





大脑的内在功能组织在进入老年期时会发生变化。年龄差异在多个空间尺度上都能观察到,从分布式大脑系统的模块性和分离程度的全局性减少,到特定网络的去分化模式。目前尚不确定去分化是否反映了随年龄增长而不可避免的大脑功能全局性转变,局限的、依赖经验的变化,或两者兼而有之。我们采用了多方法策略来探究多个空间尺度上的去分化现象。我们对年轻(n = 181)和年长(n = 120)的健康成年人收集了多回波(ME)静息态fMRI数据。我们实施了对个体差异敏感的皮层分区方法,以精确绘制每个参与者的功能图谱,同时保留组水平的分区和网络标签。ME-fMRI处理和梯度映射识别出了全局和宏观尺度的网络差异。多变量功能连接方法用于测试微观尺度、边水平的差异。老年人的BOLD信号维度较低,这与全局网络去分化一致。梯度在很大程度上不随年龄变化。边水平分析揭示了老年人离散的、特定网络的去分化模式。视觉和体感区域在功能连接组中更加整合;默认模式网络和额顶控制网络区域显示出更强的连接性;背侧注意网络与异质区域更加整合。这些发现突出了多尺度、多方法方法在描述功能性大脑老化架构中的重要性。本文发表在Cerebral Cortex杂志。
关键词:连接性、梯度、分区、网络神经科学、多回波fMRI
引言
大脑活动的自发振荡为表征大尺度功能网络提供了基础(Fox and Raichle 2007; Biswal et al. 2010; Yeo et al. 2011)。这种内在的功能网络架构由遗传因素和经验依赖的神经可塑性变化共同决定,这些变化跨越了从瞬时到数十年的时间尺度(Stevens and Spreng 2014)。老化连接组的关键组织特征包括网络内连接减少和网络间连接增加(Chan et al. 2014; Geerligs et al. 2015),导致了一个去分化或更少分离的网络架构(Wig 2017)。
现在有大量证据表明,内在网络去分化是功能性大脑老化的一个全局特征(Chan et al. 2014; Geerligs et al. 2015; Stumme et al. 2020,另见Damoiseaux 2017; Wig 2017的综述)。这可能反映了大脑对随年龄增长而发生的系统性结构、神经生理、血管或代谢改变的功能重组(Reuter-Lorenz and Park 2014; Kantarovich et al. 2022),与特定脑区功能专门化的丧失相似(Cabeza et al. 2002; Park et al. 2004; Rajah and D'Esposito 2005)。这些全局网络变化表明,网络去分化可能是一个非特异性的神经标记,与晚年大脑健康下降的其他总体指标相似。然而,这种非特异性变化也可能反映了数据获取、处理和分析静息态fMRI数据的方法中与年龄相关的系统性混淆因素(见Liem et al. 2021的综述)。其中最突出的是与年龄相关的运动差异(D'Esposito et al. 1999; Geerligs et al. 2015),它引入了虚假的与年龄相关的连接模式差异(Power et al. 2012, 2014)。这些和其他混淆因素突显了对静息态fMRI数据进行严格去噪的需求,并给该领域带来了重大挑战(Power et al. 2018; Spreng et al. 2019)。
与全局网络变化相反,越来越多的证据表明存在特定网络的去分化模式。在特定的联合网络中(Betzel et al. 2014; Keller et al. 2015; Ferreira et al. 2016; Ng et al. 2016; Rieck et al. 2017; Spreng et al. 2018; Zonneveld et al. 2019; Malagurski et al. 2020)以及联合网络和感觉运动网络之间(Meier et al. 2012; Chan et al. 2014; Song et al. 2014; Seidler et al. 2015; King et al. 2018; Manza et al. 2020; Stumme et al. 2020)都报告了与年龄相关的去分化。随年龄增长,最常观察到的特定网络去分化模式之一是默认网络和背侧注意网络之间BOLD信号反相关的减弱(Geerligs et al. 2015; Ferreira et al. 2016; Spreng et al. 2016)。这些经典的大脑网络在年轻成年人的静息状态和大多数任务中表现出强烈的反相关(Fox et al. 2005; Toro et al. 2008; 但请参见Dixon et al. 2017)。
特定网络之间更大整合的证据暗示了静息态功能连接(RSFC) fMRI在老化大脑健康全局指标之外的作用。相反,这些特定的去分化模式可能作为晚年特定领域认知变化的神经标记。事实上,可分离的网络去分化模式已经与年龄在视空间能力(Manza et al. 2020)、运动功能(King et al. 2018)、情景记忆(Andrews-Hanna et al. 2007; Chan et al. 2014; Spreng et al. 2018)、处理速度(Geerligs et al. 2015; Ng et al. 2016; Malagurski et al. 2020)和执行功能(Keller et al. 2015; Stumme et al. 2020)方面的差异相关联。Chan等人(2014)提供了早期证据,表明特定网络的去分化模式是特定领域神经认知老化的标记。他们观察到与较低的情景记忆能力相关的全局网络去分化模式(即减少的分离)。然而,这种关系在联合网络中比在感觉运动网络中更强,表明皮层联合网络中的去分化既是年龄相关情景记忆下降的敏感标记,也是特异性标记。我们报告了类似的发现,显示年龄相关的默认网络和额叶区域之间连接性增加与较低的流体认知相关,以及老年人个人过去回忆的性质和内容的特定差异(Spreng et al. 2018)。
这些报告(和众多其他报告)表明,大脑内在网络组织的变化可以提供神经认知老化的全局和特定网络标记。然而,RSFC研究之间理论、实证和方法论差异的范围,以及众多分析挑战,阻碍了对网络组织年龄相关差异的精确映射。这种精确性对于开发神经认知老化的敏感和特异性RSFC标记至关重要。在此,我们提出了一项综合的、跨方法的研究,探究年轻人和老年人样本中网络去分化的模式。我们采用了三种方法来调查从全局到边水平年龄对比的多个空间尺度上的去分化,同时试图缓解几个最常见的fMRI获取和分析挑战。我们在下面简要描述每种方法及其相关假设。图1展示了我们分析方法的概览示意图。具体的方法细节包含在各自的材料和方法部分。
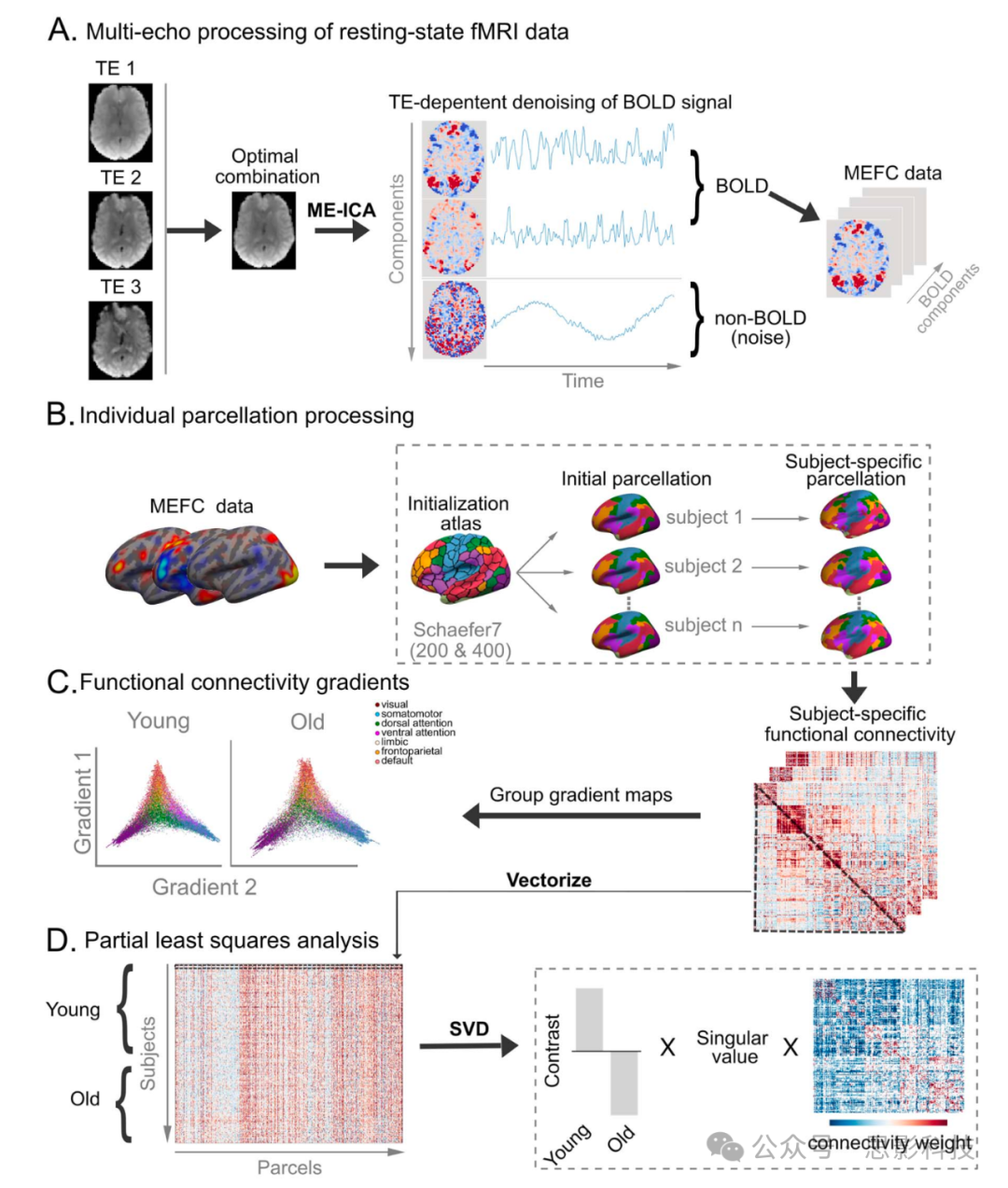
图1 研究方法工作流程。
a) 多回波静息态fMRI图像的处理。对于每次功能扫描,三个回波(TE1, TE2和TE3)被结合并使用ME-ICA进行去噪。去噪过程涉及移除非BOLD信号(噪音)成分并保留BOLD成分。MEFC图像由BOLD成分系数集组成。
b) 生成个体化分区。所有参与者的MEFC数据被重采样到一个共同的皮层表面。所有参与者首先初始化为预定义的皮层分区图谱(Schaefer图谱)。然后按参与者(特定主体分区)细化分区。对于每个参与者,从每个分区提取MEFC数据并与之相关,以创建特定主体的功能连接矩阵。
