过剖宫产分娩的婴儿表现出截然不同的微生物组成和发育轨迹,因为他们主要暴露于医院环境中的微生物;而经阴道分娩的婴儿主要由母体生殖道的细菌定植。值得注意的是,肠道微生物可能参与这一过程,通过代谢关键宿主因子(如雌激素)来调节阴道微环境和功能。剖宫产分娩通常与肠道微生物的发育受限相关,例如在胎粪中Lactobacillus spp.的丰度较低,且易感病原菌的定植率较高,这对新生儿健康有显著影响,而且可能持续到成年。然而,剖宫产分娩引起的胎儿肠道微生物结构的显著改变,可以通过出生后的母体粪便移植进行有效纠正。这进一步支持了母体肠道微生物在发育过程中,对后代肠道微生物塑造的作用。
此外,在生命的早期阶段,母体肠道微生物在婴儿肠道中的持续性和生态适应性,在随后的母乳喂养和日常护理过程中变得更加明显,逐渐成为婴儿发育中的肠道微生物的重要组成部分。
出生后微生物的传递与塑造
出生后,母亲的母乳喂养和养育行为接管了肠道微生物依赖的母婴交流界面,促进了婴儿的健康和长期福祉。母体微生物代谢产物(SCFAs和寡糖)与囊泡,以及肠道微生物相关的免疫物质(抗原肽、免疫球蛋白、细胞因子和补体成分)通过淋巴-血液循环到达乳腺,成为母乳的一部分(图13)。因此,母乳富含营养和生物活性因子,这些因子强化了婴儿的胃肠道并促进了大脑的健康发育。已经有越来越多的研究,使用培养和测序方法,报道了乳汁中部分源自母体肠道的微生物所涉及到的物种、属和菌株水平。这些微生物可能通过肠道-乳腺-口腔轴直接或间接地定植于后代的肠道。例如,双歧杆菌群体可能通过母乳,从母亲传递给婴儿。母婴连接中,噬菌体的作用也有相应证据。噬菌体与宿主细菌的共同进化,可能影响婴儿肠道微生物的多样性和成熟度。双歧杆菌噬菌体可能通过母体肠道,垂直传递到婴儿,参与影响婴儿肠道中双歧杆菌的组成和功能。因此,某些细菌的开创性定植,通过重写后代肠道中的代谢组学特征和免疫环境,增强或限制其他细菌的适应性。来自母体肠道微生物的代谢产物,以及锚定微生物的免疫球蛋白A(IgA),可以通过产后母乳传递给婴儿。这种传递间接地促进了胎儿特定微生物的定植和共生,支持有益肠道微生物的发育,促进免疫系统的成熟,从而保护婴儿免受侵入性感染。例如,母乳中由肠道微生物在发酵过程中形成的短链脂肪酸(乙酸和丙酸)及其他代谢产物,不仅作为交叉喂养底物,促进Bifidobacterium等细菌在后代肠道中的定植,还对Clostridiaceae和Peptostreptococcaceae等微生物发挥抗微生物活性。另一方面,母乳中高水平的花生四烯酸(AA)可能导致婴儿肠道微生物失调和免疫障碍。在婴儿肠道内,母乳中的蛋白质经过各种蛋白酶的消化,释放出数千种多肽,执行多种功能。相关研究通过对具有抑制Staphylococcus aureus生长活性的肽的验证,证明母乳蛋白能在后代肠道微生物的发育中起塑造作用。
