






点击蓝字 关注我们
肠道菌群衍生的色氨酸代谢物通过调节3型先天淋巴细胞改善全肠外营养相关感染

iMeta主页:http://www.imeta.science
研究论文
● 原文: iMeta (IF 23.8)
● 原文链接: https://onlinelibrary.wiley.com/doi/10.1002/imt2.70007
● DOI: https://doi.org/10.1002/imt2.70007
● 2025年2月26日,南京大学附属金陵医院王新颖课题组在iMeta在线发表了题为“Gut microbiota-derived tryptophan metabolites improve total parenteral nutrition-associated infections by regulating Group 3 innate lymphoid cells”的文章。
● 本研究阐明了全肠外营养(TPN)过程中的“微生物-代谢-免疫”的调控机制,为促进临床慢性肠衰竭(CIF)病人的肠康复治疗、降低感染易感性提供了重要的理论基础。
● 第一作者:黄龙昌、王鹏、刘帅、邓桂芳
● 通讯作者:王新颖(wangxinying@nju.edu.cn)
● 合作作者:齐欣、孙广明、高学金、章黎、张玉鹏、肖亚琴、高婷婷、古丽苏杜姆·麦提阿卜拉
● 主要单位:南京大学医学院附属金陵医院,佛山第一人民医院,华中科技大学协和深圳医院
亮 点
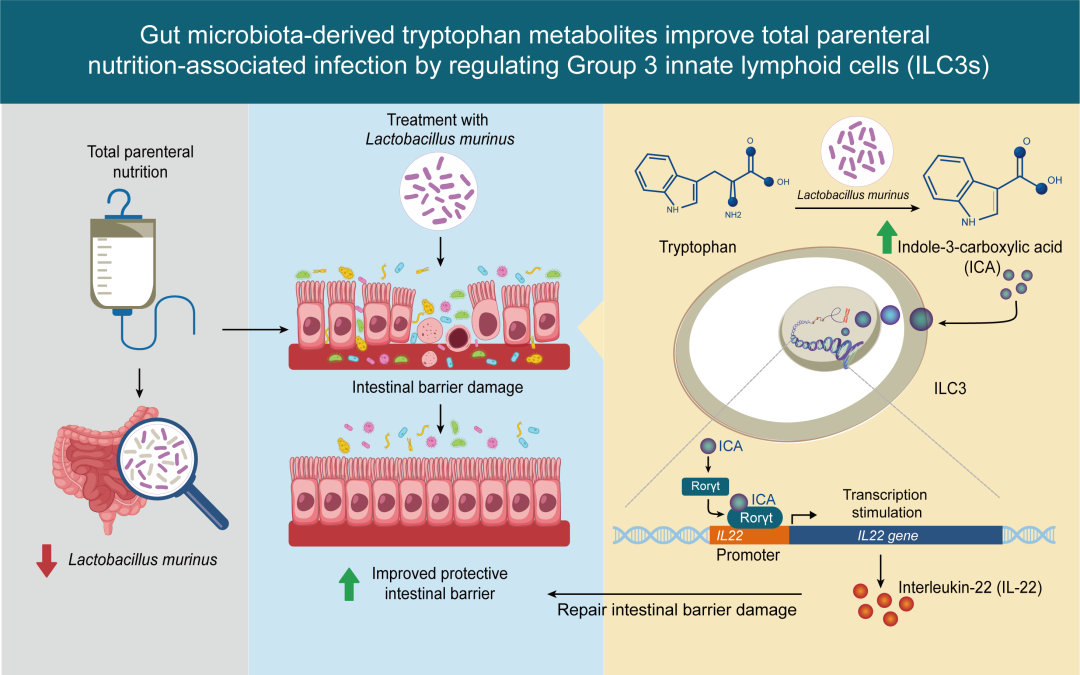
● 全肠外营养(TPN)引起的肠道微生物失调降低了肠道3型先天淋巴细胞(ILC3s)的反应性,从而增加感染风险;
● 进行TPN治疗的慢性肠衰竭(CIF)病人肠道中L. murinus丰度降低,而L. murinus及其代谢物吲哚-3-羧酸(ICA)可调节ILC3s,并改善感染状态;
● ICA可通过激活Rorγt进而促进ILC3s分泌IL-22。
摘 要
“临床营养支持”被克氏外科学称为20世纪外科实践中的四大关键进展之一。外科医生将临床营养视为一门“挽救生命”的学科,对于挽救危重症病人的生命和提高外科手术的成功率至关重要。肠外营养(PN)支持是临床营养治疗的重要组成部分,但全肠外营养(TPN)相关的一系列并发症可能对于病人治疗的效果有所影响。肠道稳态损伤与TPN相关感染的发生和发展密切相关,但其潜在机制仍不完全清楚。本研究中,通过RNA测序和单细胞RNA测序(scRNA-Seq)揭示了肠道3型先天淋巴细胞(ILC3s)分泌的白介素-22(IL-22)减少是导致TPN相关感染发生的重要因素。此外,通过对慢性肠功能衰竭病人肠道微生物群进行16S rRNA基因测序以及小鼠肠道微生物群的宏基因组测序分析,我们观察到TPN导致鼠乳杆菌(L. murinus)的丰度降低,而补充L. murinus则可以促进IL-22的分泌。机制方面,我们发现L. murinus产生吲哚-3-羧酸(ICA)增加,该物质可通过激活核受体Rorγt来刺激IL-22的分泌。这一通路能够增强肠道屏障的完整性,并减少感染风险。我们的研究结果加深了对TPN相关感染发生机制的理解,突出了肠道微生物群在维持机体免疫稳态和改善病人临床结局中的关键作用。
视频解读
Bilibili:https://www.bilibili.com/video/BV12t9AYfEgE/
Youtube:https://youtu.be/g5c6TnI7jqI
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
肠外营养(PN)是维持慢性肠功能衰竭(CIF)病人生命的重要治疗措施。当肠外营养提供超过80%的机体能量需求时,被称为全肠外营养(TPN)。然而,TPN可能损害肠道黏膜屏障的完整性,促进肠道细菌和内毒素的移位,进而引起威胁生命的严重并发症。TPN相关感染的发生率高达42.05%。已有研究表明,依赖TPN治疗的CIF病人其1.5年生存率仅为58%,其中感染是导致病人死亡的主要原因。由于TPN相关感染的具体机制尚不明确,且尚无有效的治疗方法,因此,探究TPN相关感染的发病机制对于改善使用TPN治疗的CIF病人预后至关重要。
肠道微生物群及其代谢产物与多种疾病相关,同时也是潜在的治疗靶点。肠道微生物群通过产生吲哚类化合物、胆汁酸和短链脂肪酸(SCFAs)等来调节宿主的免疫稳态。这些代谢物通过与芳香烃受体(AHR)、G蛋白偶联受体和法尼醇X受体等受体结合并激活相应的信号通路,刺激巨噬细胞、T细胞及相关细胞因子的招募,从而改善肠道屏障功能,减轻炎症。既往有研究报告显示,TPN小鼠的肠道微生物群中,厚壁菌门(Firmicutes)丰度下降,而拟杆菌门(Bacteroidetes)丰度增加,这也将TPN相关感染与肠道微生物群的改变联系起来。
肠道3型先天淋巴细胞(ILC3s)具有改善肠道屏障功能,整合细胞因子白介素(IL)-23和IL-1β所介导的信号传递等功能。这类细胞来源于肠道胶质细胞,可将信息传递给肠道上皮细胞(IECs)和肠道驻留细胞,进而改善肠道免疫并维持代谢稳态。ILC3s可通过微生物代谢物,如短链脂肪酸(SCFAs)和吲哚,激活并分泌IL-22,从而缓解结肠炎。既往有研究报道,TPN的使用将降低IL-22的表达;然而,ILC3s的变化是否与TPN相关感染有关仍不明确。本研究中,我们探究了ILC3s在TPN相关感染中的作用,同时阐明了鼠乳杆菌(L. murinus)产生吲哚-3-羧酸(ICA)进而激活ILC3s的具体机制。
结 果
TPN所引起的肠道微生物群失调导致肠道屏障损伤和感染
为了证实TPN可引起肠道屏障损伤和感染,我们对338例进行个体化营养支持的慢性肠功能衰竭(CIF)病人进行了回顾性分析(图S1)。根据肠外营养的供能比,将病人分为两组:高剂量肠外营养(H-PN)组(n = 123,肠外营养提供的能量大于总需求的80%)和低剂量肠外营养(L-PN)组(n = 215,肠外营养提供的能量小于等于总需求的80%)。对包括性别(p = 0.585)、年龄(p = 0.186)和基础疾病(p = 0.322)等在内的基线特征进行分析,两组在统计学上无显著差异(表S1)。如图1A所示,H-PN组的感染发生率明显较高(34.1%vs.10.2%,p < 0.001)。此外,H-PN组的肠道脂肪酸结合蛋白(IFABP)、白介素-6(IL-6)和C反应蛋白(CRP)水平较高(图1B,C,S2A)。我们还观察到IFABP与IL-6和CRP水平之间存在显著正相关(图1D,S2B)。这些结果表明,TPN的使用与临床不良事件的发生(如肠道屏障损伤和感染)密切相关。
随后,我们利用全肠外营养(TPN)干预的小鼠模型证明了TPN可引起肠道屏障受损、导致感染发生(图1E)。与对照组相比,TPN显著改变了小鼠肠道的组织学形态,导致TPN组小鼠的肠道组织病理学损伤评分增加,肠道绒毛缩短,隐窝变浅(图1F、G、S2C、D)。我们使用尤斯灌流系统评估了小鼠的肠道机械屏障功能,观察到在恒定电压条件下,Chow小鼠的跨膜电流显著弱于TPN小鼠(p < 0.001),这表明TPN组的小鼠肠道通透性明显增加(图1H)。我们还使用Cy5标记的D型丙氨酸(Cy5ADA)对小鼠肠道细菌进行了标记,随后进行免疫荧光染色,结果发现,TPN组小鼠中,肠道细菌突破了肠道黏膜下层,并入侵到肝脏中(图1I),这表明肠道屏障损伤严重。同时,我们观察到抗菌肽(Reg3γ,p = 0.0019)、紧密连接蛋白(Occludin,p = 0.0019和ZO-1,p = 0.0015)和溶菌酶(p = 0.0026)等肠道完整性标志物的表达显著下调(图1J-K,S2E)。通过酶联免疫吸附试验(ELISA),我们定量检测了小鼠血液中的细胞因子浓度,结果显示TPN处理导致血清中的IL-1β(p < 0.0001)、IL-6(p < 0.0001)和肿瘤坏死因子-alpha(TNF-α,p < 0.0001)的水平明显升高(图1L-N)。这些发现提示TPN干预引起肠道屏障损伤,造成感染的发生。