这些矩阵用于c) 计算年轻人和老年人的皮层梯度,以及d) 使用PLS评估功能连接的年龄相关差异,PLS执行奇异值分解(SVD)。
我们首先通过测量皮层BOLD信号协方差的时空模式差异来检查全局网络去分化,我们将这一测量称为BOLD维度性。多回波fMRI(ME-fMRI)数据采集协议的创新,结合使用多回波独立成分分析(ME-ICA)的TE依赖模型进行BOLD信号去噪,能够可靠地将BOLD信号从非BOLD(即噪音)信号分离成不同的成分(Kundu et al. 2017)。新兴的证据表明,BOLD成分的数量,或BOLD维度性,是一个具有生物学意义的指标,显示从儿童到中年期的下降反映了更大的功能整合和空间分布和分离的大尺度网络的发展(Kundu et al. 2018)。虽然这个指标此前尚未在老年人中进行过检验,但我们预测BOLD维度性作为全局网络分化的代表,在老年人中会显著低于年轻人,这与先前报告的年龄相关网络去分化一致(Betzel et al. 2014; Chan et al. 2014; Geerligs et al. 2015; Ng et al. 2016; Zonneveld et al. 2019; Madden et al. 2020; Malagurski et al. 2020; Stumme et al. 2020)。
其次,我们使用扩散图嵌入方法来估计RSFC皮层梯度,研究了宏观尺度水平上的网络去分化的年龄差异(Margulies et al. 2016; Huntenburg et al. 2018; Paquola et al. 2019; Vos de Wael et al. 2020)。梯度映射识别描述皮层表面区域连接模式转变的特征向量,主要RSFC特征向量解决方案通常区分感觉/运动皮层和跨模态皮层(Margulies et al. 2016)。由于梯度是连接组的稳健组织特征,我们预测这些模式在很大程度上能抵抗正常的年龄相关变化。然而,在宏观尺度梯度架构内可能会出现区域特定连接概况的变化,反映在不随年龄变化的宏观尺度网络组织中发生的特定网络或节点转变(Bethlehem et al. 2020)。
第三,我们使用偏最小二乘(PLS)分析检查了未阈值化的边水平连接组学识别的去分化模式的年龄差异(Krishnan et al. 2011; McIntosh and Mišić 2013)。PLS是一种多变量方法,可以在单个统计步骤中分析完整的边水平连接矩阵,消除了在预先定义的网络分区方案中进行额外阈值化的需要,使我们能够在整个矩阵中识别可靠的年龄差异。这为检测功能脑网络组织的年龄差异提供了边水平的精确度,包括特定网络的去分化模式。我们首先在经典的七网络解决方案(Yeo et al. 2011)中检查了边水平连接组学。我们预测网络内连接减少而网络间连接增加,这与先前报告和上述综述一致。由于我们不知道有任何先前研究报告未阈值化连接的统计可靠模式,我们还预计这种增加的边际分辨率将揭示迄今未报告的年龄差异。最后,基于先前的工作(Spreng et al. 2013, 2016; Grady et al. 2016),我们对默认、背侧注意和额顶控制网络的子网络拓扑进行了先验分析,这些网络源自Yeo等人(2011)的17网络解决方案。在这里,我们预测随年龄增长网络内连接性降低,默认和背侧注意网络之间的反相关减弱,以及额顶控制网络与默认和背侧注意网络区域之间的网络间连接性增强。
综合起来,这些技术为人类大脑的功能架构提供了广泛的视角,从皮层的全局协方差模式到边水平连接的精确映射。通过在认知正常的老年人和年轻人的大样本中实施综合的、多尺度分析方法,我们旨在描述老化大脑内在功能架构的全局和特定年龄差异模式。
材料和方法
参与者
参与者包括181名年轻人(平均年龄 = 22.59岁,标准差 = 3.27;57%为女性)和120名老年人(平均年龄 = 68.63岁,标准差 = 6.44;55%为女性),他们来自纽约州伊萨卡和加拿大多伦多(表1),总样本量为301人。实施了标准的纳入和排除标准,以确保所有参与者都是健康的,没有已知影响大脑或认知功能的神经系统、精神病或其他潜在医疗条件的证据。具体而言,筛选排除了患有急性或慢性精神疾病的个体。还询问了参与者目前是否使用情绪(如抑郁)、思维或心理能力(如注意力缺陷障碍)的药物,或在资格面试前3个月内是否经历了显著的健康状况变化。年轻和老年参与者分别使用贝克抑郁量表(Beck et al. 1996)或老年抑郁量表(Yesavage and Brink 1983)筛查抑郁症状。由于被评为"中度抑郁",排除了两名老年人。为了筛查正常认知功能,对参与者进行了简易智能状态检查(MMSE; Folstein et al. 1975; 年轻人平均分: 29.1; 年轻人标准差: 1.2; 老年人平均分: 28.6; 老年人标准差: 1.3)和NIH认知测试(Gershon et al. 2013)。如果参与者在MMSE上得分低于27/30,并且在NIH的流体认知指数年龄调整分数中得分低于第25百分位(Hackett et al. 2018; Scott et al. 2019),则被排除。所有参与者都是右利手,视力正常或经矫正后正常。研究程序遵守康奈尔大学机构审查委员会和约克大学研究伦理委员会的规定,包括从每位研究参与者那里获得书面知情同意。
注:情景记忆、语义记忆和执行功能是指数得分。处理速度是符号数字模态任务(口头版)的z分数。
表1. 人口统计样本
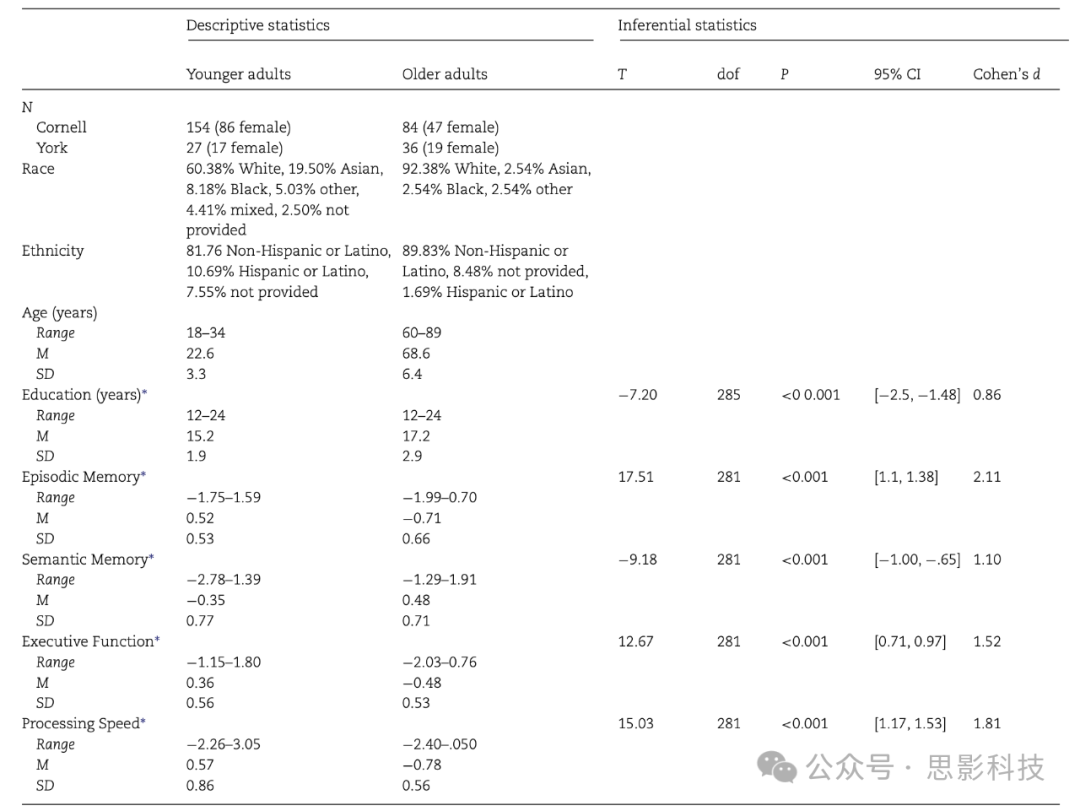
*表示组间存在显著差异。14名参与者未记录教育程度。在283名参与者中测试了情景记忆、语义记忆、执行功能和处理速度的年龄组差异。正T值反映年轻人得分更高,而负值反映老年人得分更高。在ANCOVA中加入性别、教育程度、地点和估计全脑体积(eWBV)作为协变量时,统计结果几乎相同。
认知评估
我们首先通过全面的认知评估对样本进行了表征;301名个体中的283名(163/181名年轻成年人,120/120名老年人)在大脑扫描前进行了认知测试。为情景记忆、语义记忆、执行功能和处理速度的认知领域创建了指数得分(描述性统计见表1)。情景记忆任务包括韦氏记忆量表-IV的言语配对联想(Wechsler 2009)、联想回忆范式(Brainerd et al. 2014)以及NIH认知工具箱的雷伊听觉言语学习和图片序列记忆测试(Gershon et al. 2013)。语义记忆任务包括Shipley-2词汇测试(Shipley et al. 2009)和NIH认知工具箱的图片词汇和口头阅读识别测试(Gershon et al. 2013)。执行功能指数包括连线测试(B-A;Reitan 1958)、阅读广度任务(Daneman and Carpenter 1980)、NIH认知工具箱的Flanker抑制控制和注意任务、维度变换卡片分类以及列表排序工作记忆测试(Gershon et al. 2013)。处理速度通过符号数字模态测试口头版(Smith 1982)进行测试。
所有数据都进行了z分数转换。指数得分代表一个认知领域内所有测量的平均z分数。在4个领域中,更高的分数代表更好的表现。在0.05的显著性水平下进行了脑-行为积差相关分析,并计算了95%置信区间(CI)。对4个指数得分测试进行了Bonferroni多重比较校正,显著性水平设为P < 0.013。
神经影像学
图像采集