同时,养育过程由于充满了日常接触,是母体肠道微生物向后代传播的重要部分。母体的皮肤微生物已被确定为婴儿皮肤、口腔、肠道和其他部位微生物的来源,参与建立母婴多个身体部位之间的成熟的、共生的微生物网络。荷兰的一项随机对照试验发现,与对照组相比,母婴每日皮肤接触的婴儿,在肠道微生物组成、细菌丰度以及预测的功能富集上,存在显著差异。尽管该研究不能排除母乳喂养时长等因素的影响,但结果仍然支持微生物传播和出生后变化的观点(图13)。与母亲进行每日皮肤接触,有助于婴儿的身体状况和大脑发育。三岁时,接受皮肤接触干预的儿童也表现出较少的内化和外化行为问题。这一发现,与“健康的母体肠道微生物,共同塑造后代肠道微生物特征”的观点相一致。当母亲在怀孕和母乳喂养期间暴露于感染、饮食变化和压力时,肠道微生物失调会导致大量炎症细胞因子(例如 IL-17A)或其他次级物质跨越胎盘或进入乳腺,直接到达后代体内。这可能改变物质的生物可利用性、免疫微环境以及后代初始微生物定植的过程(图13)。母体肠道微生物失调,通过内毒素血症诱导内源性乳腺炎的发展,从而导致动物宿主抗炎酶活性的降低。此外,孕期和产后母体感染可能直接为易感病原体的垂直传播创造条件(图13),进而导致新生儿败血症、脑膜炎等一系列不良后果。值得注意的是,许多病毒可能在母体感染期间,穿越胎盘并进入母乳。这些易感病原体表现出多样的致病特性,通过多种机制影响新生儿系统的发育。在“机会窗口”期间,许多细菌可能直接攻击新生儿的大脑,因为此时身体的主要屏障功能尚未完全建立。这些细菌产生的毒力因子和次级炎症介质,未经调控地渗透到大脑,导致严重的神经系统疾病,如脑炎和白质损伤。这些病理过程可能导致持久的神经系统损害和后遗症。从本质上讲,母体微生物对后代微生物定植、免疫成熟和智力发育的影响是显而易见的。在这个关键时期,母体肠道微生物群塑造了后代的早期微生物组和免疫系统,二者通过肠脑通讯,共同调节正常的神经发育。
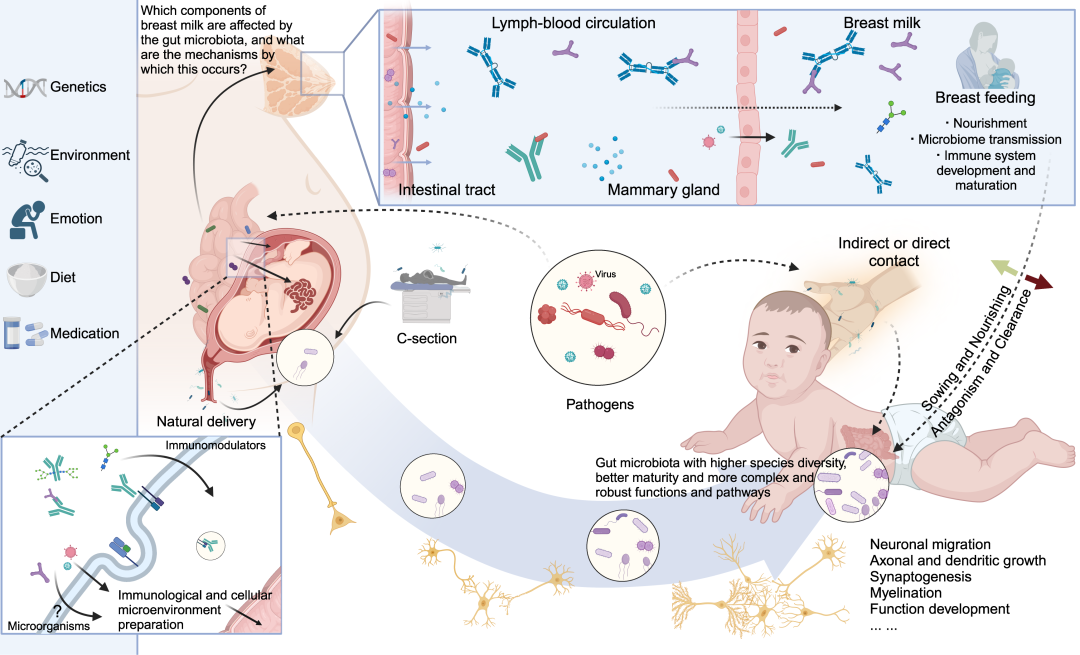
图13. 母体肠道微生物通过影响后代的肠道微生物定植调节神经发育
母体肠道微生物通过双重途径对后代的微生物定植产生深远影响。首先,它直接塑造胎儿在分娩过程中的微生物组成。此外,母体对胎儿微生物定植的影响可能从母胎界面开始,甚至在产后通过母乳喂养、抚养和母婴陪伴持续存在。母体肠道微生物及其代谢物在母胎界面的复杂交流,可能为后代提供适应性反应和微环境做准备。产后母婴接触和分娩方式决定了婴儿首次接触的微生物,从而为后代肠道微生物定植的发展奠定基调。母乳的组成受到母体肠道微生物的复杂影响,它为后代肠道微生物提供了持续的接种源。在母乳喂养和抚养过程中,后代的肠道微生物经历了成熟和稳定的过程,同时也有助于神经功能的完善。在产前和产后阶段,环境因素、饮食影响和病原暴露的复杂相互作用调节着母体肠道微生物向后代的传递过程。这一传递在后代的神经发育中起着重要作用。本图由BioRender.com制作。
母体肠道微生物通过微生物-代谢物-肠-脑轴影响后代的神经发育
微生物代谢物是微生物在物质和能量转化过程中,通过分解或合成产生的生物活性物质。几种肠道微生物代谢物在宿主的神经健康和生理过程中,发挥重要作用。例如,SCFAs从肠粘膜转移到血液循环中,干扰免疫调节和肠神经系统功能,调节神经肽的分泌并影响大脑功能。SCFAs能够穿越血脑屏障,直接与神经细胞相互作用。例如,直接激活终纹床核(BNST)中的钙/钙调蛋白依赖性蛋白激酶II(CaMKII)标记的神经元,增加它们的突触前谷氨酸释放和脂肪酸β-氧化水平,从而影响社交行为。微生物代谢物4-乙基苯酚硫酸盐也可以直接损害丘脑室旁核中的少突胶质细胞的成熟,进而改变大脑特定区域的活动和功能连接,引发焦虑。