为了验证上述效应是否与肠道微生物群的变化有关,我们进行了粪便微生物群移植(FMT)(图S2F)。实验结果发现,将接受TPN干预的小鼠粪便移植给正常小鼠后,原本正常的小鼠表现出肠道屏障损伤(图1O,S2G-J)以及炎症标志物升高(图1P,p < 0.0001)。相反,接受Chow组小鼠粪便移植的小鼠并无相似变化。这些结果表明,TPN引起的肠道微生物群改变将导致肠道屏障损伤及感染发生。
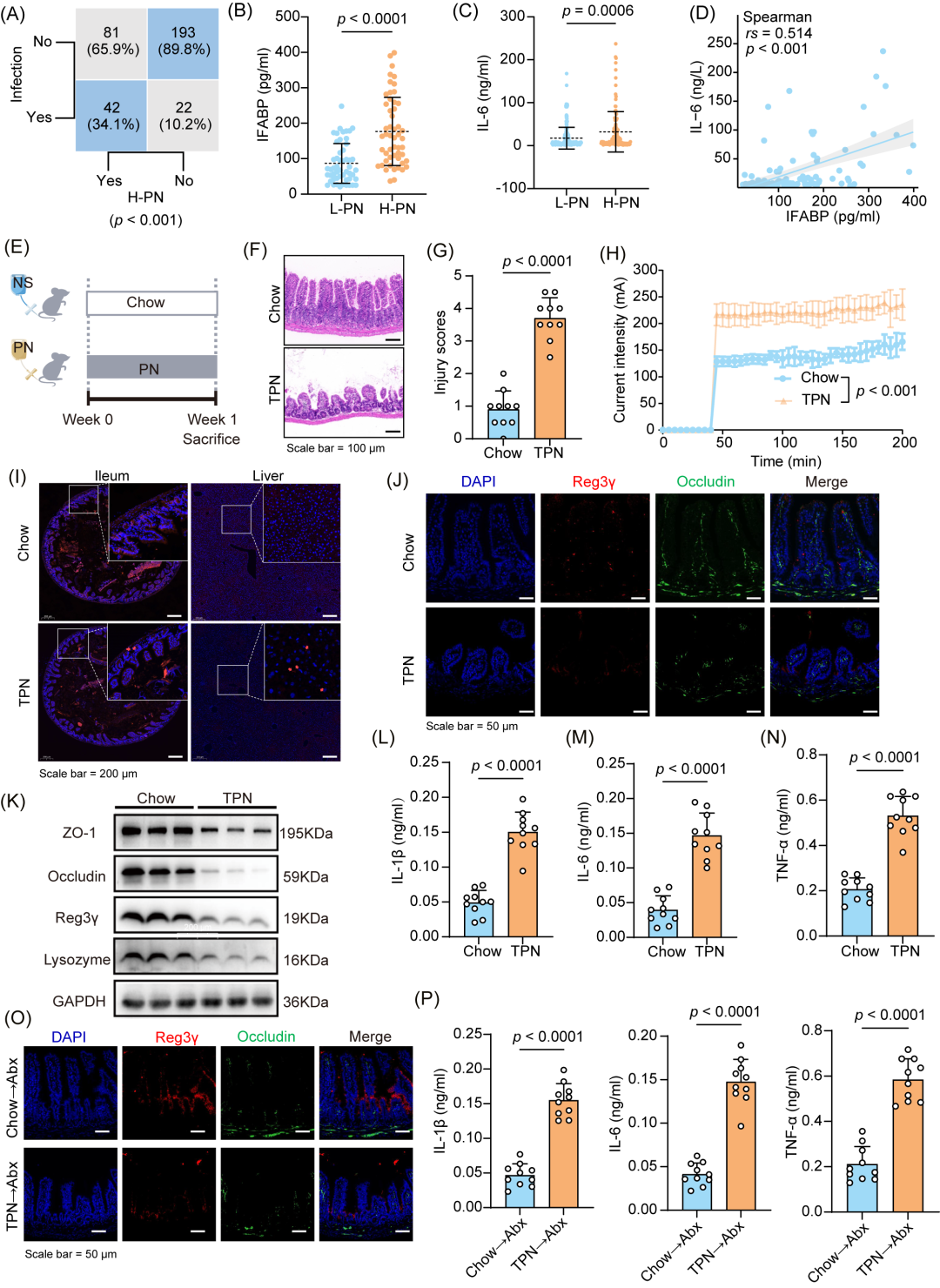
图1.全肠外营养(TPN)在人体和小鼠体内引起感染
(A)在慢性肠功能衰竭(CIF)病人中,高剂量肠外营养(H-PN)组感染发生率更高。H-PN组:高剂量肠外营养组(n = 123,PN提供的能量 > 80%机体总所需量)。L-PN组:低剂量肠外营养组(n = 215,PN提供的能量 ≤ 80%机体总所需量)。(B-C)病人血清中肠脂肪酸结合蛋白(IFABP)(n = 52人/组)和白细胞介素-6(IL-6)的水平(H-PN组n = 123,L-PN组n = 215)。(D)病人IFABP和IL-6水平的相关性分析。(E)对C57BL/6小鼠的实验设计。(F-G)代表性小肠苏木精-伊红(HE)染色图像(F)及相应的组织病理学损伤评分(G)(n = 10只/组)。(H)采用尤斯灌流系统测量小鼠肠道跨膜电流(n = 5只/组)。(I)小鼠肠道和肝脏中的细菌免疫荧光检测。(J)免疫荧光分析标准饲料(Chow)饲喂和TPN干预条件下的小鼠肠道中Reg3γ和Occludin的表达。(K)Western blot对比Chow组与TPN组小鼠肠道中ZO-1、Occludin、Reg3γ和溶菌酶(Lysozyme)的表达水平。GAPDH作为内参对照。(L-N)Chow组与TPN组小鼠血清中白细胞介素-1β(IL-1β)(L)、IL-6(M)及肿瘤坏死因子-α(TNF-α,N)的水平(n = 10只/组)。(O)免疫荧光分析抗生素处理(Abx)小鼠肠道中Reg3γ和Occludin的表达。(P)Abx小鼠在不同干预后血清IL-1β、IL-6和TNF-α水平(n = 10只/组)。p值的计算方法:Mann-Whitney U检验(B和C),Student’s t检验(G-H和L-M)。
肠道微生物群失调影响了肠道ILC3s的反应性
为了探究TPN引起肠道屏障损伤和感染风险增加的机制,我们对肠道RNA测序数据进行了功能分析。基因本体论(GO)分析结果发现,免疫反应和炎症反应相关的通路显著富集(图2A)。京都基因与基因组百科全书(KEGG)通路分析和基因集富集分析(GSEA)揭示了肠道先天免疫系统的显著变化(padj = 0.040,图2B,C)。为了探究TPN干预如何影响机体的免疫系统,我们对RNA测序数据进行了进一步分析,发现在差异排名前10位的候选细胞因子中,TPN组小鼠的IL-22表达水平显著降低(图2D,S3A)。ILC3s和其他类型的细胞(包括γδT细胞、辅助T17(Th17)细胞和Th22细胞)均可分泌IL-22。但是,两组小鼠的肠道中并未观察到Th17(p = 0.2035,图S3C,D)、Th22(p = 0.0941,图S3E,F)和γδT细胞(p = 0.1233,图S3G,H)的比例存在差异,而ILC3s的比例则变化明显(p < 0.0001,图S4A-E)。因此,TPN引起的ILC3s变化可能是促进肠道屏障损伤和炎症的发展的原因。我们进一步评估了两组小鼠中ILC3s分泌IL-22的能力,结果显示,TPN使IL-22+ ILC3s的数量和比例显著降低(p < 0.0001,图2E,F),而ILCs的则未受影响(图S4B,C)。此外,免疫荧光分析显示,TPN干预后小鼠肠道中的IL-22和Rorγt表达水平显著降低(图2G)。TPN干预后小鼠血清中IL-22的水平也明显下降(图S4F)。在高剂量肠外营养(H-PN)支持的病人中也观察到了一致的结果(图2H)。
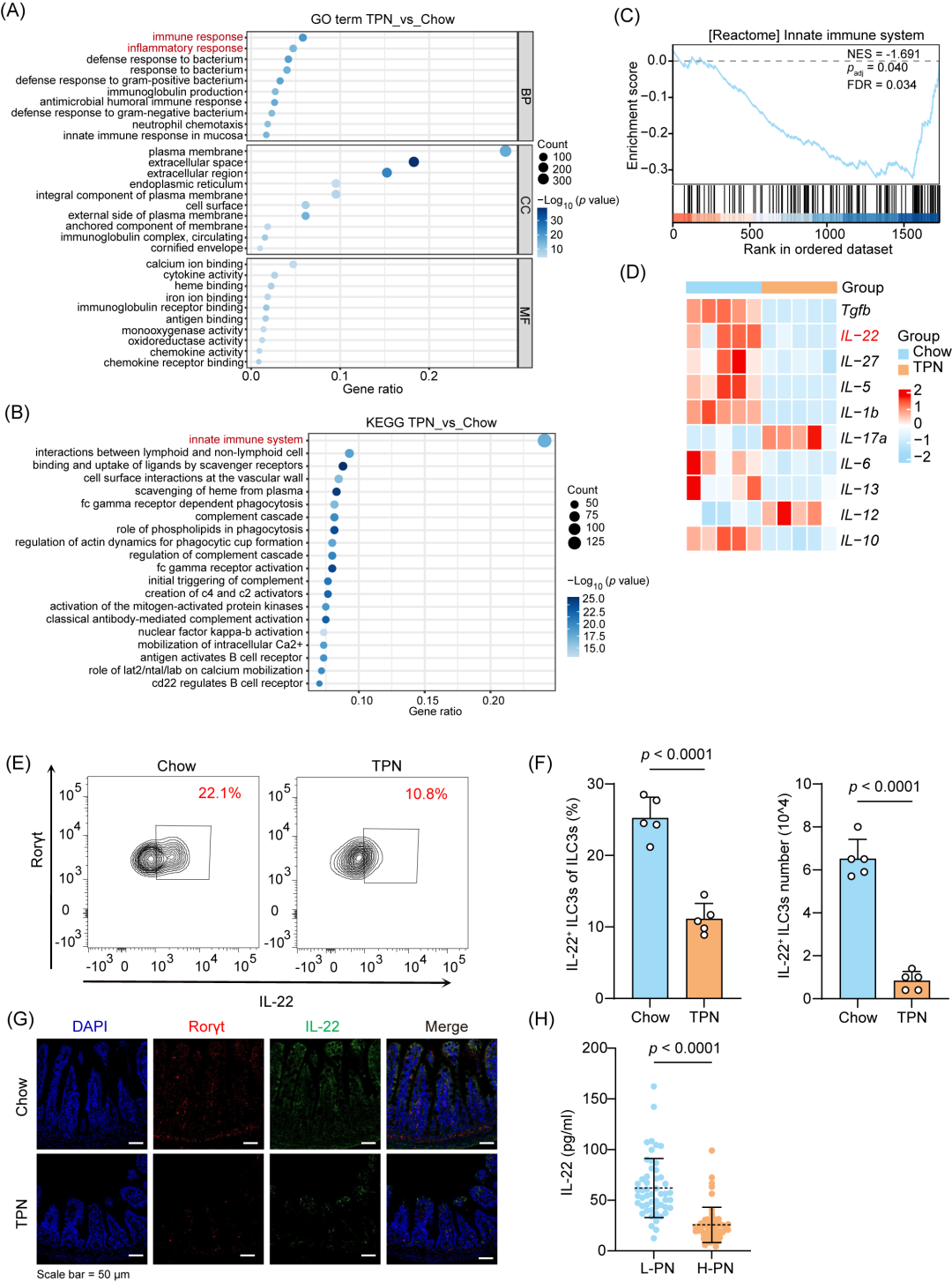
图2.TPN诱导的微环境以3型先天淋巴细胞(ILC3s)反应性降低为特征
(A-B)对小鼠肠道组织(n = 5只/组)进行基因本体论(GO)分析(A)和京都基因与基因组百科全书(KEGG)通路分析(B)。GO分析显示其中两个显著富集的通路与免疫应答和炎症反应相关。KEGG通路分析提示肠道先天免疫系统产生明显变化。(C)小鼠肠道组织的基因集富集分析(GSEA)(n = 5只/组)。(D)热图显示Chow组与TPN组小鼠肠道细胞因子mRNA的表达差异(n = 5只/组)。(E-F)代表性流式细胞分选(FACS)图(E)及不同处理条件下小鼠肠道IL-22+ ILC3s频率(F)(n = 5只/组)。(G)免疫荧光分析不同处理条件下小鼠肠道中Rorγt和IL-22的表达。(H)病人血清IL-22水平(n = 52人/组)。p值的计算方法:Student’s t检验(F)和Mann-Whitney U检验(H)。
为了进一步研究ILC3s的具体变化,我们对肠道ILCs进行了单细胞测序分析。肠道ILCs被分为七个细胞亚群(NCR+ ILC3、NCR- ILC3、ILC1/2、NK、LTi和ILCP)(图3A)。我们鉴定了各个细胞亚群中表达最为显著的前20个基因及其特征性标志物。其中,Tbx21和Itga1是NK细胞和ILC1的特征基因,Gata3和E2f1是ILC2的特征基因,而NCR+ ILC3s的特征基因是Rorc、Ccr6和Maf(图3B,C)。我们观察到TPN组中NCR+ ILC3s的比例减少(图3D,E)。同时,IL-22的表达水平也相应下降(图3F,G),这提示NCR+ ILC3s分泌的IL-22对于肠道屏障可能具有保护作用。随后,我们进一步分析了FMT后小鼠肠道ILC3s亚群的变化。结果显示,接受TPN处理小鼠粪便移植的小鼠中IL-22+ ILC3s的比例显著下降(p < 0.0001,图3H,I),IL-22水平也明显降低(p < 0.05,图S4G-J)。这些结果表明,TPN影响肠道微生物群,导致肠道免疫微环境中ILC3s反应性减弱。这种调节可能引起肠道屏障完整性的损害,并增加感染风险。
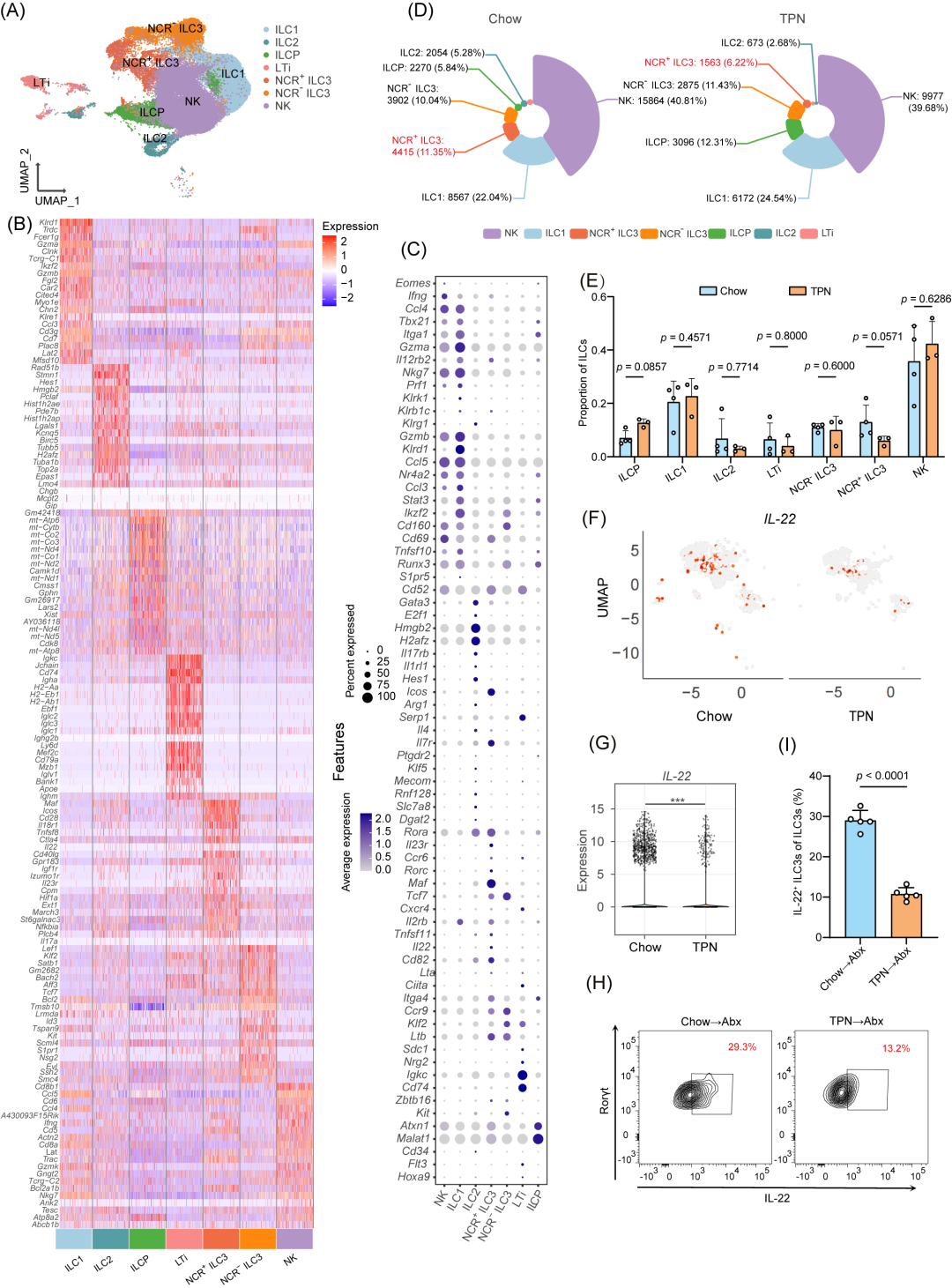
图3.Chow组与TPN组ILCs的单细胞RNA测序(scRNA-Seq)分析
(A)UMAP可视化分析不同部位ILC亚群的整体分布。(B)细胞类型特异性基因中前20位标志基因热图。(C)细胞簇中代表性差异表达基因(DEGs)。点的大小表示在该簇中表达该基因的细胞比例,颜色表示基因的平均相对表达水平。(D)Chow组(n = 4)与TPN组(n = 3)中不同ILC细胞类型的百分比。(E)Chow组(n = 4)与TPN组(n = 3)中每个样本的ILC细胞类型百分比。(F)UMAP分析NCR⁺ ILC3s,结果显示Chow组的IL-22基因表达水平高于TPN组。(G)小提琴图显示NCR⁺ ILC3s中IL-22的表达水平。(H-I)代表性荧光活化细胞分选(FACS)图(H)及不同干预方法后小鼠肠道固有层IL-22⁺ ILC3s的百分比(I)(n = 5只/组)。***,p < 0.001。p值的计算方法:Mann-Whitney U检验(E),Student’s t检验(I)。
TPN导致了肠道中的L. murinus丰度降低
为了进一步筛选差异微生物物种,我们对病人粪便样本进行了16S核糖体RNA(16S rRNA)测序,并分析了TPN干预后肠道微生物群的变化(图S5A)。尽管两组病人的Alpha(α)多样性无显著差异(图S5B),但主坐标分析提示TPN干预对于病人的肠道微生物群构成具有明显影响(图4A)。使用线性判别分析效应大小(LDAEffect Size,LEfSe)分析了细菌组成,在属水平上,两组之间肠道微生物群差异主要存在10个差异菌属(图4B,C)。
小鼠的宏基因组测序揭示了两组之间微生物群分布存在显著差异(图4D,S5C,p < 0.05)。线性判别分析(LDA)值分布的条形图显示,在属水平上,Chow组中有19个特征性差异菌属,而TPN组中存在5个特征性菌属。其中值得注意的是,Lactobacillus属在TPN组中表现出较高的LDA值(图4E,S5D,E)。