成像数据在康奈尔大学磁共振成像设施的3T GE750 Discovery系列MRI扫描仪上使用32通道头线圈采集,或在多伦多约克大学神经影像中心的3T西门子Tim Trio MRI扫描仪上使用32通道头线圈采集。扫描方案在不同地点之间进行了密切匹配。在康奈尔大学,解剖扫描使用T1加权容积磁化准备快速梯度回波序列(TR = 2530 ms;TE = 3.4 ms;7°翻转角;1 mm等向体素,176层,5分25秒)采集,采用2倍加速和灵敏度编码。在约克大学,解剖扫描使用T1加权容积磁化准备快速梯度回波序列(TR = 1900 ms;TE = 2.52 ms;9°翻转角;1 mm等向体素,192层,4分26秒)采集,采用2倍加速和广义自校准部分并行采集(GRAPPA)编码,iPAT加速因子为2。在康奈尔大学(TR = 3000 ms;TE1 = 13.7 ms,TE2 = 30 ms,TE3 = 47 ms;83°翻转角;矩阵大小 = 72 × 72;视野(FOV)= 210 mm;46个轴向切片;3 mm等向体素;204个体积(volume),2.5倍加速和灵敏度编码)和约克大学(TR = 3000 ms;TE1 = 14 ms,TE2 = 29.96 ms,TE3 = 45.92 ms;83°翻转角;矩阵大小 = 64 × 64;FOV = 216 mm;43个轴向切片;3.4 × 3.4 × 3 mm体素;200个体积(volume),3倍加速和GRAPPA编码)采集了两次10分06秒的静息态扫描,使用多回波EPI序列。要求参与者保持清醒,在昏暗的扫描室中静卧,眼睛睁开,正常呼吸和眨眼。
图像处理
使用FSL BET(Smith 2002)的默认参数对解剖图像进行了颅骨剥离。将经过颅骨剥离的解剖和功能图像提交给ME-ICA(版本3.2 beta;https://github.com/ME-ICA/me-ica;Kundu et al. 2011, 2013)。ME-ICA依赖于BOLD信号的TE依赖模型来确定每个体素中的T2*,并将BOLD信号与非BOLD噪声源分离。在进行TE依赖去噪之前,对时间序列数据进行了最小预处理:丢弃前4个体积(volume),计算去斜、运动校正和解剖-功能配准的图像,并将体积在TE之间进行空间对齐。然后使用T2*图进行解剖-功能配准。T2*图比原始EPI图像更精确地描绘了灰质和脑脊液隔室(Speck et al. 2001;Kundu et al. 2017),这在老化研究中是一个重要考虑因素,因为这些边界常常因扩大的脑室和更大的蛛网膜下腔而变得模糊。随后,在TE之间进行最优组合和去噪。感兴趣的输出包括:(i) 由BOLD成分组成的空间图,(ii) 仅包含BOLD成分的重建时间序列,以及(iii) BOLD成分系数集。
在本地空间对去噪后的时间序列进行图像质量评估,以识别和排除配准不成功、残余噪声(帧间位移 > 0.50 mm且去噪后时间序列显示DVARS > 1,Power et al. 2012)、时间信噪比 < 50或保留的BOLD类成分少于10个的参与者(见补充图1的组时间信噪比图)。
在后续步骤中使用了本地空间中经过去噪的BOLD成分系数集,这些系数集针对功能连接分析进行了优化(Kundu et al. 2013)。我们将这些称为多回波功能连接(MEFC)数据。如下所述,在估算连接矩阵后,采取了额外措施来解释ME-ICA独立成分数量的变化。使用FreeSurfer v6.0.1(Fischl 2012)将MEFC神经图像映射到每个参与者的共同皮层表面。为了最大化结构和功能数据强度梯度之间的对齐(Greve and Fischl 2009),首先将MEFC数据按运行线性配准到T1加权图像。使用这个配准的逆变换将T1加权图像投影到本地空间,并使用三线性体积到表面插值将MEFC数据重采样到皮层表面(fsaverage5)。这产生了一个皮层表面图,其中每个顶点或表面点都是从体素数据插值得到的。一旦在表面上,运行被串联,每个顶点的MEFC数据被归一化为零均值和单位方差。
个体化分区
经ME-fMRI处理的数据提供了优秀的可靠性和时间信噪比,足以进行个体精确映射(Lynch et al. 2020, 2021)。实施了个体化功能分区方法来识别个人特定的功能网络节点(Chong et al. 2017)。这些个体化分区在梯度和边水平连接分析中都被使用,以便于比较年轻人和老年人之间的RSFC。对标准化模板的不良配准可能无法捕捉皮层功能组织的个体差异,这些配准问题可能在不同年龄组之间系统性地不同(Laumann et al. 2015; Wang et al. 2015; Braga and Buckner 2017; Chong et al. 2017; Gordon et al. 2017; Kong et al. 2019, 2021)。派生功能定义的、个人特定的皮层分区可以解释个体水平的差异,从而减轻组间比较中的系统性配准偏差。采用个体化分区方法还可能减少噪音伪影的影响,这些伪影可能掩盖小但可靠的组间差异,增加检测可靠脑-行为关联的能力(Kong et al. 2021)。
我们使用群体先验个体分区(GPIP; Chong et al. 2017)生成了参与者特定的功能分区,以检查功能性脑网络组织的个体差异。这种方法能够更准确地估计参与者特定的个体功能区域(Chong et al. 2017),并且对RSFC与行为的关联更敏感(如Mwilambwe-Tshilobo et al. 2019)。这种方法的主要优势是在参与者之间保持了分区标签的对应关系,同时允许分区边界根据每个参与者特定的功能网络组织进行移动——从而提供了一个在人群中共享的连接模式。从初始预定义的群体分区图谱开始,GPIP首先根据每个个体的静息态fMRI数据细化其分区边界。然后,使用群体稀疏约束联合估计所有受试者的浓度(逆协方差/偏相关)矩阵。GPIP在这两个步骤之间迭代,持续更新分区标签直到收敛,定义为每个分区不超过一个顶点改变或20次迭代。与其他基于群体的分区方法相比,GPIP已显示可以改善分区内BOLD信号的同质性和功能专门化区域之间的界限(Chong et al. 2017)。
我们从每个顶点提取MEFC数据,并在200和400个分区的分辨率下对全部301名参与者应用上述分区。对于每种分辨率,MEFC数据被初始化为Schaefer等人(2018)开发的群体分区图谱。我们使用这种皮层分区方案作为我们个体化皮层分区图的初始化参考,以及下面描述的边水平连接组学分析。我们选择Schaefer图谱有三个原因:(i)它是功能派生的,因此与当前研究目的更紧密一致,(ii)它对不同粒度水平具有高空间分辨率(我们在这里报告400和200节点分区的发现),(iii)它是文献中最常用的皮层分区图谱之一,提供Yeo 7网络和17网络解决方案内节点的内在分区(在我们的边水平连接组学分析中使用;Yeo et al. 2011)。
在用Schaefer分区初始化后,重复两步迭代过程20次,以产生代表整个皮层表面最优分区的最终分区。我们通过取给定分区中所有顶点对的平均相关系数,然后在所有分区中平均来计算同质性。在每次重复时都重复这个过程,以观察迭代分区过程中同质性的增量变化。首先在参与者水平计算同质性,然后在整个群体中平均以得到群体估计。对于一部分参与者,最终分区中的一些分区合并到内侧壁(没有数据存在的地方)或属于对侧半球的分区中。由于分区可能反映参与者特定的功能组织神经生物学变异,分配给对侧半球的分区被允许保留其原始群体图谱标签。在400分区分辨率下,69名老年人和35名年轻人出现了与内侧壁合并的分区,这些参与者平均2-3个分区;62名老年人和24名年轻人出现了迁移到对侧半球的分区,平均2-3个分区。在200分区分辨率下,18名老年人和10名年轻人出现了与内侧壁合并的分区,这些参与者平均1个分区。在这个分辨率下没有分区迁移到对侧半球。