早期生命中的健康肠道微生物代谢物以适当的水平,协助身体参与多种生理过程,如促进肠蠕动和收缩,参与细胞信号转导、神经递质的合成和释放,负责建立代谢和免疫的稳态。因此,母体肠道微生物在新生儿微生物定植和发育中,显著影响早期生命中的微生物代谢过程并促进建立稳定健康的微生物-宿主互惠共生关系(图14)。相反,饮食变化和感染暴露等条件下,母体微生物的损害可能会破坏这种微妙的协同关系。简言之,母体肠道微生物与后代肠道微生物和代谢稳态的关系,可能对代谢和神经系统紊乱的发展产生影响(图14)。例如,母亲具有代谢障碍(如葡萄糖耐量受损),其后代也往往会发展出代谢紊乱、免疫功能障碍和神经发育缺陷。在患有妊娠糖尿病(GDM)的母亲组中,新生儿的肠道微生物也呈现出与母亲相似的变化趋势。与健康对照组相比,GDM组新生儿粪便代谢物中的甘油磷酸胆碱、甘醇酸和鼠李糖的丰度显著较低,而核黄素和牛磺酸的丰度显著较高。这些变化与母亲粪便中观察到的趋势一致,提示GDM母亲可能导致后代微生物分布和功能异常,这将不利于后代的代谢功能和营养吸收,进而对生长发育带来许多潜在风险。在小鼠中,母鼠高脂饮食诱导肠道微生物失调、神经递质变化、代谢功能异常,以及与大脑和神经发育、离子转运和膜电位调节相关基因的转录变化,导致神经发育缺陷。综上所述,当母体状态发生改变时,母体肠道微生物及其代谢物的组成发生显著变化,通过垂直传播到后代,重新编程婴儿的肠道微生物和代谢能力,影响外周细胞和器官(包括大脑)的代谢功能,从而改变神经发育轨迹。上述研究强调了在早期生活中,维持健康的母体微生物和代谢状态的重要性,因为这对后代的代谢健康和神经发育具有深远的影响。
母婴互动在分娩后迅速增加,伴随有许多感官、知觉和行为变化。显而易见的是,这些感官感知和母性行为的缺失,会影响后代在成长过程中的身体和心理发育。相比之下,母体感染或特定菌株的定植,会导致母体的照料行为(如舔舐、梳理、嗅探等)减少,从而影响母体对后代的哺育,进而损害后代获取必需营养物质的能力。从机制上看,鼠仔表现出丝氨酸/苏氨酸激酶Akt信号传导受损,血清中胰岛素样生长因子1(IGF1)的分泌和稳定性受损,导致后代各系统的发育延迟和受损。当然,某种程度上,目前仍不清楚肠道微生物的动态变化是否介导了母体行为的变化。此外,涉及微生物和行为的代谢物/神经递质(如血清5-HT)的研究,尤其是在接近母亲的后代行为中的作用,为探索微生物产生的神经活性化合物如何影响后代神经发育,提供了新思路和视角。
另一方面,母体在哺乳期间产生的肠道微生物代谢物可以绕过后代的系统发育,直接与脑细胞相互作用,影响胎儿大脑结构和功能的发展。有趣的是,越来越多的微生物代谢物被用于临床试验,作为复杂疾病的潜在治疗方法。母乳是婴儿最好的天然食物,包含了几乎所有婴儿生长发育所需的营养成分,包括蛋白质、脂肪、碳水化合物、维生素、激素以及与免疫系统和神经系统发育密切相关的其他生物活性物质。母乳中的长链脂肪酸、铁、胆碱、叶酸和神经鞘脂与后代许多脑区的早期髓鞘形成显著相关,为支持语言、认知和行为功能发展的脑连接提供了基础。从机制上看,人乳中的肌醇增强了神经元对诱导突触形成的跨突触相互作用的反应能力,促进了神经元的连接性。值得注意的是,人乳中的20-α-羟基胆固醇通过Gli依赖性途径,在小鼠中诱导少突胶质细胞生成,从而逆转白质损伤。然而,由于母体的地理差异、饮食习惯、健康状况、分娩时的孕期等因素,以及母乳中哪些成分受到肠道微生物影响尚不清楚,母乳成分的异质性使得相关研究面临严峻挑战。目前在这一领域的进展大多仍局限于强调母体肠道微生物代谢的膳食成分,通过母乳对婴儿的代谢、免疫调节和营养支持作用。值得注意的是,哺乳对婴儿在关键发育期的神经发育的影响,尚未得到充分探讨。为了全面了解母乳中肠道微生物代谢物的变化对婴儿在不同生命阶段和健康状态下发育路径的作用及潜在不良影响,还需要更多全面的组织学研究。