此外,我们根据每个样本或组别中前10个属制作了桑基图,结果显示,接受高剂量肠外营养(H-PN)治疗的病人和TPN干预的小鼠中Lactobacillus的数量均有所减少(图4F,S5F-H)。对于不同菌属丰度和病人临床特征进行相关性分析,结果发现,Lactobacillus的丰度减少与病人发生感染之间存在显著关联(p < 0.001,图4G)。基于上述结果推测,Lactobacillus的丰度减少可能是TPN引起感染风险增加的一个重要原因。对于小鼠进行宏基因组分析,其结果显示,Lactobacillus属中有四个菌株(murinus、reuteri、johnsonii和animalis)在菌株水平上存在差异(图4E,S5I,p < 0.05)。
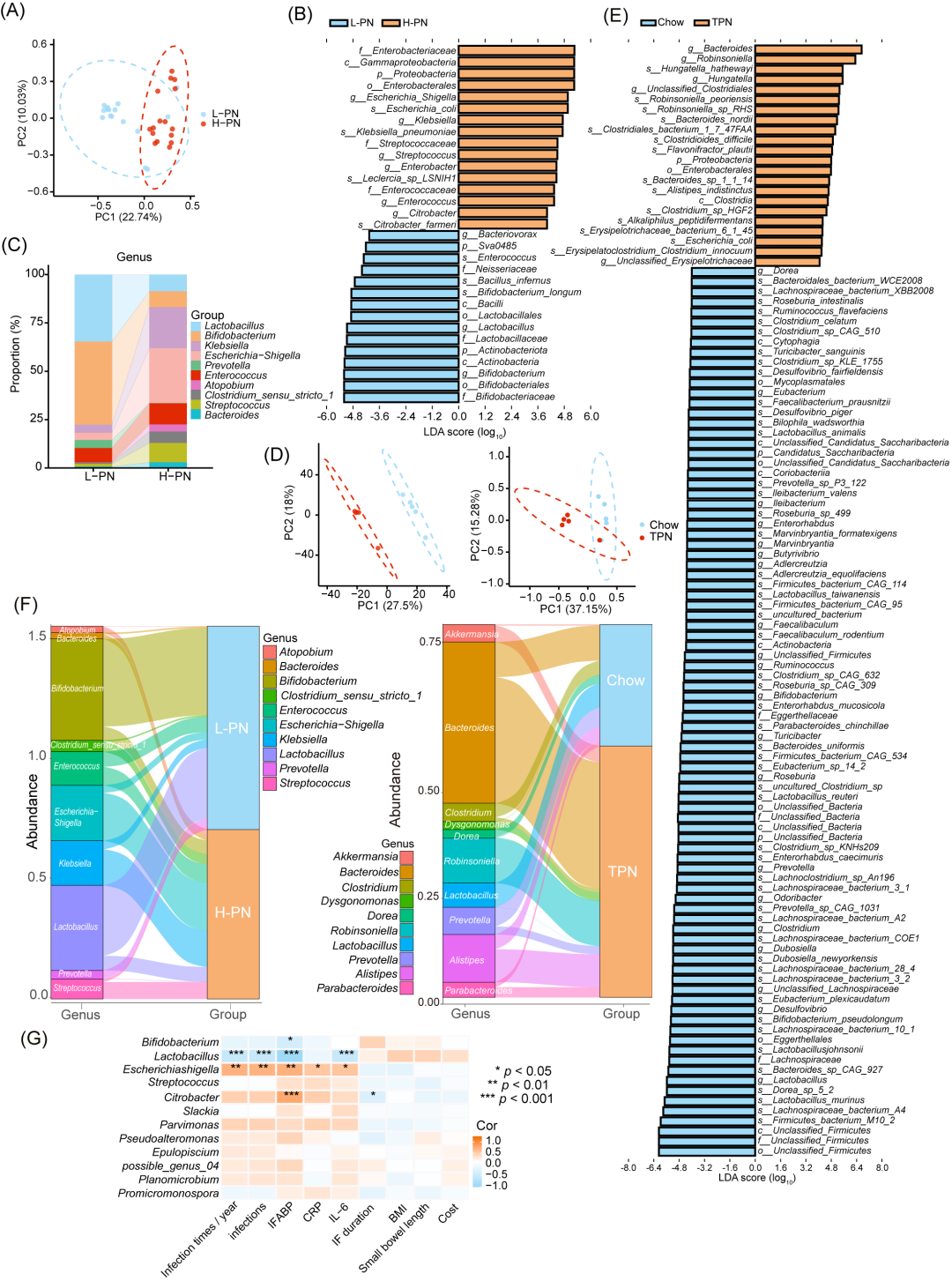
图4.TPN引起人类和小鼠肠道菌群失调
(A)病人粪便样本(n = 16人/组)16Sr RNA基因序列的主坐标分析(PCoA)。(B)通过线性判别分析(LDA)鉴定病人肠道菌群的显著性差异菌群。(C)16s分析中丰度最高的前10属细菌。(D)PCoA分析显示小鼠(n = 5只/组)肠道菌群在属水平(左图)和种水平(右图)的相对丰度。(E)小鼠肠道丰度菌群的LDA分析。(F)桑基图展示人类和小鼠粪便样本中丰度最高的前10属水平细菌。(G)热图显示病人肠道菌群中特定菌种的丰度与慢性肠功能衰竭(CIF)病人临床特征之间的相关性。
代谢活跃的L. murinus通过调节ILC3s改善TPN相关感染
为了研究Lactobacillus在TPN相关感染中的作用,我们对TPN小鼠进行了菌株干预(图S6A)。我们发现,L. murinus对于肠道屏障的完整性具有保护作用(图S6B-D)。此外,我们发现,H-PN组病人粪便样本中的L. murinus丰度显著降低(p < 0.0001,图5A)。我们进一步分析了CIF病人的临床特征与L. murinus的相关性,发现L. murinus丰度与病人感染率之间存在明显负相关(p = 0.0017,图S6E)。这些结果表明,在临床病人中L. murinus减少与TPN相关肠道屏障损伤和感染风险增加存在关联。
为了进一步研究L. murinus的作用,我们将磷酸盐缓冲盐水(PBS)、灭活的L. murinus(i-L.m)或活的L. murinus(live L.m)给TPN小鼠进行了灌胃(图5B)。我们发现,只有活的L. murinus能够改善肠道黏膜病理改变(图5C,D,S6F),同时可上调ZO-1、Occludin、Reg3γ和溶菌酶等肠道屏障完整性标志物的表达(图5E,S6G,H)。此外,活的L. murinus还可以降低炎症因子的水平(p < 0.0001,图5F-H),表明只有代谢活跃的L. murinus才能发挥肠道屏障保护作用。
对病人的IL-22表达水平和L. murinus丰度进行了进一步分析,结果显示两者之间存在正相关(rs = 0.689,p < 0.001,图S6I)。在使用活L. murinus干预后,IL-22+ ILC3s的比例显著增加(p < 0.0001,图5I)。免疫荧光实验进一步表明,活L. murinus可上调肠道中Rorγt和IL-22的表达(图5J)。这些发现表明,活的L. murinus可以增强IL-22+ ILC3s的分化,从而改善肠道屏障功能并提高机体抗感染能力。
L. murinus的代谢产物ICA在改善TPN相关肠道屏障损伤中发挥重要作用
为了研究TPN治疗后小鼠肠道菌群代谢物的变化,我们使用液相色谱-质谱联用技术(LC-MS)分析了其粪便中代谢物的组成。3D主成分分析(3D-PCA)和火山图分析显示,TPN组和Chow组之间存在明显的代谢特征差异,共鉴定出218种差异代谢物(Chow组vs.TPN组,117种上调,101种下调)(图5K,L)。我们对这218种差异代谢物进行了KEGG通路分析,共识别出了19条富集的代谢通路,前五条分别为氨基糖苷和核苷糖代谢、色氨酸代谢、组氨酸代谢、甘油磷脂代谢和果糖与甘露糖代谢(p < 0.001,图5M)。其中,色氨酸代谢通路差异最大,p = 0.0009。
色氨酸代谢通路对于肠道微生物群对宿主代谢和免疫稳态的影响至关重要。细菌菌株与色氨酸代谢物的相关性分析显示,ICA、吲哚-3-乙酰醛和吲哚-3-丙酸等吲哚化合物与L. murinus之间存在显著正相关(p < 0.05,图5N)。其中,ICA在变量重要性投影(VIP)得分中分值最高(图5O),且其水平与粪便中L. murinus丰度表现出显著正相关性(rs = 0.721,p < 0.001,图S6J)。在接受高剂量肠外营养(H-PN)治疗病人的粪便中,ICA水平下降(图S6K),而在活L. murinus干预后,TPN小鼠粪便中的ICA水平也显著增加(p < 0.0001,图S6L)。因此,我们推测,在CIF病人及TPN小鼠中,L. murinus可能通过产生ICA保护肠道屏障,降低感染易感性。
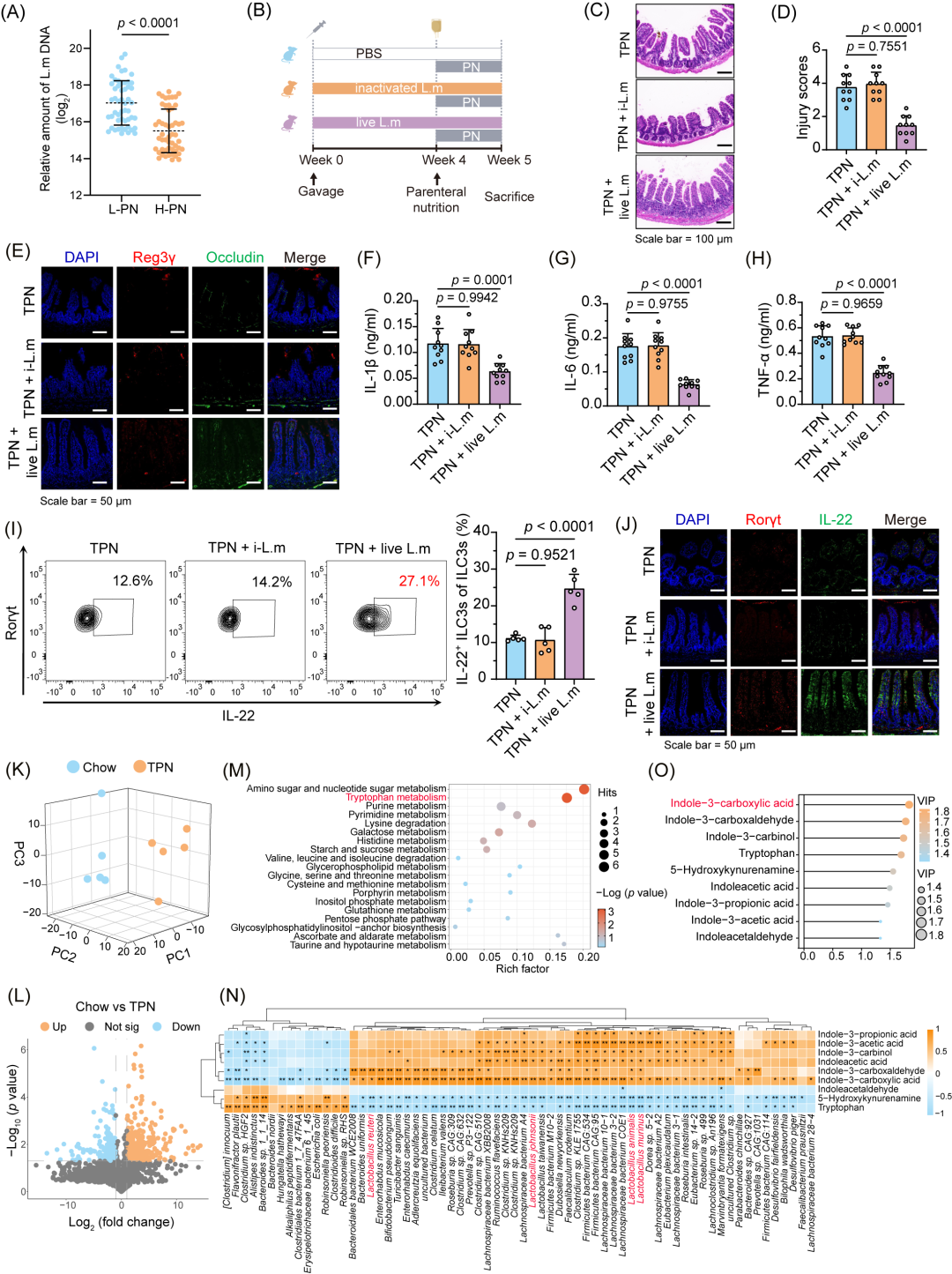
图5.具有代谢活性的L. murinus通过增强ILC3s功能改善肠道屏障损伤
(A)采用实时PCR(RT-PCR)检测低剂量肠外营养(L-PN)和高剂量肠外营养(H-PN)病人粪便中L. murinus的丰度(n = 52人/组)。(B)活性或灭活L. murinus干预的小鼠实验设计方案。(C-D)代表性肠道HE染色图像(C)及肠道损伤评分(D)(n = 10只/组)。(E)免疫荧光分析不同处理条件下小鼠肠道中Reg3γ和Occludin的表达。(F-H)给予不同处理后TPN小鼠血清中IL-1β(F)、IL-6(G)和TNF-α(H)的水平(n = 10只/组)。(I)代表性荧光活化细胞分选(FACS)图及不同处理后小鼠肠道IL-22⁺ ILC3s的百分比(n = 5只/组)。(J)免疫荧光分析不同处理后TPN小鼠肠道中Rorγt和IL-22的表达。(K-L)3D主成分分析(3D-PCA)(K)及火山图(L)展示Chow组与TPN组小鼠粪便的差异代谢特征。(M)依据来自Chow组与TPN组小鼠粪便的218种差异代谢产物,对代谢组通路进行富集分析(n = 5只/组)。(N)热图展示肠道菌群(物种水平)的丰度与Chow组小鼠富集的9种差异性色氨酸代谢产物之间的相关性。(O)变量重要性投影(VIP)评分基于3D-PCA评估代谢物对样本分类的贡献。VIP评分 > 1.5的分类变量在区分过程中具有显著意义。p值的计算方法:Kruskal-WallisH检验(Dunnett’s t检验校正)(A),单因素方差分析(One-way ANOVA,Dunnett’s t检验用于事后分析)(D和F-I)。所有统计检验均为双侧检验。
为了验证这一假设。我们给TPN小鼠进行了ICA或PBS口服灌胃(图6A)。口服ICA可使粪便中的ICA浓度达到与活L. murinus干预相似的水平(图S7A,S6L)。ICA治疗改善了肠道黏膜病理损伤(图6B,S7B,C),同时上调了ZO-1、Occludin、Reg3γ和溶菌酶等肠道屏障完整性标志物的表达(图6C,S7D,E)。此外,ICA治疗还可以显著降低机体炎症因子的水平(p < 0.0001,图6D)。ICA干预后,IL-22+ ILC3s的比例显著增加(p < 0.0001,图6E),并上调了肠道中IL-22的表达水平(图6F)。
芳香族氨基酸氨基转移酶(ArAT)是L. murinus产生ICA所必须的关键酶(图S7F)。因此,为了确定ArAT失活是否会使L. murinus对肠道屏障的保护作用消失,我们构建了一个无功能的ArAT突变株(图S7G,H)。我们对L. murinus ΔArAT干预后的TPN小鼠的粪便样本进行了LC-MS分析(图6G),正交偏最小二乘法-判别分析(Orthogonal Partial Least Squares-Discriminant Analysis,OPLS-DA)显示,野生型和突变型L. murinus干预后,小鼠的代谢谱存在显著不同(图6H)。在敲除L. murinus的ArAT后,TPN小鼠粪便中的ICA水平显著降低(图6I);同时,这些小鼠的肠道病理损伤和与肠道屏障相关的标志物均无改善(图6J,S7I-L)。此外,炎症标志物的表达(p > 0.01,图6K-M)和IL-22+ ILC3s的比例(p = 0.6802,图6N)也没有显著变化。这表明在ΔArAT敲除后,L. murinus丧失了对于肠道屏障的保护作用。总而言之,ArAT及ICA对L. murinus保护肠屏障、改善TPN相关感染至关重要。
