








草本植物的种子或颖果起源于一个单心皮的子房,其中只有一个胚珠,并包含主要的储藏组织——胚乳。对于大多数草本作物(即谷物),作物的价值很大程度上由胚乳决定,无论是在数量上还是在质量上。
胚乳是由胚囊中央细胞中的两个极核与一个精子细胞核受精产生的,这产生了一个三倍体(3n,3C)的核,而二倍体(2n,2C)的胚则起源于第二个精子细胞核与卵细胞的受精。胚乳的主要功能是为发育中的以及后来萌发的胚提供营养物质。与包括拟南芥(Arabidopsis thaliana)在内的许多物种不同,草本植物的胚乳是一个持久的种子结构。它是人类和牲畜营养的主要热量来源,并为无数制造食品、商品和生物燃料提供原材料。
尽管草本植物胚乳的重要性,但在许多作物种类中,其发育尚未得到彻底研究,更不用说非作物种类了。在草类植物中,胚乳的发育具有相当的一致性,特别是在早期阶段(Weatherwax,1930;Bennett等人,1975)。尽管已知存在偏差,但这些通常涉及发育的次要方面。通常,大多数草类植物的胚乳在成熟时是淀粉质和干燥的,这当然是一个有价值的特性,但也有例外。例如,在对169个草类属(占该科总属数的25%以上)的调查中,发现有30个属在成熟时具有液态或软质胚乳的物种,胚乳的粘性状态可以保持几十年。另外9个属被发现具有半固态胚乳(Terrell,1971)。
在草类植物中,胚乳的发育在玉米(Zea mays)中得到了最好的描述,这是由于历史、经济、解剖和遗传原因;因此,我们将主要参考从这种谷物获得的知识,作为草本植物胚乳发育的范例。在适当和可能的情况下,我们将突出和讨论玉米和其他草类物种之间的差异。在这里,我们提供了胚乳发育阶段的概述,包括遗传、分子和细胞调控机制的独特特征。有兴趣深入了解草类植物胚乳发育不同方面的读者可以参考几项以前的工作(Kiesselbach,1949;Bennett等人,1975;Kowles和Phillips,1988;Lopes和Larkins,1993;Olsen等人,1999;Becraft,2001;Larkins等人,2001;Olsen,2001,2004,2007;Sabelli等人,2005b,2007)。对突变体的分析为胚乳发育的调控提供了大量知识,相关文献非常广泛。我们不会回顾每一个突变,而是将讨论我们认为提供关键见解的选定突变。有许多出版物包含了关于影响谷物胚乳发育的突变的丰富信息(Jarvi和Eslick,1975;Nelson,1980;Neuffer和Sheridan,1980;Satoh和Omura,1981;Bosnes等人,1987;Kowles等人,1992;Scanlon等人,1994;Kurata等人,2005;Dolfini等人,2007)。
禾本科植物胚乳发育:主要事件
禾本科植物胚乳的发育有多个不同的阶段,这些阶段可能会有相当大的重叠(见图1)。这些阶段分别如下:早期发育,包括双受精、合胞体形成和细胞化;分化阶段,包括主要细胞类型(转移细胞、糊粉层细胞、淀粉胚乳细胞和胚胎周围细胞)的形成,有丝分裂和内复制的时期,以及贮藏物质的积累;以及成熟阶段,包括程序性细胞死亡(PCD)、休眠和脱水。
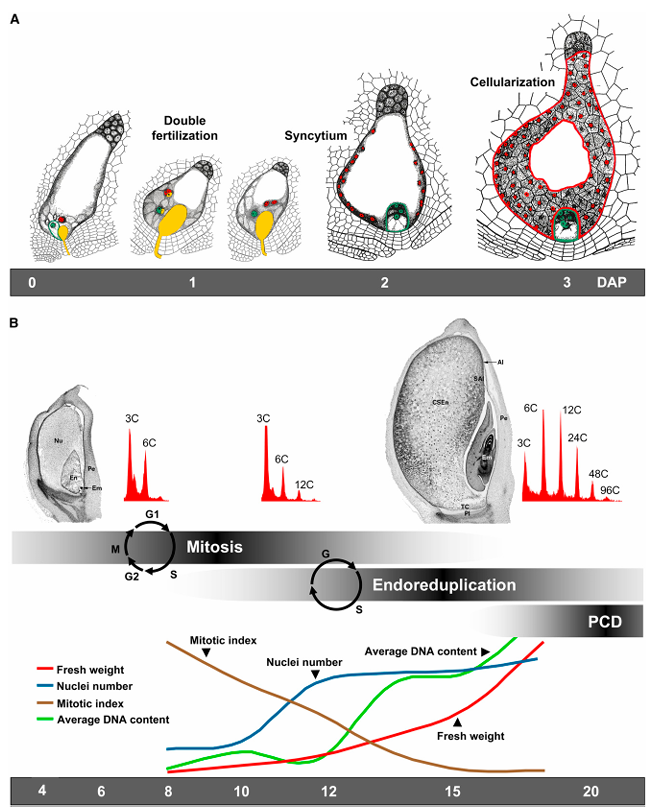
图1. 胚乳发育阶段。虽然这个图示指的是玉米,但它也是其他禾本科植物胚乳发育的良好示例。