母体肠道微生物通过微生物-免疫-肠-脑轴影响后代的神经发育
出生后的早期对于免疫系统的发育至关重要。除了遗传学和宿主生物学,肠道微生物在婴儿免疫成熟和整体健康中扮演着重要且不可逆的角色。肠道微生物与局部免疫系统的相互作用,引发了功能变化,甚至超出了胃肠道,影响了系统性或中枢免疫状态并改变了神经发育轨迹。肠道微生物的成分或释放到环境中的可溶介质,激发宿主适应性免疫反应,刺激或调节免疫细胞的分化和发育,调节炎症反应,促进屏障完整性。这些物质可能涉及炎症和神经发育轴,从而影响或改变神经发育轨迹。此外,也有大量研究报告了它们直接调节大脑中的神经细胞(如小胶质细胞——主要的脑内免疫细胞)成熟和功能的作用。相反,新生儿由于免疫系统不成熟,在面对新的外部环境时,严重依赖母体来源的微生物、代谢物和免疫物质的转移,以建立机体稳态(图14B)。母体免疫力和肠道微生物在母胎界面的影响,妊娠和哺乳期间微生物的转移,以及通过哺乳转移免疫物质、微生物成分(如脂多糖、肽聚糖和DNA)和代谢物,是后代生命早期微生物和免疫训练的重要来源。在分娩过程中,后代获得的母体来源微生物,可刺激肠道上皮细胞的激活,从而获得免疫耐受性,调节自然防御和先天免疫识别。母乳中的微生物有助于婴儿肠道微生物的定植,刺激新生免疫系统中的T细胞和产IgA型B细胞的激活与分化。与此同时,某些微生物产生的代谢物,在调节后代免疫系统发育中的作用越来越被关注。在小鼠模型中,母体摄入膳食可溶性纤维,增加了母体肠道微生物代谢物对后代系统性免疫反应的影响。母乳中的人类乳寡糖和乳蛋白更多地参与了早期生命阶段中的后代肠道屏障的调节和成熟,作为底物,促进肠道细胞的分化和粘液生成,塑造肠腔内免疫微环境的轮廓。此外,乳液来源的外泌体(mev)可能通过炎症信号通路和炎症小体的激活,调节后代肠道微生物结构和肠道免疫。然而,目前仍难以全面理解这些因素影响后代神经发育的结果,因此,迫切需要深入探讨早期发育阶段中的微生物-免疫-大脑轴的沟通机制和通路。关于免疫物质的转移,许多研究阐明了母体抗体转移,在促进后代健康发育中的关键作用。例如,小鼠乳汁中的IgA影响了其与后代肠道中的共生菌的结合,这反过来通过互惠抑制关系,决定了RORγ+ Tregs的数量,进而影响肠道Treg的分化和功能以及小鼠世代的免疫调节基调。然而,关于母体微生物如何影响抗体跨越胎盘细胞屏障和通过乳腺转移的类型与效率的研究仍然有限。撇开抗体转移在早期生命免疫稳态-神经发育交换中的复杂作用不谈,目前尚不清楚,当后代发育过程中的血脑屏障尚不完全时,微生物介导的异常改变或过量的母体衍生抗体,是否会损害大脑结构和功能。这表明,虽然我们关注母婴免疫联系,但可能还需要更仔细地评估肠道微生物与自身免疫疾病之间的关联,尤其是通过检查已知可以到达或存在于后代大脑中的抗体。类似地,除了抗体转移外,母体肠道微生物还调节一些免疫细胞、细胞因子和补体成分,这些都通过母乳影响婴儿的发育。单细胞测序和流式细胞实验显示,受母体肠道微生物影响的免疫细胞(如B细胞、浆细胞和巨噬细胞)也出现在人类和小鼠的乳汁中。母乳中的补体通过C1依赖、抗体独立的机制直接裂解某些革兰阳性肠道共生菌的特定成员,导致膜攻击复合物的沉积并随之发生细菌溶解,从而保护肠道免受病原体侵害。然而,这些免疫细胞及相关免疫介质在后代早期生命中的神经发育过程中的作用,仍需进一步阐明。