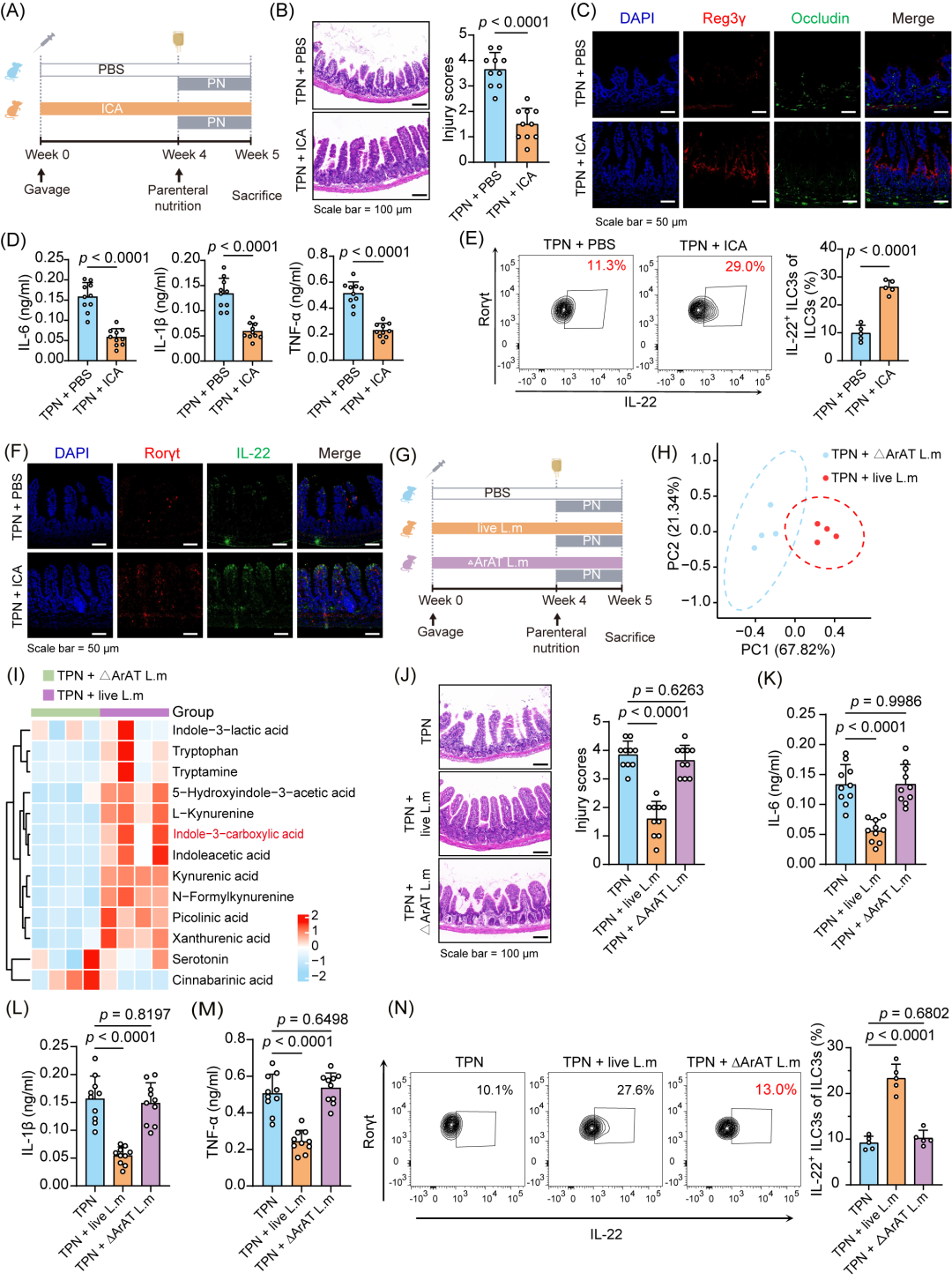
图6.吲哚-3-羧酸(ICA)是调节ILC3s的关键代谢物
(A)ICA干预的小鼠实验设计方案。(B)代表性小鼠肠道HE染色图像及损伤评分(n = 10只/组)。(C)免疫荧光分析PBS或ICA处理后TPN小鼠肠道中Reg3γ和Occludin的表达。(D)不同处理条件下TPN小鼠血清中IL-1β、IL-6和TNF-α的水平(n = 10只/组)。(E)代表性荧光活化细胞分选(FACS)图及不同处理后小鼠肠固有层IL-22⁺ ILC3s的百分比(n = 5)。(F)免疫荧光分析不同处理后TPN小鼠肠道中Rorγt和IL-22的表达。(G)活菌或ΔArAT L. murinus干预的小鼠实验设计方案。(H)偏最小二乘判别分析(PLS-DA)用于评估接受ΔArAT或活L. murinus处理的TPN小鼠粪便代谢物特征(n = 4只/组)。(I)色氨酸代谢组学分析的热图,显示接受ΔArAT或活L. murinus处理的TPN小鼠粪便样本的代谢物变化(n = 4只/组)。(J)代表性小鼠肠道HE染色图像及损伤评分(n = 10只/组)。(K-M)不同处理后TPN小鼠血清中IL-1β(K)、IL-6(L)和TNF-α(M)的水平(n = 10只/组)。(N)代表性FACS图及不同处理后小鼠肠道IL-22⁺ ILC3s的百分比(n = 5)。p值计算方法:Student’s t检验(B和D-E),单因素方差分析(One-way ANOVA,Dunnett’s t检验用于事后分析;J-N)。
ICA通过靶向Rorγt促进ILC3s分泌IL-22
为了进一步研究L. murinus和ICA是否通过调节ILC3s发挥作用,我们构建了Rorγt敲除小鼠(Rorc-/-)从而耗竭ILC3s。随后,对Rorc-/-小鼠进行了TPN建模和干预(图S8A),结果发现,ILC3s的敲除使L. murinus及其代谢物ICA对肠道屏障的保护作用受到抑制(图S8B-D)。
接下来,为了探索ICA增强ILC3s功能的机制,我们从小鼠肠道中分离了ILC3s,并进行了体外干预实验(图7A)。与对照组相比,ICA处理后,IL-22+ ILC3s的比例增加(图7B,C)。基于单细胞RNA测序数据对NCR+ ILC3s进行差异基因分析后,我们确定了可能参与调节IL-22分泌的基因。在这些基因中,我们观察到TPN组中Rorc的表达显著下降(p < 0.0001,图7D)。相比之下,另一个可能参与ILC3s分泌IL-22的关键转录因子芳烃受体(Ahr)的表达水平在Chow组和TPN组小鼠的NCR+ ILC3s中类似(图S8E)。这一发现提示,ICA可能通过Rorc调节ILC3s功能。同时,体内和体外实验均表明,ICA可显著提高Rorc的mRNA和蛋白水平(图S8F-I)。随后,我们评估了在Rorγt抑制剂GSK805(0.5µM)处理后,ICA对IL-22+ ILC3s的影响(图7E),发现GSK805抑制了ICA诱导IL-22+ ILC3s分化的能力(图7F)。
接下来,我们评估了ICA是否会影响Rorγt与其靶基因组元素的结合。通过染色质免疫沉淀结合定量聚合酶链式反应(ChIP-qPCR),我们评估了Rorγt在IL-22启动子区域的占位情况,发现ICA明显增强了Rorγt与该位点的结合(图7G,S8J)。分子对接模拟显示,ICA可能与Rorγt形成稳定的复合物,其结合能为-6.1kcal/mol(图7H)。荧光素酶报告基因实验进一步显示,ICA可激活Rorγt的转录活性(图7I)。上述结果表明,ICA可能通过增强Rorγt的转录活性,从而促进IL-22的表达(图S8K)。
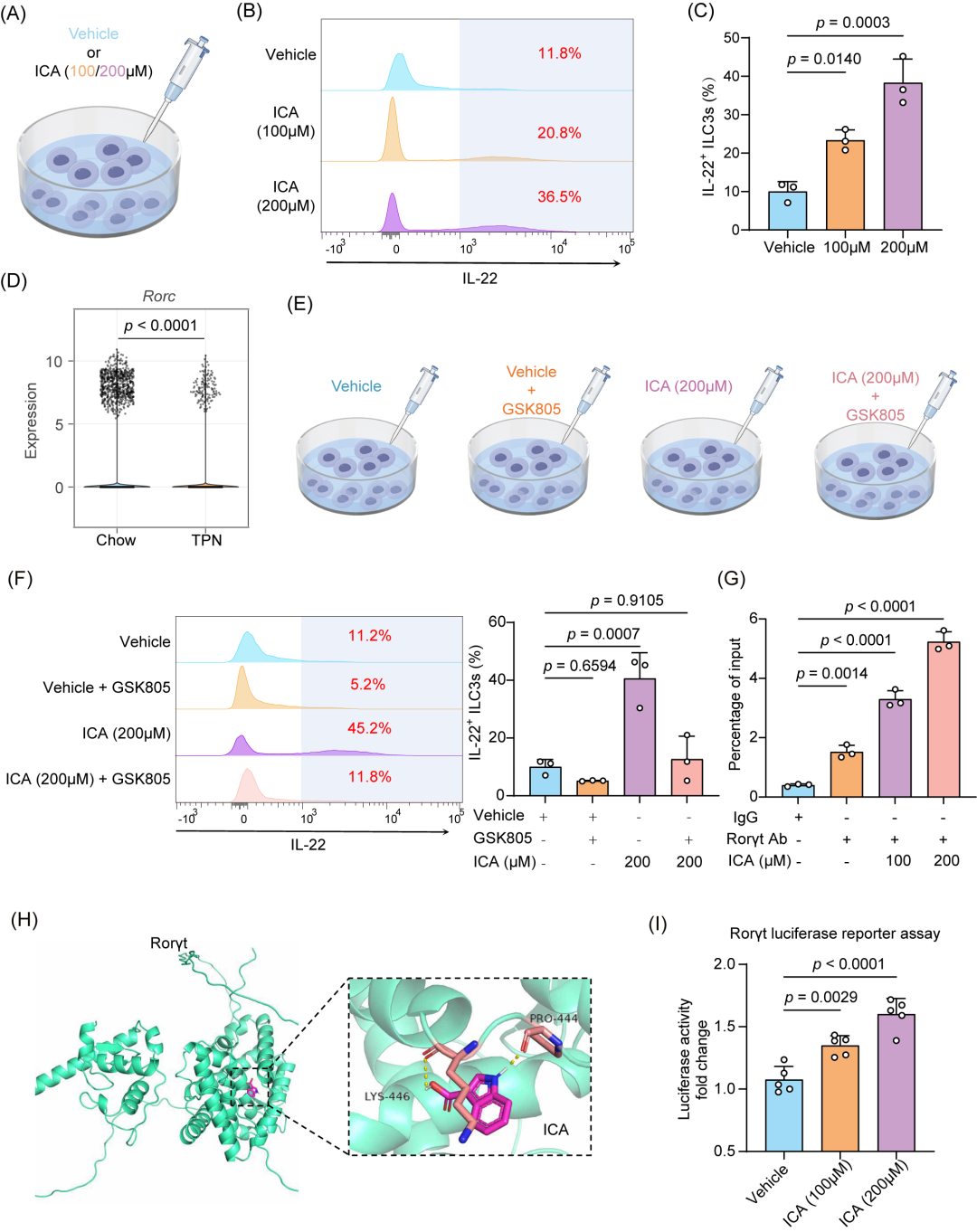
图7.ICA通过靶向Rorγt促进ILC3s的功能
(A)用不同浓度的ICA处理ILC3s。(B-C)代表性FACS图(B)及处理72小时后不同浓度ICA处理的IL-22⁺ ILC3s比例(C)。(D)小提琴图显示NCR⁺ ILC3s中Rorc的表达情况。(E)用ICA(200µM)、GSK805(0.5µM)或两者联合处理ILC3s。(F)代表性FACS图及不同处理后IL-22⁺ ILC3s的比例。(G)ChIP-qPCR分析ICA对Rorγt结合IL-22基因启动子的影响(n = 3)。(H)通过分子对接识别ICA作为潜在的Rorγt激动剂,显示在Rorγt结合口袋内的疏水范德华相互作用(碳原子以红色显示)。(I)荧光素酶报告基因实验表明ICA激活Rorγt的转录活性。将含有Rorγt结合序列的PGL3构建体转染入HEK-293细胞,然后用溶剂(DMSO)或指定浓度的ICA处理细胞,随后进行荧光素酶活性检测。数据以平均值±标准误(n = 5)表示。p值通过单因素方差分析(One-way ANOVA,事后分析采用Dunnett’s t检验;C、F-G和I)计算。所有统计检验均为双侧检验。
讨 论
TPN(静脉营养支持)治疗可能损伤肠道屏障,促进细菌和内毒素进入全身循环,从而引发感染并导致病人的不良临床结局。预防TPN相关的肠道屏障损伤并减少TPN相关感染对于治疗CIF病人至关重要。既往已有研究报告表明,TPN会导致肠道微生物群失调,但TPN引起肠道微生物群失调如何导致肠道屏障损伤及其后续感染的具体机制仍未被阐明。
本研究中,我们观察到TPN干预影响小鼠的肠道先天免疫功能。此外,通过流式细胞术和单细胞RNA测序(scRNA-seq)分析,我们发现TPN治疗将导致肠道IL-22+ ILC3s的数量和比例下降。许多研究已证实ILC3在肠道中发挥保护作用。这类细胞能够整合细胞因子介导的信息,并将其传递给肠上皮细胞(IEC)和驻留免疫细胞,从而增强肠道免疫功能并维持代谢稳态。既往研究表明,活化的ILC3所产生的细胞因子,尤其是IL-22,能够刺激肠上皮细胞产生抗菌肽(如Reg3γ)和粘蛋白,从而维持肠道屏障的功能。因此,我们假设,ILC3功能的抑制与TPN相关感染密切相关。然而,ILC3在TPN相关感染中的确切作用仍有待阐明。
进一步的研究表明,TPN引起的肠道菌群失调导致了ILC3功能的改变,并与TPN相关的感染有关。将TPN组小鼠的粪便微生物群移植到Abx小鼠中,结果导致肠道中IL-22+ ILC3s的比例下降,从而增加了其肠道屏障损伤和感染易感性。与之相反的是,移植Chow组供体的粪便微生物群并不会引起这些改变。此外,在接受TPN治疗的临床病人和TPN小鼠的粪便样本中均观察到L. murinus的减少,这表明L. murinus可能在调节ILC3功能中发挥着关键作用,并与TPN相关的感染有关。据我们所知,此前尚无研究表明L. murinus可以调节ILC3并改善TPN相关的感染。其他研究还观察到宿主与微生物群之间存在相互作用。研究人员发现,阿司匹林治疗后,肠道中Parabacteroides goldsteinii的丰度下降,同时抑制了胆汁酸代谢并导致肠道屏障损伤。
L. murinus可调节色氨酸代谢,并在多种疾病中发挥保护作用,该效果是由于其具有抗菌活性和免疫调节功能。L. murinus可以通过产生吲哚乳酸来抑制Th17细胞的分化,从而预防盐敏感性高血压。激活TLR2还可以诱导巨噬细胞释放IL-10,促进肠道缺血/再灌注损伤的修复。近期的研究表明,L. murinus与肠道屏障功能之间存在正相关。此外,L. murinus通过分泌细胞外囊泡调节巨噬细胞极化,并改善化疗所导致的肠道炎症。这些研究促使我们对于L. murinus如何改善ILC3s功能和TPN相关感染进行探索。
我们的研究表明,ICA是L. murinus重要代谢物,此前并无ICA改善ILC3s功能的相关研究报道。于君教授等人指出,由瑞士乳酸菌(Lactobacillus helveticus)产生的ICA可通过抑制CD4+ Treg细胞的分化,增强CD8+ T细胞的功能,从而提高结直肠癌中抗PD-1治疗的疗效。此前的研究仅报道了ICA对植物免疫功能的调节作用,本研究与于君教授等人的研究一起,揭示了ICA在动物或人类疾病免疫调节中的潜在应用前景。