功能连接矩阵
根据每个参与者的个体化分区解决方案构建了一个连接矩阵。由于MEFC数据由ICA系数集(每个接受的成分×顶点的系数权重)按运行连接而成,我们从每个分区内的顶点提取并平均MEFC数据,以获得分区级别的系数集。通过计算每个分区系数集之间的积矩相关来估计连接性,从而得到一个n个分区 × n个分区的功能连接矩阵(Ge et al. 2017)。在这种方法中,RSFC被计算为跨分区的ICA系数的相关性,而不是通常所做的BOLD信号时间序列的相关性(见Kundu et al. 2013)。然后应用经典的Fisher's r-to-z转换来标准化相关值的分布,并解释MEFC数据自由度(即去噪ICA系数的数量,即BOLD成分的数量)在个体间的变化(Kundu et al. 2013)。
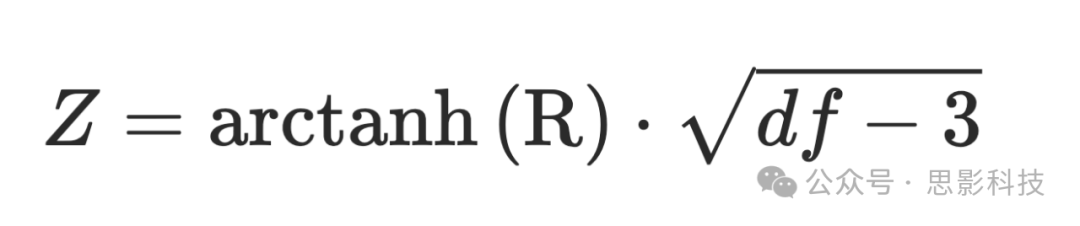
其中R是积矩相关值,df是去噪ICA系数的数量。使用近似独立的成分系数计算功能连接使得全局信号回归变得不必要(Spreng et al. 2019)。重要的是,ME-ICA有效地从fMRI数据中去除了距离依赖的RSFC运动混淆(Power et al. 2018)。如补充图2所示(另见补充材料),帧间位移对年轻人和老年人RSFC的影响相当,排除了运动作为下面报告的结果中的潜在混淆因素。
分析
BOLD维度性
ME-fMRI和ME-ICA处理框架的一个独特优势是BOLD和非BOLD信号被分离成独立的成分。然后可以检查ME-ICA识别的BOLD成分数量这一新的"BOLD维度性"指标,作为大脑全局网络架构的数据驱动表示,并用于研究随年龄变化的情况(如Kundu et al. 2018)。我们评估了BOLD维度性在2次数据运行中的测试-重测可靠性。然后使用独立样本t检验和ANCOVA比较了年龄组之间的总BOLD维度性,控制了性别、教育、地点和估计全脑体积(eWBV;由FreeSurfer得出的灰质和白质总和除以总颅内体积)。为了观察BOLD维度性随年龄增长在整个生命周期的轨迹,将来自独立发展样本的BOLD维度性数据(n = 51,10名女性;平均年龄 = 21.9岁;年龄范围,8.3-46.2岁;详见Kundu et al. 2018)与当前数据合并。由于我们的样本由2个离散的年龄群组成,这些额外的数据点被用来正确拟合年龄和BOLD维度性之间的函数。为了使样本具有可比性并解释跨数据集采集的差异,BOLD维度性按获取的时间点数量进行了缩放。然后将年龄和BOLD维度性之间的关系拟合为幂律函数(未缩放版本见补充图3)。BOLD信号维度性的进一步表征,包括与参与系数、模块性和分离的图论分析度量的关联,以及BOLD信号维度性与全脑RSFC的关系,在补充材料中报告(补充表1和补充图4和5)。
梯度和流形离心率
皮层梯度允许功能连接的低维(即宏观尺度)表示,划分全脑功能连接的转变(Margulies et al. 2016; Huntenburg et al. 2018)。梯度图嵌入已可靠地划分了从单模态到异质模态皮层、视觉到体感运动皮层等皮层转变(Margulies et al. 2016; Bethlehem et al. 2020; Hong et al. 2020)。使用BrainSpace工具箱(https://github.com/MICA-MNI/BrainSpace; Vos de Wael et al. 2020)中的函数计算皮层梯度,在MATLAB中实现。对于每个参与者,400 × 400 GPIP功能连接矩阵按行阈值化为上10%的连接,以仅保留最强的正连接(Margulies et al. 2016; Hong et al. 2019)。在稀疏矩阵上计算余弦相似度,作为下面使用的扩散图嵌入算法的输入,生成一个捕捉顶点之间全脑连接模式相似性的矩阵(Margulies et al. 2016; Hong et al. 2019)。
然后我们应用了扩散图嵌入,这是一种来自图拉普拉斯家族的非线性维度流形学习技术(Coifman et al. 2005),以识别个体参与者水平的梯度成分。每个梯度代表从高维相似矩阵估计的低维嵌入/特征向量。在嵌入空间中,全脑功能连接模式相似性较高的顶点出现得更靠近,而不相似的顶点则相距较远。因此,每个嵌入轴可以解释为基于连接模式相似性/差异性的方差轴。嵌入空间中的欧几里得距离等同于以这些点为中心的概率分布之间的扩散距离,每个距离等同于"梯度差异"分数。该算法由单个参数α控制,它控制采样点在流形上密度的影响(Margulies et al. 2016)。我们在本研究中使用α = 0.5,这将扩散图嵌入与拉普拉斯特征图区分开来,并允许在嵌入空间的估计中包括全局和局部关系。进行了迭代普鲁克分析旋转,以将参与者特定的梯度成分与年轻-老年群体平均模板对齐,并实现群体比较。使用基于表面的线性模型进行群体对比,如Surfstat(Worsley et al. 2009;)中实现的那样,控制性别、教育、地点和eWBV。
我们计算了流形离心率指标,以量化梯度空间中顶点的扩散性。遵循Bethlehem等人(2020)和Park等人(2021)的方法,我们对每个参与者在2D梯度空间中每个顶点与全脑中位数的平方欧几里得距离进行求和。顶点在梯度空间中的位置代表该顶点在每个梯度轴上的坐标。任何两个顶点的接近程度表明它们在每个梯度上的功能连接概况有多相似。网络内顶点越分散,功能连接概况就越可变和去分化。然后比较了年龄组之间的平均流形离心率。使用非参数自旋测试置换检验确定统计显著性,这克服了由于BOLD数据固有的空间自相关而导致的检验统计量偏差(Alexander-Bloch et al. 2018)。还进行了流形离心率的ANCOVA,控制性别、教育、地点和eWBV。
边水平连接组学
使用PLS(偏最小二乘法)检验了区域间功能连接的组间差异。PLS是一种多变量方法,通过识别两组变量中线性组合的最大协变来确定它们之间的关联(McIntosh and Lobaugh 2004; McIntosh and Mišić 2013)。关键是,PLS能够对未阈值化的连接矩阵进行全脑对比,从而更精确地映射边水平的年龄差异。在我们的分析中,一组变量是个体RSFC矩阵,而另一组代表组别分配或个体差异指标(如BOLD维度性;见补充材料)。
使用Schaefer图谱(Schaefer et al. 2018; Yeo et al. 2011; 400 × 400矩阵;200 × 200矩阵作为补充分析,补充图6)在全脑水平评估功能连接。受先前工作的启发(如Grady et al. 2016; Spreng et al. 2016; Sullivan et al. 2019),我们还检查了默认、背侧注意和额顶控制网络子网络之间的RSFC。对于子网络分析,我们首先根据Schaefer等人(2018)的映射,将400个分区重新分配到Yeo 17网络解决方案的相应网络。接下来,我们为8个子网络之间的成对连接创建了一个矩阵:背侧注意(DAN-A和DAN-B)、额顶控制(CONT-A、CONT-B和CONT-C)和默认(DN-A、DN-B和DN-C),得到一个192 × 192的分区矩阵。400 × 400分区结果的完整17网络特征,以及200 × 200矩阵的17网络和子网络特征,可在补充图7-9中找到。在每个水平,使用所有参与者的分区功能连接矩阵创建了一个数据矩阵X。X矩阵的组织方式是每行对应一个观察(每个参与者,嵌套在年龄组中),每列中的单元格对应于每个参与者连接矩阵中的唯一连接(矩阵的下三角)。计算了每组内的列均值,并对X中的数据进行了均值中心化。然后将均值中心化的数据提交给奇异值分解(SVD)以提供相互正交的潜变量。每个潜变量代表一个特定关系(如RSFC × 组),由三个元素组成:(i)左奇异向量,由最佳表达协方差的加权连接模式组成,(ii)右奇异向量,代表研究设计变量的权重,可以解释为组间数据驱动的对比权重,以及(iii)标量奇异值,代表每个潜变量所解释的设计变量(组)和RSFC之间的协方差强度。通过取左奇异向量和每个参与者RSFC矩阵的点积来计算大脑连接分数。因此,大脑连接分数代表参与者表达给定潜变量捕获的连接模式程度的单一度量。