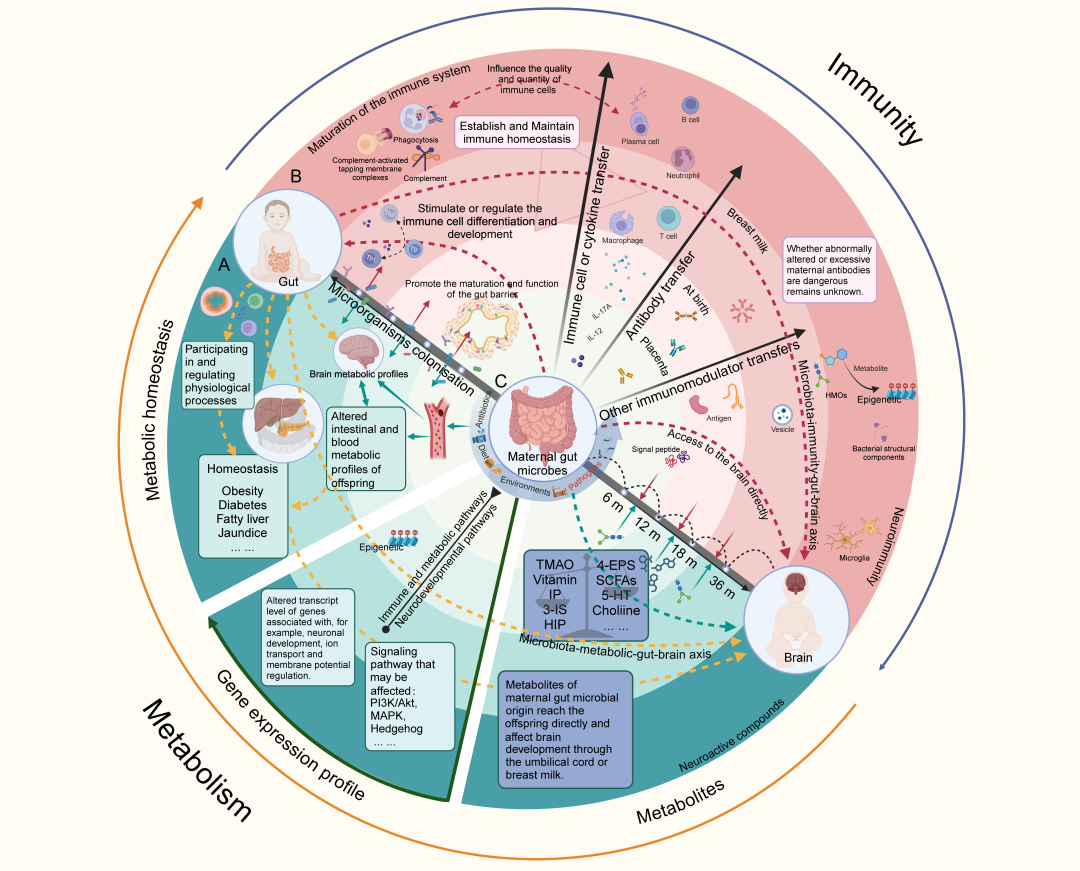
图14. 母体肠道微生物通过微生物-代谢/免疫-肠道-大脑轴影响后代神经发育
(A) 母体肠道微生物及其代谢物对后代的代谢稳态产生三重影响,促进与发育中的大脑进行直接和间接的沟通。(B) 母婴免疫转移在调节后代肠道微生物结构、引导免疫成熟和调节神经免疫健康方面具有深远影响。(C) 中轴线代表后代肠道和大脑的连续发育轨迹。这个过程受到代谢和免疫因素之间持续而强烈的相互作用的影响。这些系统的复杂相互作用强调了早期发育的动态特性及母体对后代的长期影响。本图由BioRender.com制作。
结 论
肠道微生物通过多种途径(包括免疫调节、宿主细胞死亡、代谢等)影响肠外器官(图15)。相反,肠外器官也可能在组成和功能层面上影响肠道微生物。因此,所有这些轴可能是双向的。此外,精确调控肠道微生物的那些有前景的策略,可能帮助我们在未来应对肠外器官的疾病。
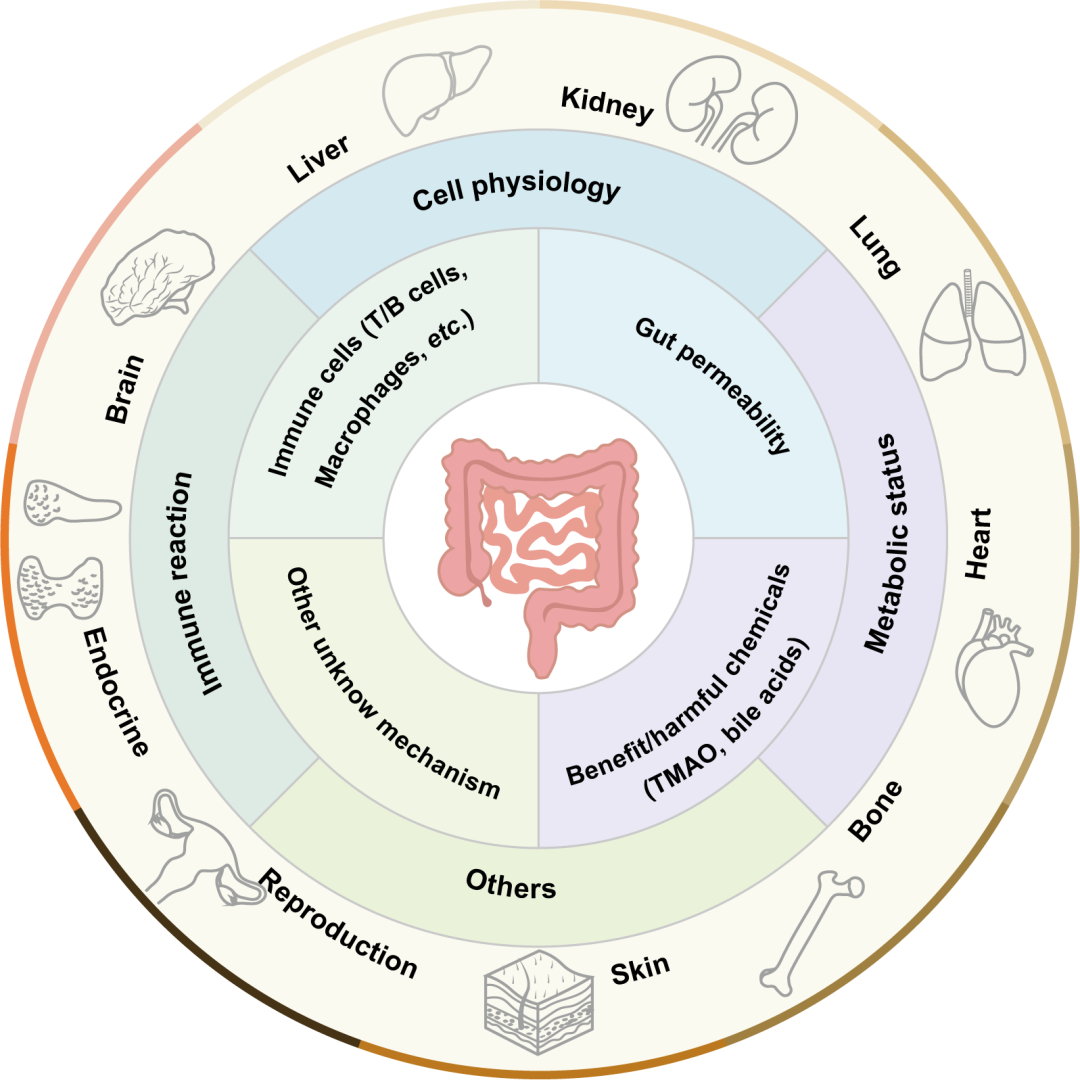
图15. “肠-X轴”的一般调节机制
肠道、肠道微生物及其产生的化学物质可能通过影响免疫细胞功能、肠道通透性以及其他未知机制,调节多个器官的病理生理,包括代谢状态、免疫反应等。
代码和数据可用性
本研究未创建或分析新数据。补充材料(文本、图、表、中文翻译版本或视频)也可从线上(http://www.imeta.science/)获取。
引文格式:
Xu Lin, Zuxiang Yu, Yang Liu, Changzhou Li, Hui Hu, Jia-Chun Hu, Mian Liu, Qin Yang, Peng Gu, Jiaxin Li, Kutty Selva Nandakumar, Gaofei Hu, Qi Zhang, Xinyu Chen, Huihui Ma, Wenye Huang, Gaofeng Wang, Yan Wang, Liping Huang, Wenjuan Wu, Ning-Ning Liu, Chenhong Zhang, Xingyin Liu, Leming Zheng, Peng Chen. “Gut-X Axis.” iMeta 4: e270. https://doi.org/10.1002/imt2.270.
作者简介
林旭(第一作者)
● 南方医科大学顺德医院医师、博士、博士后,中国营养学会注册营养师,兼任中国卫生与健康医疗大数据学会糖尿病专业委员会青年委员、广东省精准医学应用学会微生态医学分会委员、广东省药学会内分泌代谢用药专家委员会委员兼秘书。
● 研究方向为骨质疏松症与人体肠道微生态及遗传因素。以第一作者在Nat Commun.等杂志发表论文6篇,主持国家自然科学基金面上项目1项、参与省级以上课题5项。