我们发现,L. murinus及其衍生物ICA可以直接靶向Rorγt,促进ILC3s分泌IL-22,进而改善肠道屏障损伤,减少TPN引起的感染。既往研究表明,肠道微生物衍生物既可以作为部分Ahr激动剂,也可以作为竞争性Ahr抑制剂,调节Ahr的激活,从而影响免疫细胞的功能。然而,我们的研究提出了一种新的机制,即肠道微生物及其衍生物通过直接调节免疫细胞来保护肠道屏障,并减少感染。
我们的研究也存在一些局限性。ILC3s改善TPN相关肠道屏障损伤的具体机制仍需进一步探索。现有证据表明,ILC3s通过多种机制增强肠道免疫并减轻炎症,例如保护肠道干细胞免受基因应激,分泌粒细胞-巨噬细胞集落刺激因子或IL-2,调节T细胞和巨噬细胞。然而,关于ILC3s如何改善TPN引起的肠道屏障损伤的具体机制仍需进一步阐明。
此外,我们的研究还观察到,TPN导致IL-1β、IL-5、IL-27和IL-17A等细胞因子的明显变化。这些变化显示了TPN治疗将对复杂的肠道免疫功能调节网络产生影响。尽管我们的研究重点是分泌IL-22的免疫细胞,但我们并未进一步研究其他细胞因子产生细胞的贡献,如产生IL-1β的巨噬细胞和中性粒细胞,或产生IL-5的嗜酸性粒细胞和肥大细胞等。这些相互作用及其对肠道屏障功能的影响可在未来的研究中进行系统性的探索。
最后,研究中的另一个局限之处在于,未能使用STOP/CD4小鼠模型进行ILC3s的靶向干预。未来的研究应对此进行补充,这将帮助我们更精确地理解ILC3s特异性机制,同时,对于ICA在TPN相关肠道屏障功能障碍中的治疗潜力提供了更深入的见解。
结 论
本研究证实了L. murinus及其色氨酸代谢产物ICA可促进IL-22+ ILC3s的分化,从而保护肠道屏障并降低TPN相关感染风险。此外,我们还证明了ICA可通过激活Rorγt促进ILC3s分泌IL-22,进而减轻TPN引起的肠道屏障损伤。这些发现阐明了宿主-微生物群代谢-免疫调节的全新机制,并强调了L. murinus或ICA作为辅助治疗改善CIF病人TPN相关感染的潜力。
方 法
研究志愿者纳入和样本采集
在2017年8月至2022年8月期间,共338名志愿者被纳入此项研究(图S1)。纳入标准如下:(1)年龄处于18至75岁;(2)预期肠外营养(PN)持续时长大于14天;(3)病情稳定;(4)诊断为CIF。排除标准为:(1)入院前或入院后一个月内使用过抗生素;(2)严重肝肾功能衰竭;(3)妊娠;(4)肠外营养方案有重大变更。在收集临床数据、粪便样本以及血液样本之前,每位参与者均签署了知情同意书,且每位参与者均获得了补偿。本研究已获金陵医院伦理委员会批准(伦理编号:2021NZKY-024-01)。
肠外营养策略
在住院期间,所有志愿者均接受个体化的营养支持治疗。根据志愿者每天住院期间的静息能量消耗、氮水平、血液营养指标以及液体平衡情况,制定个性化的饮食计划并适当调整。
实验动物
C57BL/6小鼠和Rorc-/-小鼠购自江苏集萃药康生物科技股份有限公司(中国南京)。本研究使用无特定病原体条件下饲养的12周龄雄性C57BL/6小鼠。这些小鼠在20~26℃的环境中饲养,光照/黑暗周期为12小时,湿度为40%~70%。所有小鼠在研究开始前均给予食物和水,且活动不受限制。我们使用Powerandsamplesize(https://powerandsamplesize.com/)计算每组小鼠的样本量为10只。
全肠外营养小鼠模型
该造模方法与既往研究中的描述一致。根据不同的干预方案,随机将20只小鼠分为两组,每组10只。小鼠在术后第1天允许饮水,然后分别给予标准饲料(普通饲料组)或全肠外营养支持(TPN组),泵注速度为0.40 mL/h,持续7天后执行安乐死。
菌群移植小鼠模型
该研究涉及两组无特定病原体(SPF)条件下饲养的12周龄雄性C57BL/6小鼠,分别为Chow→Abx组(n = 10)、TPN→Abx(n = 10)。两组小鼠均给予无菌且60Co照射处理的食物,以确保一致的饮食和微生物暴露(1010085;中国上海协同生物)。小鼠移植2周前给予添加混合抗生素的饮用水,其中包括氨苄青霉素、甲硝唑、新霉素(1 g/L)、万古霉素(500 mg/L)和三氯蔗糖(8 g/L)。然后每隔一天给两组小鼠灌胃来自Chow组和TPN组的盲肠内容物稀释液,共4周。盲肠内容物需要称重、合并后与预先稀释的30%甘油/PBS溶液混合,最终浓度为100 mg/mL。然后在厌氧室中通过100 µm尼龙滤器过滤混合液,以去除纤维和大颗粒物质。每次取保存在-80℃下、等量分装0.25 mL稀释后的盲肠内容物进行灌胃。
ICA灌胃小鼠模型
共20只小鼠随机分为两组(每组10只):TPN+PBS组和TPN+ICA组。在开始给予全肠外营养(TPN)前4周,每周小鼠灌胃ICA(10 mg/kg)三次,直至TPN后1周,之后将小鼠安乐死。ICA的剂量是基于之前的研究确定的。在TPN+PBS组中,小鼠灌胃的是PBS对照溶液。
单菌株干预
将30只小鼠随机分为三组(每组10只):TPN+PBS组、TPN+失活L. murinus组和TPN+活L. murinus组。另选取30只小鼠随机分为三组(每组10只)进行ΔArAT L. murinus干预实验:TPN+PBS组、TPN+活L. murinus组和TPN+活ΔArAT L. murinus组。ΔArAT L. murinus由Biosafe Plasmid Bacterial Strain Resource公司提供。MRS Broth(DF0881-17-5;BD Difco,弗林特莱克,新泽西州,美国)中,L. murinus(JCM1717;ATCC,马纳萨斯,弗吉尼亚州)、Lactobacillus reuteri(23272,ATCC,马纳萨斯,弗吉尼亚州)、Lactobacillus johnsonii(33200,ATCC,马纳萨斯,弗吉尼亚州)、Lactobacillus animalis(JCM 5670,ATCC,马纳萨斯,弗吉尼亚州)和ΔArAT L. murinus在37℃下厌氧培养。失活的L. murinus在65℃下孵育15分钟。从TPN前4周开始,直至小鼠安乐死,期间小鼠每天灌胃200 µL含有10-8个菌落的PBS菌悬液。
尤斯灌流实验
从安乐死的小鼠身上获取回肠组织,剪开并用冷的Krebs's溶液(2.5 mmol/L氯化钙、1.98 g/L葡萄糖、4.7 mmol/L氯化钾、1.2 mmol/L磷酸氢二钠、117mmol/L氯化钠、25 mmol/L碳酸氢钠和1.2 mmol/L氯化镁)冲洗三次。组织部分被固定并置于尤斯室(Physiologic Instruments,圣地亚哥,加利福尼亚州,美国)中进行测试,暴露表面积为0.03 cm2。
小肠固有层细胞提取
分离出小肠组织,用生理盐水冲洗掉粪便物质,并用眼科镊和眼科剪切除脂肪组织、派尔氏小结和肠系膜。然后纵向切开,将小肠分成1厘米长的肠段,在用磷酸盐缓冲盐水(PBS)冲洗。最后将肠段转移到含有HEPES缓冲液(10 mM)、DTT(1 mM)和EDTA(30 mM)的洗涤液容器中。在37℃下以200 rpm/min的速度清洗两遍,每次持续20分钟。剩余组织转移到含有消化酶溶液的C管中,在37℃下以150 rpm/min的速度清洗20分钟。将C管放在gentleMACSTM分离器(130-093-235,德国米伦西生物技术公司,伯吉施格拉德巴赫)上获得细胞悬液。将离心重悬后的细胞与Percoll溶液混合,并进行密度梯度离心。随后DMEM重悬中间细胞层。
流式细胞术
将荧光标记染料与单克隆抗体偶联后,用于对单细胞悬液进行染色。在对细胞表面进行染色之前,使用Fixable Viability Dye(65-0865-14,eBioscience,圣地亚哥,加利福尼亚州,美国)来检测活细胞/死细胞。使用鼠源抗体对表面标志物进行染色,如下所示:CD90.2(53-2.1)、CD45(30-F11)、lineage标志物(17A2/RB6-8C5/B220/Ter-119/M1/70)、NK1.1(PK136)和KLRG1(2F1)。上述所有抗体均按照制造商的说明使用。