所有PLS(偏最小二乘法)潜变量都使用置换测试进行了统计评估。X的行被随机重排并反复进行SVD(奇异值分解),如上所述。这个过程重复1000次,以创建在相应PLS分析的零假设下的奇异值分布:全脑(或子网络)RSFC没有组间差异。对每个潜变量计算P值,作为大于或等于原始奇异值的置换奇异值的比例。重要的是,置换测试涉及整个多变量模式,并在单个分析步骤中执行,因此不需要多重比较校正(McIntosh and Lobaugh 2004)。
使用自举重采样来估计每个RSFC边的权重可靠性。在保持组成员身份的同时,随机重采样参与者(X中的行)并进行替换。对矩阵进行SVD(奇异值分解),并重复该过程1000次,为奇异向量中的权重生成采样分布。为了识别对整体连接模式做出统计显著贡献的个别连接,我们计算了奇异向量中每个权重与其自举估计标准误之间的比率。如果自举分布近似单位正态,则自举比率等同于z分数(Efron and Tibshirani 1986)。自举比率在±1.96的值处阈值化,对应95%置信区间。
网络水平贡献
PLS(偏最小二乘)分析识别了组间不同和/或与个体差异指标协变的区域间连接模式。对于每个分析,还检查了网络水平效应。为了量化PLS导出的功能连接模式的网络水平贡献,从正负RSFC权重构建了两个单独的加权邻接矩阵。对于两个矩阵,节点代表由个体分区定义的分区,而边对应于每个成对连接的阈值化自举比率。通过根据Yeo等人(2011)报告的网络分配分配每个分区,并取给定网络中所有连接权重的平均值,从而量化网络水平功能连接贡献,生成7 × 7矩阵(17网络解决方案为17 × 17矩阵;检查默认、背侧注意和额顶控制子网络时为8 × 8矩阵)。通过置换测试计算网络内和网络间平均连接的显著性。在每次置换中,随机重排每个节点的网络标签,并重新计算网络内和网络间平均连接。重复此过程1000次,生成表明网络分配和连接模式之间没有关系的经验零采样分布(Shafiei et al. 2019)。通过估计采样分布值大于或等于原始值的比例来确定网络矩阵中成对连接的显著性。
弹簧嵌入图
使用Pajek软件(Mrvar and Batagelj 2016)从RSFC数据的组平均矩阵渲染弹簧嵌入图。包含前5%正连接的稀疏矩阵被输入Pajek。绘制正边权重,其阈值与之前健康老化研究(Chan et al. 2014; Geerligs et al. 2015)中应用的阈值相似,允许直接与先前结果进行比较。基于Yeo 7网络或17网络解决方案(Yeo et al. 2011)分配分区,以优化社区(即网络)结构以便可视化。
结果
为了探究老化大脑的内在功能架构,我们在年轻和老年健康成人中实施了多方面、多尺度的数据采集和分析方案(见图1和材料与方法)。为识别随年龄变化的网络去分化全局模式,我们首先评估了ME-fMRI BOLD信号维度性的年龄差异,该信号是ME-ICA的输出。接下来,我们检查了特定网络的去分化模式,对比年轻人和老年人之间的宏观尺度梯度和边水平网络连接组学。在每一步,我们都检查了网络组织与年轻人和老年人认知功能之间的关联。每项分析的脑-行为关联在补充材料中报告(补充表2-4;补充图10和12)。所有结果都在适当情况下报告了地点、性别、教育和eWBV的协变量。
BOLD维度性
收集了两次10分钟的静息态ME-fMRI扫描。BOLD维度性,即ME-fMRI信号中独立BOLD成分的数量,在不同扫描之间保持稳定(r(299) = 0.79,P < 0.001 [0.75, 0.83];图2a)。年轻成年人表现出比老年人更高的BOLD维度性(t(299) = 15.38,P < 0.001;Cohen's d = 1.81;图2b)。当包括地点、性别、教育和eWBV作为协变量时,这一结果依然成立(F(1,281) = 97.07,P < 0.001;ηp2 = 0.26)。在包括额外8-46岁样本(Kundu等人2018)的全生命周期发展背景下,幂函数为年龄和BOLD维度性之间提供了合适的拟合(R2 = 0.547;图2c)。BOLD维度性与认知的关联在补充表2-4和补充图10a中报告。
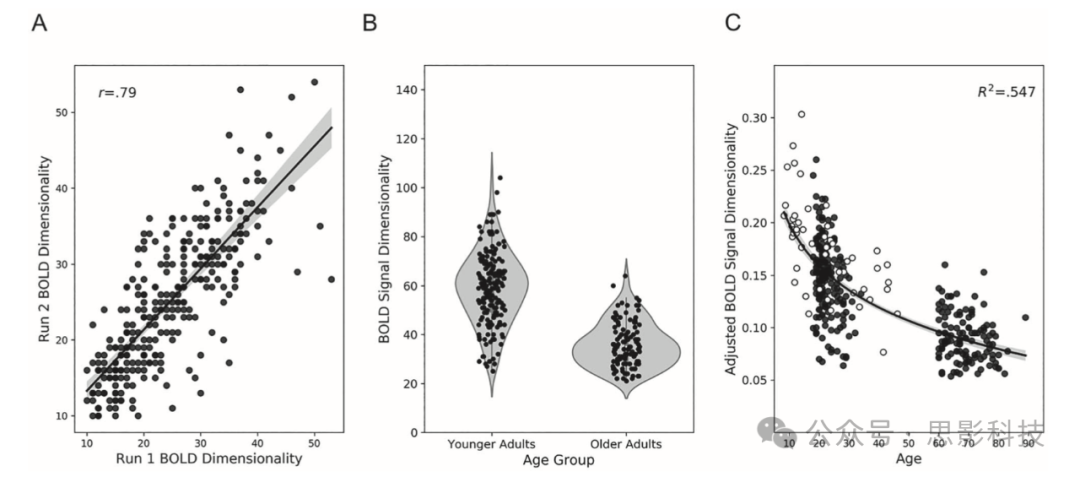
图2 BOLD信号维度性。
a) 两次ME-fMRI扫描之间的高测试-重测可靠性。
b) 小提琴图显示年轻人和老年人在不同扫描中总BOLD信号维度性的分布。
c) 散点图显示按年龄划分的BOLD信号维度性,叠加了幂分布和95%置信区间。白色点来自Kundu等人(2018)的贡献。调整后的BOLD信号维度性 = 接受的BOLD成分总数/获取的时间点数量。
梯度分析
接下来,我们描述了年轻人和老年人RSFC的宏观尺度梯度(如Margulies等人2016; Hong等人2019)。在两个组中,主梯度从感觉和运动区域延伸到诸如默认网络等跨模态系统(图3a),表明皮层的宏观功能组织随年龄总体保持不变。然而,这种地形组织中出现了区域性的年龄差异(FWE P < 0.05;簇定义阈值P < 0.01;图3a)。在主要梯度上的全皮层年龄组比较显示,老年人在右侧顶上小叶和体感皮层有更高的梯度值,但在枕叶和腹侧颞叶区域有较低的值。
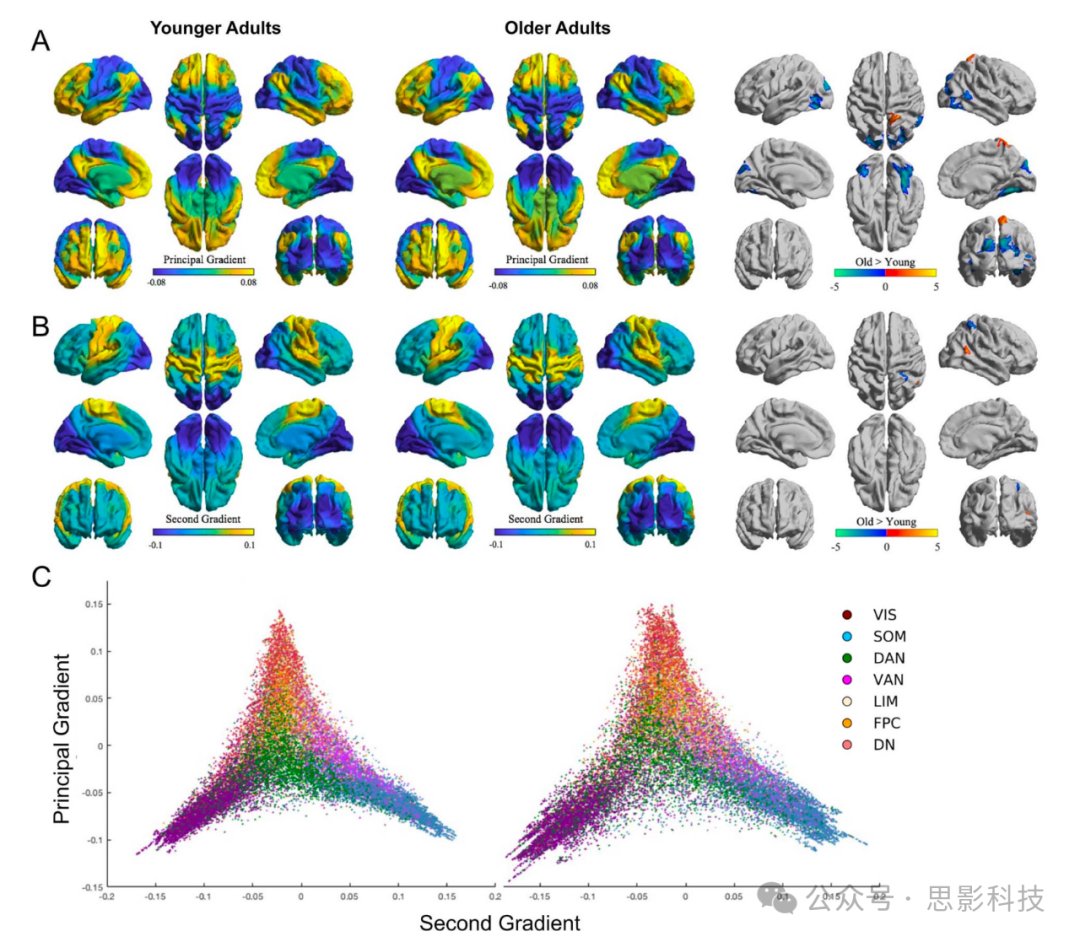
图3 年轻人和老年人皮层连接的梯度
a) 年轻人(左)和老年人(中)的平均主要梯度,代表从单模态到跨模态皮层的功能连接相似性方差轴,范围从最低到最高。
b) 年轻人(左)和老年人(中)的平均第二梯度,代表从视觉到体感运动皮层的功能连接相似性方差轴,范围从最低到最高。老年人 > 年轻人对比显示在FWE P < 0.05,簇定义阈值P < 0.01下的统计显著簇(a和b,右)。
c) 代表主要-第二梯度值的顶点散点图。这个梯度流形分别描绘了年轻人(左)和老年人(右)。散点图颜色表示根据Yeo等人(2011)的7网络解决方案的功能网络。
VIS = 视觉,SOM = 体感运动,DAN = 背侧注意,VAN = 腹侧注意,LIM = 边缘,FPC = 额顶控制,DN = 默认。
在检查第二梯度时,视觉和感觉/运动网络在皮层梯度组织方面的差异也很明显。如同先前的研究(如Margulies等人2016),第二梯度在两个组中都区分了视觉和体感运动皮层(图3b)。然而,组间全皮层比较揭示了这种地形学中的微妙差异。特别是,我们观察到老年人相比年轻人在颞顶交界处的梯度值增加,同时在顶上小叶/顶内沟的一个段落中值减少。这些结果表明,沿第二梯度轴的区域随年龄增长也会改变其连接概况,再次在感觉/视觉区域观察到变化。