于祖祥(第一作者)
● 北京大学基础医学院博士生。
● 主要研究方向为肠道菌群与心血管疾病,以第一作者发表sci论文一篇。

刘洋(第一作者)
● 南京医科大学一年级博士生。
● 研究方向为肠道微生物与神经发育,获南京医科大学校级一等奖学金,参与发表多篇SCI论文。

李长洲(第一作者)
● 南方医科大学南方医院整形外科博士生。
● 研究方向为肠道微生物对皮肤微生态及皮肤疾病的影响,以第一作者发表SCI论文3篇。

胡绘(第一作者)
● 医学博士,同济大学附属东方医院南院检验科微生物学与分子诊断组副组长,主持分子诊断组工作。
● 研究方向为肺癌分子标志物的机制研究和转化应用,主持国家自然科学基金青年一项,省市级课题和人才计划四项,以第一作者/通讯作者在Nature Communications,Journal of Hematology & Oncology等期刊上发表SCI论文17篇。

胡家淳(第一作者)
● 中国医学科学院/北京协和医学院药物研究所在读博士生。
● 研究方向为基于肠道菌的中药活性成分PK-PD研究。以第一作者身份发表学术论文2篇。

刘勉(第一作者)
● 妇产科学博士,博士后。
● 研究方向聚焦女性生殖健康与肠道菌群互作机制,在肠-生殖轴调控领域形成特色研究体系。近五年以第一/共一在Gut、Journal of Translational Medicine等期刊发表系列成果,揭示菌群失衡影响生殖功能的作用及其分子机制。现主持国家自然科学基金青年项目,参与国家级、省部级科研项目多项。