体外提取的细胞暴露于20 ng/mL的IL-23(200-23,PeproTech,克兰伯里,新泽西州,美国)4小时后进行测试。在使用上述抗体进行表面染色后,使用Foxp3/Transcription Staining Buffer(00-5523-00,eBioscience,圣地亚哥,加利福尼亚州,美国)对靶细胞进行固定和通透处理。然后用IL-22-PerCP eFluor710(IC582N,R&D,明尼阿波利斯,明尼苏达州,美国)和ROR gamma (t)(12-6981-82,Invitrogen,沃尔瑟姆,马萨诸塞州,美国)对细胞进行染色。使用FlowJo软件(版本10.8.1,Tree Star,伍德伯恩,俄勒冈州,美国)对通过Attune NxT 流式细胞仪(BD Biosciences,富兰克林莱克泽,新泽西州,美国)获得的染色细胞进行分析。如图S4A所示,采用流式细胞术分选门控技术来识别ILC3细胞。
ILC3细胞的分离和培养
使用SORP FACSARIA II流式细胞仪分选出纯度为97%的具有Lineage−CD45+CD90.2+NK1.1−KLRG1−表型的细胞(即ILC3细胞)。然后在每孔10,000个细胞的96孔板中加入RPMI-1640培养基,于37℃下培养12小时。培养基中添加了10%的胎牛血清(FBS)、2-巯基乙醇(80 µM)、丙酮酸钠(1 mM)、非必需氨基酸、谷氨酰胺(2 mM)、HEPES缓冲液(10 mM)、100 µg/mL链霉素和100单位/mL青霉素(均购自Gibco,美国马萨诸塞州沃尔瑟姆市)。使用鼠源重组IL-23(40 ng/mL,200-23;PeproTech,美国新泽西州克兰伯里)处理细胞,并以100 µM和200 µM浓度的ICA进行处理。72小时后,检测上清液中的IL-22,随后进行mRNA和流式细胞术分析。
ChIP-qPCR
使用商业化的酶促ChIP试剂盒(53040;Active Motif,加利福尼亚州卡尔斯巴德市,美国)进行了ChIP分析。ILC3核蛋白裂解液在1%甲醛交联后进行了超声处理以破碎染色质,随后用Rorγt抗体(14-6988-82,eBioscience,加利福尼亚州圣地亚哥市,美国)或对照IgG进行免疫沉淀。通过PCR和定量实时PCR(qRT-PCR)对来自输入样品和免疫沉淀样品的目标基因组区域进行扩增。PCR产物在2%琼脂糖凝胶上进行电泳,并使用BG-gdsAUTO550凝胶成像系统(北京贝因基因科技有限公司)进行可视化,然后使用ImageJ进行定量。用于IL-22启动子假定Rorγt结合位点的引物为5'-CTAGTTGTCAGGTGCTATCT-3'(正向)和5'-TGTGCAAGCATAAGTCTCA-3'(反向)。
荧光素酶报告基因检测
构建了包含Rorγt结合序列的IL-22启动子荧光素酶报告质粒Rorγtp-luc,将其插入到pGL3-basic载体(Promega公司,美国威斯康星州麦迪逊市)中。将HEK-293细胞接种于含1%胎牛血清的DMEM培养基的12孔板中,并且不添加抗生素。待细胞达到50%汇合度后,用Rorγtp-luc或Rorγtcp-HA以及作为内对照的Renilla荧光素酶报告质粒进行瞬时转染。转染后24小时,细胞分别暴露于溶媒对照、100 µMICA或200 µMICA中24小时。随后收集细胞并裂解,用于荧光素酶检测。使用双荧光素酶报告基因检测试剂盒(DL101-01,南京维泽迈公司)测定荧光素酶活性,并通过Dual-Glo®荧光素酶检测试剂盒(Promega公司,美国威斯康星州麦迪逊市)进行评估。结果以Renilla荧光素酶水平为参照进行标准化,并以相对倍数变化的形式表示。
提取L. murinus的DNA
使用Tianamp Stool DNA试剂盒(4992205,北京天根生化科技有限公司,中国)按照制造商的说明进行操作从粪便中提取DNA。使用iTaq Universal SYBR(Bio-Rad,美国加利福尼亚州赫尔克斯市;1725125)进行定量PCR(qPCR),使用Fast real-time PCR系统(Thermo Fisher Scientific,美国马萨诸塞州沃尔瑟姆市;7900HT)进行实时PCR(RT-PCR)。L. murinus的引物列于表S2中。
RNA提取和RT-PCR
使用TRIzol LS试剂(15596018CN,Invitrogen公司,美国马萨诸塞州沃尔瑟姆)从小鼠肠道和细胞系中提取总RNA。使用GeneAmp PCR系统9700(Applied Biosystems公司,美国佛罗里达州福斯特城)分析用1µg总RNA逆转录得到的mRNA的相对表达量,使用PrimeScriptTM RT试剂盒(RR037B,Takara公司,日本久留米)进行逆转录。通过2-ΔCt方法比较样本获得的循环阈值(Ct值)。以β-肌动蛋白作为内参检测宿主基因。表S2提供了所有引物。
蛋白印迹实验
从冷冻的小肠组织中提取蛋白质,用SDS-PAGE(4%和8%)将样品转移到聚偏二氟乙烯膜上进行进一步分析。按照制造商的说明使用针对ZO-1(ab59720,Abcam,剑桥,英国)、Occludin(ab216327,Abcam,剑桥,英国)、REG3G(ab239610,Abcam,剑桥,英国)、溶菌酶(ab108508,Abcam,剑桥,英国)和GAPDH(BA3874,BOSTER,圣马特奥,加利福尼亚州,美国)的抗体。使用EPS600电泳仪(坦诺,上海,中国)进行蛋白质印迹,通过Image Lab(版本4.0,Bio-Rad)进行观察,并使用ImageJ(美国国立卫生研究院,贝塞斯达,马里兰州)进行半定量分析;1.52a)。本研究中使用的关键资料列于表S3中。
组织免疫荧光
组织在10%甲醛中固定过夜,然后用石蜡包埋,并切成5毫米厚的切片。从切片上移除石蜡后,使用乙醇和PBS溶液对其进行脱水处理。使用0.2%的Triton X-100溶液(9036-19-5,美国圣路易斯的赛默飞世尔科技公司)进行渗透处理,然后使用10%的正常羊血清(D9663,美国圣路易斯的赛默飞世尔科技公司)进行封闭。在切片上,将一抗在4℃下孵育过夜。应用一抗后,将切片在室温下放置一小时,同时加入相应的二抗。用ProLongTM Gold Antifade试剂(Invitrogen)对切片进行染色处理,并在Zeiss LSM880偏光显微镜下进行观察。
分子对接
使用AutoDock Vina对Rorγt和ICA进行了分子对接研究。该程序利用“迭代局部搜索”算法进行连续的局部搜索,并确定最优的分子对接构象。选择具有最低结合能的解决方案作为最终结果。
苏木精-伊红(HE)染色和Chiu评分
将小鼠肠道分阶段固定,石蜡包埋并切片,放置在载玻片上进行HE染色。染色后,由两名独立的病理学家进行盲法组织学评分,遵循先前描述的标准。根据每组中肠组织切片的一般值对所有个体进行统计分析。
16S rRNA测序
16S rRNA的测序操作与先前研究中所概述的方案类似。后续的数据分析是利用了由杭州开泰生物技术有限公司(中国南宁;网址:https://kaitai.cloud)提供Shengxin云工具这一配套的在线平台进行分析。
mRNA测序
从Chow组和TPN组的小肠组织样本中采集了样本,并使用RNAiso Plus提取总RNA。mRNA测序在NovaSeq™ X Plus,PE150平台(Illumina,圣地亚哥,加利福尼亚州,美国)上进行。使用HISAT2软件(V2.2.1,http://ccb.jhu.edu/software/hisat2)对清洁后的测序数据进行比对,并使用Ensembl数据库(Mus_musculus.GRCm39.109.gtf)进行注释。使用StringTie软件包(https://ccb.jhu.edu/software/stringtie/)进行转录组组装。使用HTSeq(http://htseq.readthedocs.io/en/release_0.9.1)进行定量分析。使用edgeR软件(http://www.bioconductor.org/packages/release/bioc/html/edgeR.html)识别差异表达基因(DEGs)。差异基因选择的阈值设定为|log2FC| > 1且padj < 0.05。我们使用clusterProfiler软件(http://www.bioconductor.org/packages/release/bioc/html/clusterProfiler.html)进行GO/KEGG/Reactome分析。