最后,我们在2D梯度嵌入空间中为年轻人和老年人绘制了主要-第二梯度流形散点图(图3c)。老年人显示出更分散,因此去分化的顶点。我们通过计算流形离心率——每个参与者所有顶点与中位数的欧几里得距离之和——来量化这种分散性,并在组间进行比较。结果显示老年人的流形离心率显著更高(t(299) = −10.74,PSPIN < 0.01,Cohen's d = 1.26;F(1,281) = 47.18,P < 0.001,ηp2 = 0.14,包括地点、性别、教育和eWBV协变量)。有关与行为的关联,见补充表2-4和补充图10b。
由于BOLD维度性和流形离心率都表现出显著的年龄组差异,我们进行了事后积矩相关分析,以检验这些大脑组织的全局测量是否可靠地关联。在年轻人(r(179) = −0.575,P < 0.001,[−0.66,−0.47])和老年人(r(118) = −0.255,P < 0.005,[−0.42,−0.08])中都观察到负相关,即更高的BOLD维度性与流形中更不分散、更紧凑的顶点相关。在控制年龄的偏相关计算中,对全样本进行时关系仍然存在(Pr(298) = −0.391,P < 0.001,[−0.48,−0.29])。不重叠的95% CI表明年轻人的负相关显著更强。当重复使用协变量时,结果相似(年轻:Pr(161) = −0.45,P < 0.001,[−0.53,−0.36];老年:Pr(113) = −0.23,P < 0.05,[−0.35,−0.11];全样本:Pr(280) = −0.34,P < 0.001,[−0.44,−0.23]),尽管组间CI有重叠。补充图11说明了每个年龄组中的关系。
边水平连接组学
我们接下来检查了年轻人和老年人之间边水平的区域间功能连接差异。图4a和b显示了组平均连接矩阵。通过按网络成员排列的弹簧嵌入布局可以观察到组间前5%正连接的定性差异(图4c和d)。弹簧嵌入图表明老年人的背侧注意网络和额顶控制网络更加整合。
图4 年轻人和老年人的功能连接组学
a)年轻人和b)老年人400分区数据的平均RSFC。c)年轻人和d)老年人平均相关矩阵的7网络解决方案(5%边密度)的弹簧嵌入图。功能上高度相关的节点被分组在一起。e)使用多变量PLS分析来识别年轻人和老年人之间RSFC的年龄相关差异。红色表示年轻人的RSFC显著更高,蓝色表示老年人的RSFC显著更高。f)和g)网络贡献代表年轻f)和老年g)成人网络内和网络间显著正负边权重的总结。网络内和网络间的平均正负自举比表示为相对于置换零模型的每个z分数的P值。更高的z分数表示比零分布预测的更大连接。
VIS = 视觉,SOM = 体感运动,DAN = 背侧注意,VAN = 腹侧注意,LIM = 边缘,FPC = 额顶控制,DN = 默认。
PLS(全脑)
使用PLS(偏最小二乘)定量评估了79,800个区域间连接(即400 × 400功能连接矩阵的下三角)的年龄相关差异。一个显著的潜变量(置换P < 0.001)揭示了RSFC的年龄差异模式,在整个连接组中观察到增加和减少(图4e)。网络内和网络间边的网络贡献分析揭示了显著的年龄效应。老年人在所有7个网络中表现出较低的网络内连接,以及边缘、额顶控制和默认网络之间的较低连接(图4f)。老年人在视觉和体感运动网络的系统间表现出更大的网络间连接(图4g)。使用200分区方案检查时,年龄相关差异的总体模式相似(补充图6)。大脑连接分数与认知的关联在补充表2-4和补充图12中报告。
PLS(子网络)
在先验的、有针对性的子网络分析中,我们检查了默认、背侧注意和额顶控制网络子网络之间功能连接的年龄组差异。图5a和b显示了年龄组子网络的平均矩阵。每个组前5%正连接的弹簧嵌入表示(图5c和d)表明老年人显示出默认网络(DN-A)和额顶控制网络(CONT-C)的更多整合。

图4 遵循Yeo 17网络解决方案的默认(DN)、背侧注意(DAN)和额顶控制(CONT)子网络的功能连接。a)年轻人和b)老年人的平均组连接。c)年轻人和d)老年人平均相关矩阵的弹簧嵌入图(5%边密度)。功能上高度相关的节点被分组在一起。e)年轻人和老年人之间DAN、CONT和DN的RSFC差异。f)和g)网络贡献代表年轻f)和老年g)成人网络内和网络间正负边权重的总结。
PLS对区域间功能连接的定量比较揭示了一个不同的年龄差异模式(置换P < 0.001;图5e)。年轻成年人(图5f)显示出更多的网络内连接。在年轻人中也观察到CONT-A和CONT-B之间,以及DN-A到DN-B和DN-C之间的网络间连接。年轻人中CONT-B和DN-B之间也观察到网络间连接。老年人(图5g)显示出背侧注意网络与额顶控制和默认网络之间更大的网络间连接(DAN-A到CONT-B和CONT-C;DAN-B到CONT-B、CONT-C、DN-A和DN-B),以及额顶控制与默认网络之间更大的连接(CONT-A到DN-A;CONT-B到DN-C;CONT-C到DN-B)。老年人还显示出额顶控制子网络之间更大的连接(CONT-A到CONT-C;CONT-B到CONT-C)。使用200分区方案观察到类似的连接模式(补充图9)。子网络大脑连接分数与认知的关联在补充表2-4和补充图12中报告。
连接组学地点复制
为验证我们的边水平分析结果是稳健和可复制的,且不受PLS(偏最小二乘)模型潜在过拟合的影响,仅在Ithaca样本上进行了全部和子网络PLS分析。然后使用Ithaca样本得出的权重和Toronto样本个体RSFC矩阵计算大脑连接分数,并在组间进行比较。在保留的Toronto样本中复制了年龄组差异(t(61) = 6.42,P < 0.001,Cohen's d = 1.63;F(1,57) = 21.13,P < 0.001,ηp2 = 0.27,包括性别、教育和eWBV协变量)。在子网络分析中,年龄组差异也在保留的Toronto样本中得到复制(t(61) = 7.01,P < 0.001,Cohen's d = 1.79;F(1,58) = 24.16,P < 0.001,ηp2 = 0.29,包括性别、教育和eWBV协变量)。这些地点复制分析(补充图13)表明PLS结果对潜在的模型过拟合问题是稳健的,且在Ithaca样本中观察到的功能性大脑老化的边缘水平效应也在Toronto地点观察到。
认知
总体上,观察到了预期的年龄组认知差异。年轻成年人在情景记忆(t(281) = 17.51 P < 0.001;Cohen's d = 2.11)、执行功能(t(281) = 12.67,P < 0.001;Cohen's d = 1.52)和处理速度(t(281) = 15.03,P < 0.001;Cohen's d = 1.81)指数上表现更好。老年人有更高的语义记忆指数得分(t(281) = 9.18,P < 0.001;Cohen's d = 1.10;见表1)。在使用ANCOVA控制地点、性别、教育和eWBV测试年龄组差异时,效应仍然存在(情景:F(1,277) = 194.07,P < 0.001,ηp2 = 0.41;语义:F(1,277) = 37.55,P < 0.001,ηp2 = 0.12;执行功能:F(1,277) = 132.70,P < 0.001,ηp2 = 0.32;处理速度:F(1,277) = 97.21,P < 0.001,ηp2 = 0.26)。
检查了认知与BOLD信号维度性、流形离心率以及全脑和子网络分析的大脑连接分数之间的关联(见补充表2-4和补充图10、12和14)。虽然观察到几个显著的脑-行为关联,但在模型中加入地点作为协变量后,所有这些都低于统计显著性阈值。