郑乐民(通讯作者)
● 北京大学心血管研究所副所长,博雅特聘教授,基础医学院党委委员,全国重点实验室主任助理。1998年本科毕业于北京师范大学;2005年在美国克利夫兰医院获博士学位;2005年至2007年在Cleveland Heartlab. Inc(克利夫兰心脏实验室)从事研究科学家工作;2007年至2015年在北京大学医学部任副教授;2015年至今在北京大学心血管研究所任研究员。主要从事心脑血管代谢方面的研究工作。
● 围绕“代谢产物如何影响心脑血管疾病”这个关键问题,发现了琥珀酸等多种心血管疾病新的代谢相关风险因素和TMAVA等新的活性肠道菌群代谢物及作用机制,并创新构建代谢探针与纳米探针实现心血管代谢相关在体成像以辅助诊断。共发表SCI文章131篇,其中98篇SCI责任作者,近五年成果多发表在Signal Transduction and Targeted Therapy(2024),Advanced Functional Materials(2024),Neuron(2023),Cell Metabolism(2023),Nature Communications(2022),European Heart Journal(2021),Advanced Material(2020),Gastroenterology(2020)等国际知名杂志期刊。获得国家“万人计划”科技创新领军人才(万人),国家自然基金委“优秀青年基金”,中组部“万人计划”青年拔尖人才基金,教育部“新世纪人才基金”等,并担任科技部重大专项项目负责人与北京市重点基金项目主持人等。作为第一完成人获得2022年北京医学科技奖一等奖和2022年度北京市自然科学奖二等奖。

刘星吟(通讯作者)
● 南京医科大学/南方科技大学教授,肠道微生态学术带头人;江苏特聘医学专家;江苏特聘教授;江苏省333高层次人才;承担科技部重点研发和国家自然科学基金项目多项。获华夏医学科技一等奖和教育部优秀科技成果二等奖,中国生物物理学会优秀女科学家奖。获2023年Cell Press最有影响力和最受欢迎的中国作者之一。
● 从事肠道微生物及其代谢与宿主表观遗传系统互作对神经发育障碍疾病和肿瘤发生发展的调控机制。在Cell Metabolism, Cell Host & Microbe, GUT, Cell Reports, Genome Medicine, Gut Microbes等国际知名杂志发表论文50多篇,授权专利8项,转化2项。担任中国医药教育微生态与健康专委会副主任委员、中国神经科学学会儿童认知和脑功能障碍分会委员、中国生物物理学会肠菌分会委员、担任知名微生物学国际期刊Microbiological Research 杂志Associate Editor。

张晨虹(通讯作者)
● 上海交通大学生命科学技术学院、微生物代谢国家重点实验室研究员。
● 入选第四批国家“万人计划”青年拔尖人才、国家自然科学基金“优青”、仲英青年学者、上海市教委曙光人才计划、上海市科委青年科技英才扬帆计划等。担任中国微生物学会微生物组专业委员会委员、中国生物物理学会肠道菌群分会委员、上海市微生物学会基础微生物专业委员会委员、上海市预防医学会微生态专业委员会委员等。研究方向为营养调控肠道菌群与代谢性疾病,以第一作者或通讯作者在国际上具有影响力的杂志Science、Cell、Cell Metabolism、 Nature communications、Microbiome、ISME Journal等发表论文。2022-2024连续三年入选科睿唯安 “全球高被引科学家”。

刘宁宁(通讯作者)
● 研究员/博士生导师,上海交通大学医学院单细胞组学与疾病研究中心副主任。
● 入选国家优青、上海市优秀学术带头人、上海市青年拔尖人才等。担任中国抗癌协会整合肿瘤学分会第三届委员会委员、中国医药教育协会微生态与健康专委会委员、中国菌物学会遗传与分子生物学专委会委员、上海市微生物学会临床微生物学专委会副主任委员、上海市预防医学会卫生检验专委会副主任委员、上海市医学会分子诊断专科分会青委会副主任委员等。兼任hLife、Fungal Genetics and Biology等期刊编委和Cell Host & Microbe等期刊审稿人。研究方向聚焦真菌感染与癌症发展,通过整合微生物组、单细胞组学等技术,解析癌症发展过程中的真菌与宿主相互作用的分子机制,提供抗真菌抗癌的新策略和新靶标。以通讯/第一作者在Cancer Cell, Nat Microbiol, Nat Protoc, Nat Commun (3篇), Cell Res, PNAS等杂志发表论文30余篇;获授权发明专利4项;主持国自然优青、国家重点研发计划课题、国自然面上和青年等17个项目。研究成果入选“ESI高被引论文”和上海交通大学“十大科技进展”,被同期Cancer Cell杂志前瞻评述和F1000prime高度评价,被Cell、Nat Rev Microbiol、Nat Rev Cancer、Nat Rev Clin Oncol、Nat Rev Gastro Hepat等杂志多次引用和正面评价。