宏基因组测序实验
使用Illumina HiSeq测序平台对粪便样本的清洁数据进行了生成,并通过Readfq V8进行了处理。使用Bowtie 2.2.4软件过滤了宿主来源的读取数据,该软件将清洁数据与参考数据库(http://bowtiebio.sourceforge.net/bowtie2/index.shtml)进行比对。随后,对清洁数据进行了SOAPdenovo(V2.04)分析(http://soap.genomics.org.cn/soapdenovo.html)。接着,使用SOAPdenovo(V2.04)和MEGAHIT(V1.0.4)进行混合组装。
宏基因组测序统计分析
基于MetaGeneMark软件(网址:http://topaz.gatech.edu/GeneMark/,版本2.10),所有五个Scaftigs都被预测包含开放阅读框(ORF)。采用CD-HIT软件(网址:http://www.bioinformatics.org/cd-hit,V4.5.8)来预测ORF,消除冗余并生成一个独特的初始基因目录。随后使用Bowtie(美国洛杉矶;版本2.2.4)基于每个样本的清洁数据创建了一个初始基因目录,根据特定参数设置确定每个基因的读数数量。随后的分析包括基本统计摘要、相关性分析以及基因计数的维恩图,均使用基因目录中的丰度数据进行。采用DIAMOND软件(网址:https://github.com/bbuchfink/diamond/,版本0.9.9)将unigenes与细菌、病毒、真菌和古菌的序列进行比对。后续的数据分析利用杭开凯泰生物技术有限公司提供的Shengxin云工具(网址:https://kaitai.cloud)进行。
非靶向代谢组学分析
使用配备SCIEX 6500 QTRAP+系统的Orbitrap LC-MS质谱仪对小鼠盲肠内容物中的代谢物进行分析。离子源参数设置如下:Curtain Gas:25psi,Temperature:400°C,IonSpray Voltage:+5500/-4500V,Ion Source Gas 2:60psi,Ion Source Gas 1:55psi,andDP:±100V。使用软件Compound Discoverer(Thermo Fisher Scientific)对代谢物进行定量分析。根据VIP分数超过1.5的代谢物进行显著差异的鉴定,其倍数变化大于2.0或小于0.5且p值小于0.05。使用统计方法对这些代谢物的KEGG通路进行富集分析。使用R版本3.1.3(R软件统计计算,奥地利维也纳)进行数据可视化。
液相色谱-质谱技术(LC-MS)定量分析色氨酸
分离操作采用的是超高效液相色谱(UHPLC)系统(安捷伦1290 Infinity,安捷伦,美国圣克拉拉市),标准样品在自动进样器中于4℃下保存,柱温维持在40℃。定期将质量控制(QC)样品引入样品队列,按照预先设定的实验样品数量之后,以评估系统稳定性和重复性。将与目标分析物相对应的标准物质混合物添加到样品队列中,以帮助校正色谱保留时间。采用SCIEX 5500 QTRAP质谱仪进行质谱分析。使用Multiquant 3.0.2软件提取色谱峰面积和保留时间。通过使用目标化合物的标准物质来校正保留时间,以方便代谢物的鉴定。
单细胞RNA测序实验
在模型建立后的第7天,我们使用SORP FACSARIA II流式细胞仪从Chow小鼠和TPN小鼠中分离出肠道ILCs。使用Single Cell 3'基因表达文库和凝胶微珠试剂盒(V3.1,1000121,10×Genomics)以及Chromium单细胞G芯片试剂盒(1000120,10×Genomics)来捕获细胞并合成cDNA。我们将细胞悬液加载到Chromium单细胞控制器(10×Genomics)上,并在油包水体系中生成单细胞凝胶珠,按照制造商的说明进行操作。每个通道引入约10,000个细胞。单细胞RNA测序(scRNA-Seq)的库制备按照制造商的既定协议执行。使用NovaSeq 6000平台(Illumina)实现了每个细胞至少100,000个读数的深度,双端读长为150个碱基。测序服务由CapitalBio技术(中国北京)提供。
单细胞RNA测序(scRNA-seq)的统计分析
scRNA-seq数据分析由北京CapitalBio技术公司完成,借助了CapitalBio云分析平台。使用Cell Ranger软件(版本7.1.1)来实现数据的比对、计数条形码、过滤以及计数唯一分子标识符(UMI)。此外,还使用R包Seurat3.0进行聚类。基因数少于200个的细胞、基因计数排名前1%的细胞,或者线粒体基因比例超过25%的细胞被归类为异常值,并随后从分析中排除。使用UMAP和t分布随机邻域嵌入(t-SNE)进行可视化。这些分析聚焦于每个聚类的前20个基因标记物。使用R编程包来可视化所得数据。使用Monocle R包构建单细胞轨迹图。利用SingleR(https://bioconductor.org/packages/devel/bioc/html/SingleR.html)对细胞类型进行注释,这是一个能够从scRNA-seq数据中无偏地识别细胞类型的工具。
统计学与可重复性
临床数据通过SPSS(版本25,IBM公司,纽约州阿蒙克市,美国)进行统计分析。实验数据使用Prism 10(GraphPad公司,加利福尼亚州拉霍亚市,美国)进行分析。对于非匹配数据的组间比较,使用Mann-Whitney U检验分析非参数数据,使用Student’s t检验分析正态分布数据。通过Spearman相关性分析评估L. murinus富集度与IL-22水平之间的关系。对于涉及多个组的组间比较,对于非匹配数据使用Kruskal-Wallis H检验,对于正态分布数据使用单因素方差分析(One-way ANOVA),随后使用Dunnett's t检验进行事后校正。双侧p值小于0.05为显著性阈值。
代码和数据可用性
所有测序数据均已存入GSA中的BioProject访问编号PRJCA034984(https://ngdc.cncb.ac.cn/bioproject/browse/PRJCA034984)和PRJCA035609(https://ngdc.cncb.ac.cn/bioproject/browse/PRJCA035609)下。本文中报道的代谢组学数据已存入OMIX,中国科学院北京基因组研究所国家生物信息中心(访问编号:OMIX008699,https://ngdc.cncb.ac.cn/omix/release/OMIX008699和OMIX008700,https://ngdc.cncb.ac.cn/omix/release/OMIX008700)。所使用的数据和脚本保存在GitHub https://github.com/hlc999/HLC_001上。补充材料(图表、表格、图形摘要、幻灯片、视频、中文翻译版本和更新材料)可在在线DOI或iMeta Science网站(http://www.imeta.science/)中找到。
引文格式:
Longchang Huang, Peng Wang, Shuai Liu, Guifang Deng, Xin Qi, Guangming Sun, Xinying Wang, et al. 2025. “Gut microbiota-derived tryptophan metabolites improve total parenteral nutrition-associated infections by regulating Group 3 innate lymphoid cells.” iMeta e70007. http://doi.org/10.1002/imt2.70007.
作者简介