讨论
大脑老化以大脑活动和功能连接模式的去分化为特征。在这里,我们采用了全面的多方法方法来检查多个空间尺度上内在网络去分化的模式。具体来说,我们应用新方法来识别年轻人和老年人之间RSFC的全局、宏观尺度梯度和边水平差异。BOLD维度性,即ME-fMRI信号中BOLD(即非噪声)成分的数量,在老年人中较低,表明老年时大脑网络向去分化的全局转变。相比之下,宏观连接梯度的组织在很大程度上随年龄保持不变。然而,连接梯度的区域和全局差异确实出现了。使用PLS进行的边水平多变量分析也揭示了老年人去分化的区域和网络特定模式。在整个皮层连接组中,与年轻成年人相比,老年人的视觉和体感运动区域与其他大尺度网络更加功能整合。在包括默认、背侧注意和额顶控制网络的有针对性的子网络分析中,老年人表现出更大的默认-执行耦合和默认与背侧注意网络之间减少的反相关。通过检查多个空间尺度上功能连接组的年龄差异,我们揭示了老化大脑的内在网络架构以全局以及地形离散的、网络特定的功能去分化模式为特征。这些发现为全局和网络特定的去分化模式提供了证据,为未来研究RSFC改变作为神经认知老化的潜在敏感和特定标记奠定了基础。
BOLD信号维度性和全局网络去分化
相比年轻成年人,老年人的BOLD信号维度性显著降低,反映了持续到晚年的网络去分化的普遍模式。这一发现建立在早期报告的从青春期到成年早期和中期维度性降低的横断面研究基础上(Kundu et al. 2018; 图2)。早期成人发展中维度性的降低主要归因于前额叶和其他跨模态皮层之间的功能整合,反映了从局部连接到远程连接的转变,以及空间分布但内在一致的大脑网络的形成(Kundu et al. 2018)。功能性大脑组织的这种转变与这一时期的认知发展相平行,后者以更具整合性和复杂性的认知功能的出现为特征(Zelazo and Carlson 2012),这在结构连接组中也是明显的(Park et al. 2021)。
BOLD信号维度性的下降始于青春期。在年轻成年人中,较低的维度性反映了更大的功能整合和大尺度大脑网络的出现(Kundu et al. 2018)。然而,我们观察到BOLD信号维度性持续降低到老年,这表明网络整合可能在中年达到拐点(Zonneveld et al. 2019)。在这一点之后,维度性的持续降低可能不再由网络整合驱动,而是由全局网络瓦解和BOLD信号中相干网络成分的相关丧失驱动。重要的是,我们使用这种新的BOLD信号维度性指标的发现与早期报告的年龄相关的网络模块性(Geerligs et al. 2015)和网络分离(Chan et al. 2014)降低一致。事实上,这些测量在我们的样本中与维度性可靠且正相关(见补充表1)。然而,与这两种网络组织的图论分析测量不同,BOLD信号维度性不依赖于皮层分区方案、网络定义或特定网络指标的选择。因此,我们认为维度性可能作为晚年功能性大脑健康的有用的、数据驱动的标记。在这方面,重要的下一步是改进我们对年龄相关维度性降低的机制理解。这种全局转变可能是由于随年龄增长而发生的系统性结构、神经生理、代谢或脑血管变化(如Tsvetanov et al. 2020; Kantarovich et al. 2022)。
最后,作为应用于健康老化样本的新指标,我们承认有重要的未来方向来更全面地质疑BOLD信号维度性作为功能性大脑老化信息标记的有效性和适用性。需要额外的工作来对这一指标进行验证,遵循原始验证研究在年轻和中年成年人中概述的路线图(Kundu et al. 2013, 2017, 2018)。作为T2*信号衰减的物理特性,BOLD信号的TE依赖性(驱动ME-ICA BOLD信号去噪)应该在很大程度上对年龄差异具有鲁棒性。直接测试这一假设将是未来研究的重要方向。总的来说,BOLD信号的TE依赖性以及在健康年轻样本中进行的验证研究使我们对BOLD信号维度性作为大脑老化的可靠、信息丰富的标记充满信心。
梯度和宏观尺度连接组学
老年BOLD信号维度性的降低表明向去分化网络架构的全局转变。我们调查了这种全局转变是否可能包括更精确的地形模式,反映为大脑区域之间连接概况的更大相似性。我们通过检查年轻人和老年人的宏观连接梯度来测试这一假设。虽然这是首次报告使用ME-fMRI和个体化分区方法的梯度分析,但我们的发现在很大程度上重现了在年轻成年人中观察到的连接梯度(Margulies et al. 2016)。观察到从感觉/运动到跨模态联合皮层(主要梯度)和从视觉到体感运动皮层(第二梯度)的功能连接模式转变。这种梯度架构在年轻人和老年人中相似,表明梯度的宏观组织通常随年龄保持不变,这与之前的观察一致(Bethlehem et al. 2020)。然而,在两个梯度图中都出现了特定的年龄相关区域差异。
两个梯度中的年龄相关差异包括视觉、体感运动和注意网络内的区域。这些簇内的差异表明,相对于其对应的梯度锚(主要梯度中的单模态或跨模态,第二梯度中的体感运动或视觉),分化程度降低。具体来说,背侧注意网络的节点——被认为参与外部导向注意和视觉运动控制过程的顶上小叶,以及体感运动区域显示出与跨模态区域更相似的连接概况。这与早期报告一致,也与当前边水平分析中观察到的模式一致,即晚年背侧注意网络和默认网络之间反相关减少(Spreng et al. 2016)。
沿主要梯度,视觉区域在老年人中更突出地锚定在单模态轴上。这一发现与RSFC的弹簧嵌入图(图4c和d)一致,前5%中视觉区域更加孤立。这表明老年人的主要梯度可能更多地由异质模态系统与视觉系统的分化驱动,而在年轻成年人中,沿这个轴线异质模态系统与体感/运动系统的分化更为显著。这一发现与边水平结果(下面讨论)形成对比,后者显示视觉和体感运动皮层与异质模态联合区域的年龄相关整合增加。重要的是,梯度并不直接指示功能连接强度,而是特定区域与皮层其余部分之间RSFC的低维模式。因此,连接概况向视觉区域的转移并不涉及这些区域的功能整合,而是它们在梯度轴上的权重。在梯度映射之前对连接矩阵进行阈值处理可能导致了第一梯度单模态锚向视觉皮层的年龄相关转移。虽然是推测性的,我们认为这种阈值处理可能将超阈值连接驱动向更强的局部连接而不是更弱的远程功能连接的过度表示。这与初级感觉区域中长程联合纤维通路相对于局部连接的体积和完整性的年龄相关减少一致(Raz et al. 2005; Kochunov et al. 2012)。这些年龄相关的结构差异反过来会使功能连接概况偏向更局部化的模式,导致梯度图向单模态锚的年龄相关转移。尽管如此,这些发现强调了阈值决策的重要性和潜在影响,我们在下面讨论边水平连接时会回到这一点。
梯度流形,即前两个连接嵌入梯度的散点图,代表了功能层次结构的组织(图3c)。流形离心率描绘了区域与流形中心的欧几里德距离,显示了第一梯度对第二梯度的相互关系。在老年人中,我们观察到流形中顶点的更大扩散,离心率更高。这不限于视觉或体感区域,整个流形组织更加分散,表明全局去分化。这一观察与之前观察到的整个生命周期中流形分散增加一致(Bethlehem et al. 2020)。这种流形离心率指标在组内和组间与BOLD维度性的全局测量呈负相关。在老年人中观察到的较低BOLD维度性与较高的流形离心率相关,为全局去分化提供了跨方法的收敛证据。