A. 在受精后3到4天(DAP,即天数后受精)内发生双受精、合胞体形成和胚乳细胞化。花粉管和精子核以黄色显示,女性配子体中心细胞的极核和胚乳核以红色显示,卵细胞核和胚胎核则以绿色显示。图中还展示了胚乳发育的不同阶段的轮廓。
早期发育
双受精
在许多物种中,包括玉米(Kiesselbach,1949年)、小麦(Triticum aestivum;Bennett等人,1975年)、薏苡(Coix lacryma-jobi;Weatherwax,1930年)、高梁(Paspalum scrobiculatum;Narayanaswami,1954年)和中国结缕草(Eragrostis unioloides;Deshpande,1976年),单倍体(一个精子核与卵细胞核的融合)和双倍体(一个精子核与两个极核的融合以形成胚乳)是同时发生的。有趣的是,三倍体初级胚乳核的有丝分裂活动非常迅速,而在合子中的这一过程则经历了一个明显的暂停。最近在拟南芥中的发现表明,中央细胞的增殖需要来自受精卵细胞的积极信号(Nowack等人,2006年),这意味着极核被“预处理”以进行更快的细胞周期活动。是否在禾本科家族中存在类似的机制尚不清楚。在高梁和甘蔗(Saccharum officinarum;Narayanaswami,1954年)以及不定胚乳玉米突变体(Lin,1978年)中,受精前存在多余的极核,这表明存在一个潜在的途径,可能导致早期细胞周期活动和过早的胚乳增殖,这些在正常情况下通常被抑制。
合胞体形成
禾本科植物的胚乳遵循一种常见的胚乳发育模式,即核型(或合胞体型)(Lopes和Larkins,1993;Olsen,2004),在这种模式中,最初的三倍体核在没有细胞壁形成和细胞质分裂的情况下经历几轮通常是同步的分裂,导致合胞体的形成。极核的受精产生了初级胚乳核,它在数小时内开始迅速分裂。如前所述,合子中的细胞分裂总是较慢。例如,在玉米中,当合子第一次分裂时,已经有四到八个胚乳核(Randolph,1936),在高梁中多达24个胚乳核(Narayanaswami,1954),以及中国结缕草中有大量的胚乳核(Deshpande,1976)。当胚胎细胞开始增殖时,它们的分裂速度比胚乳核慢(与分生组织细胞的分裂速度相当),很可能是因为胚乳核的增殖不涉及细胞质、细胞膜和细胞壁的合成。观察到胚乳增殖速度的下降(大约等于胚胎细胞的速度)与合胞体的细胞化同时发生,这支持了这种解释(Bennett等人,1975年)。实际上,合胞体胚乳的细胞周期通常缺乏子核间形成细胞板的过程,这加强了这样一种观点,即负责在体细胞中创建细胞骨架某些部分的程序被抑制了。因此,合胞体胚乳发育可以被视为一种进化策略,通过抑制控制细胞质分裂和细胞壁形成的程序,迅速填充中央细胞的大而预先形成的细胞质,并达到更大的基础细胞数量,以支持这种组织的增长,并为胚胎的发育做好准备,特别是在胚乳中专门用于营养吸收的特定细胞尚未分化的时期。母源的胚乳和反足细胞可能通过氨基酸、核苷酸和碳水化合物支持合胞体胚乳的生长(Bennett等人,1975年;Radchuk等人,2006年)。

















 642
642

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