黄龙昌(第一作者)
● 南京大学在读博士研究生。
● 研究方向为肠道菌群及免疫治疗。以第一作者在iMeta、United European Gastroenterology Journal、Frontiers in Nutrition等期刊发表SCI论文3篇。

王鹏(第一作者)
● 佛山市第一人民医院胃肠外科医生,胃肠外科研究所科研助理。
● 研究方向,肠外营养相关并发症,肠道菌群与机体代谢及肠道免疫。广东省医学会肠内肠外营养学分会委员。主持国家自然科学基金青年项目1项,以第一作者(含共同)在iMeta、Nature metabolism,Cellular and Molecular Gastroenterology and Hepatology,Frontiers in Nutrition,Frontiers in Microbiology等国际权威期刊发表文章5篇。

刘帅(第一作者)
● 南京大学在读博士研究生。
● 研究方向为肠道菌群与肠外营养相关疾病。以第一作者(含共同)在iMeta、Medical Science Monitor、Aging等期刊发表SCI论文3篇。

邓桂芳(第一作者)
● 华中科技大学协和深圳医院临床营养科副主任,硕士研究生导师。
● 研究方向为肠外营养肝损伤的机制研究;妊娠期糖尿病胰岛素抵抗的机制研究。主持国家自然科学基金青年项目、广东省自然科学基金、深圳市科创委基础研究面上项目等国家、省市级项目多项。以第一或通讯作者在iMeta、Critical reviews in food science and nutrition、Clinical Nutrition等国际权威期刊发表论文共24篇,其中ESI (Essential Science Indicators) 收录高引用论文2篇。

王新颖(通讯作者)
● 东部战区总医院普通外科副主任、全军普通外科研究所副所长、普通外科临床营养中心主任,博士生导师。
● 研究方向为普通外科重症的临床一线救治及营养治疗。2020年获第十六届中国青年科技奖。军队高层次科技创新人才工程学科拔尖人才、江苏省杰出青年基金获得者、全国临床营养杰出贡献专家(中华医学会)、国之名医·青年新锐(人民日报社)、江苏省“333高层次人才培养工程”第二层次培养对象、江苏省医学重点人才。主持国家公益性行业科研专项项目1项,国家自然科学基金6项,国家、军队和省部级重大科研任务18项,总经费2700万元。研究结果在iMeta、Nature Metabolism、JAMA Surgery、In J Surg和JSCM等国际顶刊发表SCI论文90余篇。作为主要执笔人制定中国临床营养指南/共识6部。主编/副主编国家医学教材《临床营养学》等专著8部,参编15部。研究结果获国家科技进步一等奖、江苏省科技进步一等奖等国家、军队和省部级奖励7项。目前担任中华医学会肠外肠内营养学分会副主任委员、中国医师协会外科医师分会临床营养专家工作组组长、中华医学会外科学分会营养支持学组委员、中华医学会理事会组织工作委员会委员、中国营养学会医用食品与营养支持分会常务委员、国家食品药品监督管理局药品审评专家,军队后勤科技装备评价专家库专家,江苏省外科学分会营养外科学组组长。担任《肠外与肠内营养》杂志副主编,《中华临床营养杂志》《中华胃肠外科杂志》和Clinical Nutrition及Science Reports编委。
更多推荐
(▼ 点击跳转)
iMeta | 引用16000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 兰大张东组:使用PhyloSuite进行分子系统发育及系统发育树的统计分析
iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVI
iMeta封面

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

3卷2期

3卷3期

3卷4期

3卷5期

3卷6期

4卷1期
iMetaOmics封面

1卷1期

1卷2期

2卷1期
期刊简介
“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述,重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊!相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录!2024年6月获得首个影响因子23.8,位列全球SCI期刊前千分之五(107/21848),微生物学科2/161,仅低于Nature Reviews,学科研究类期刊全球第一,中国大陆11/514!
“iMetaOmics” 是“iMeta” 子刊,主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任,是定位IF>10的高水平综合期刊,欢迎投稿!
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science

















 3591
3591

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