吴文娟(通讯作者)
● 上海市东方医院南院检验科主任、同济大学教授/博导/博士后合作导师、中国女医师协会检验医学会分会副主任委员、中华医学会检验医学分会微生物学组委员、中国合格评定国家认可委员会主任评审员、国家卫生健康委药敏标准委员会专家委员、国家卫生健康委全国真菌病监测网专家委员、上海市微生物学会临床微生物学专业委员会主任委员、上海市医学会分子诊断专科分会副主任委员。
● 研究方向为病原微生物快速检测及耐药机制研究、医院感染控制等。上海市优秀学术技术带头人,主持国家科技重大专项课题2项、国家自然科学基金4项、省市级课题和人才计划20余项,发表论文140余篇,编写行业标准、专著10余项,授权专利6项。

黄莉萍(通讯作者)
● 南方医科大学南方医院妇产科主任,主任医师/教授,博士生导师/博士后合作导师。
● 国家健康科普专家,现任中国妇幼保健协会围产营养与代谢专委会常委、中国优生科学协会第八届理事会理事、广东省医院协会妇产科分会副主委、广东省医师协会围产医学分会副主委、广东省医师协会妇产科医师分会常委、广东省医学会围产分会常委、广东省医学会第妇产科学分会青委副主委、广东省健康管理委员会母胎医学专业委员会常委等学术职务。在南方医院从事妇产科临床医疗、教学、科研工作23年,在围产医学领域,尤其在高龄及多胎妊娠监护、妊娠期高血压、复发性流产及产科危急重症救治等方面有较深入的研究和丰富的临床经验,先后获广东省卫生系统“青年岗位能手”、广东省最强科室之“实力中青年医生”、第五届“羊城好医生”等称号。主持国家自然科学基金区域联合重点项目、面上项目等国家级项目5项,在Gut、Adv Sci、JEV等国际知名期刊发表学术论文。发现肠道菌群紊乱是子痫前期的潜在关键病因,首次提出“肠道-胎盘轴”概念,研究成果以封面论文发表于Gut,同期获JAMA专题述评,并被评为“2020年中国肠道菌群十大研究成果之一”。

王琰(通讯作者)
● 中国医学科学院药物研究所,博士,研究员,博士生导师,药物代谢研究室主任,“创新药物非临床药物代谢及PK/PD研究”北京市重点实验室主任。“十三五”中国医学科学院创新工程首席专家。
● 多年从事药物代谢及药代动力学研究。主要贡献是揭示了肠道菌与口服天然药物相互作用的本质,阐明了难吸收天然药物“为什么有效”的化学基础和生物学原理,解析了多个天然药物(小檗碱等)在肠道菌作用下的药代/药效/机理/个性化治疗的科学内涵,主编了国内第一部“肠道菌与药物代谢”专著。担任中国药理学会药物代谢专委会副主任委员,分析药理学专委会常委,天然药物药理专委会委员;中国药学会应用药理学专委会委员;北京药理学会常务理事、药物代谢专委会主任委员;北京药学会临床研究与转化副主任委员,药物化学生物学专委会委员;“药学学报”及英文刊Acta Pharmaceutica Sinica B (影响因子14.7)期刊编委,“中国药学杂志”、“药物分析杂志”、“中草药”、“沈阳药科大学学报”编委等。

王高峰(通讯作者)
● 南方医科大学南方医院整形外科教授,博导,广东省杰青。
● 研究方向为皮肤微生态对于皮肤屏障修复,创面愈合,皮肤再生的影响,正在开展Chinese Skin Microbiome (CSM) Study Group多中心多组学的皮肤微生物基础与临床研究。以第一作者或末位通讯作者在Cell Host& Microbe(2021, 2024), Science Advances(2022), JAMA Dermatology(2020), J Allergy Clinical Immunol (2024), Allergy(2023), J Invest Dermatol(2018, 2023), Small(2024), Acta Biomaterials (2023), Acta Pharmacol Sin (2023), Adv Dru Deli Rev (2023), J Dermatol(2020) 等期刊发表论文。

陈鹏(通讯作者)
● 南方医科大学教授、博士生导师。
● 研究方向为肠道菌群与器官衰竭。
更多推荐
(▼ 点击跳转)
iMeta | 引用16000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 兰大张东组:使用PhyloSuite进行分子系统发育及系统发育树的统计分析
iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVI

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

3卷2期

3卷3期

3卷4期

3卷5期

3卷6期

1卷1期

1卷2期
期刊简介
“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述,重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊!相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录!2024年6月获得首个影响因子23.8,位列全球SCI期刊前千分之五(107/21848),微生物学科2/161,仅低于Nature Reviews,学科研究类期刊全球第一,中国大陆11/514!
“iMetaOmics” 是“iMeta” 子刊,主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任,是定位IF>10的高水平综合期刊,欢迎投稿!
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science

















 650
650

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