需要注意的是,我们应用了扩散图嵌入,这是一种来自图拉普拉斯家族的非线性维度流形学习技术(Coifman et al. 2005)。这种方法是文献中最广泛实施的方法之一(如Margulies et al. 2016; Hong et al. 2019; Murphy et al. 2019; Bethlehem et al. 2020; Vos de Wael et al. 2020)。然而,鉴于梯度映射在老年人群中的新颖性,未来研究的一个方向将是批判性地评估全范围的方法以及算法参数(Hong et al. 2020),包括在梯度分析中纳入排斥属性(Böhm et al. 2020),以更清晰地了解离散网络的分离和年龄差异。
边水平连接组学
为了更精确地研究边水平连接模式,我们采用了多变量分析方法。由于PLS(偏最小二乘)使用SVD(奇异值分解)在单个分析步骤中测试所有边的年龄差异,我们报告了整个功能连接组的RSFC差异,消除了应用功能连接强度或密度阈值的需要。对年轻人和老年人完整连接组的视觉检查(图4a-d)揭示了老年人网络去分化的全局模式,与我们的维度性发现和先前报告一致(Betzel et al. 2014; Chan et al. 2014; Geerligs et al. 2015; Malagurski et al. 2020; Stumme et al. 2020)。这些定性差异在组分析(图4e)和聚合网络矩阵(图4f和g)中得到了统计验证。如预测的那样,年轻成年人显示了强健的网络内连接模式,以及跨模态网络之间的连接(Bullmore and Sporns 2009; Gratton et al. 2012)。
尽管宏观尺度梯度得以保存,边水平分析揭示了网络特定连接模式的显著年龄差异。首先,老年人在这里研究的7个经典网络中的网络内连接较低。减少的网络内功能连接是正常老化的标志(Damoiseaux 2017综述)。我们推测,网络内一致性的降低可能是老年BOLD信号维度性降低和全局网络去分化的关键决定因素。除了较低的网络内一致性,边水平分析还揭示了3种不同的、网络特定的去分化模式。最显著的是老年人视觉和体感运动区域与所有其他网络的更大整合(图4g)。之前已经观察到视觉和体感区域的功能整合。Chan等人(2014)报告了视觉皮层与其他大脑网络的分离减少,尽管他们的分析中没有明确量化这一点。同样,在一项关于RSFC年龄差异的大规模研究中,节点参与度(功能整合的图论标记)的年龄相关增加仅限于视觉和体感网络(Geerligs et al. 2015)。此外,Stumme等人(2020)报告RSFC的年龄差异在视觉和体感皮层最为显著。虽然先前的研究报告了随年龄增长的感觉运动整合模式,但这些并未成为功能性大脑老化的核心特征。如上文关于梯度分析结果的讨论所述,梯度矩阵的统计阈值处理可能显著影响这些发现。基于阈值的方法强调了最强连接中的年龄相关差异,这些连接通常与异质模态皮层相关,可能掩盖了其他网络中不太显著的年龄差异。这在我们的发现中特别明显,其中体感运动和视觉网络在阈值化的弹簧嵌入图(图4c和d)中相对于联合网络观察到的差异显示出小的年龄差异。相比之下,对未阈值化矩阵的分析揭示感觉运动网络的整合是老化连接组最显著特征之一(图4e-g)。
我们关于视觉网络更大整合的发现与基于任务的研究相平行,这些研究将跨模态区域对感觉联合皮层的更大自上而下调节识别为功能性大脑老化的中心特征(Alain et al. 2022)。在感觉信号保真度年龄相关下降的背景下,跨模态皮层的更大激活被解释为早期感觉处理的自上而下调节需求增加(Payer et al. 2006; Clapp et al. 2011; Li and Rieckmann 2014; Spreng and Turner 2019b)。事实上,感觉下降和运动减缓解释了老年人认知功能个体变异的大部分(Salthouse 1996; Baltes and Lindenberger 1997)。这表明,对这些初级感觉运动区域(以及顶上小叶的视觉注意和视觉运动控制区域,见上文"梯度分析")的更大调节可能是维持晚年复杂思维和行动所必需的(Alain et al. 2022)。虽然超出了当前研究的范围,我们推测这种跨模态和感觉运动皮层之间更大交互的任务驱动需求可能反过来塑造这些网络在老年的内在功能架构(Stevens and Spreng 2014)。
我们还对默认、背侧注意和额顶控制网络进行了针对性的边水平年龄差异分析。先前的工作表明,这些网络在目标导向的认知任务中相互作用(Spreng et al. 2010; Dixon et al. 2018; Murphy et al. 2020),在任务和休息期间显示相似的连接概况(Spreng et al. 2013),并在老年期经历显著变化(Grady et al. 2016; Ng et al. 2016; Sullivan et al. 2019)。对于这个先验分析,我们采用了从17网络解决方案(Yeo et al. 2011)衍生的3个网络的子网络地形。这使我们能够更精确地研究年龄相关变化。重要的是,正如我们在全连接组分析中观察到的那样,阈值化的弹簧嵌入图(图5c和d)未能揭示边水平分析中出现的默认、背侧注意和额顶控制网络区域之间连接的显著年龄差异(图5e-g)。虽然预测的网络内连接减少模式在子网络中得到重现,但我们观察到两种额外的网络特定去分化模式。如预测的那样,默认和额叶脑区的年龄相关耦合增加,这是我们描述为老化的默认到执行耦合假说(DECHA; Turner and Spreng 2015; Spreng and Turner 2019a)的模式。这种模式在7网络分析中没有出现(图4)。然而,当应用于边水平子网络矩阵(图5e-g)时,CONT-A到DN-A、CONT-B到DN-C和CONT-C到DN-B子网络出现了明显的DECHA模式(图5g)。虽然我们在这里没有发现与认知的可靠关联,但我们认为这种去分化模式可能反映了晚年认知架构的转变(Turner and Spreng 2015; Spreng et al. 2018),对认知老化有积极和消极的后果(Spreng and Turner 2019a)。
在这个子网络分析中出现了第二种去分化模式。老年人显示背侧注意网络与其他两个联合网络之间的连接增加。这种模式在DAN-B子网络中特别明显,该子网络包括顶上小叶。先前的报告显示老年人背侧注意和默认网络之间的反相关减少(Keller et al. 2015; Spreng et al. 2016)。这些边水平发现也与我们的梯度分析收敛,其中DAN-B的节点顶上小叶在连接梯度上显示年龄差异,其功能连接概况与其他跨模态区域更相似。DAN-B子网络包括涉及自上而下或目标导向注意控制的额叶眼场和中心前回区域。这再次与神经认知老化的神经调节解释一致,即更多的注意资源分配可能被用于提高晚年的感知表征(Li et al. 2006; Li and Rieckmann 2014; Alain et al. 2022)。未来的研究将需要直接测试这些假设,将网络特定的去分化模式与领域特定的认知变化联系起来。
认知功能
我们的发现表明,全局和网络特定的去分化是功能老化连接组的核心特征。在最后一系列分析中,我们调查了这些网络变化是否与认知功能相关。我们观察到BOLD信号维度性和边水平连接与行为的显著相关。然而有趣的是,当在统计模型中包括地点作为协变量时,所有观察到的关联都低于统计显著性阈值。即使大脑和行为年龄效应在两个地点都得到复制,也是如此(见补充图13和补充表5)。因此,我们在这里不进一步解释大脑和行为关联,并在补充表2-4中报告所有未校正和偏相关(见地点特定散点图补充图14)。虽然我们非常小心地匹配数据收集协议和核心人口统计,但研究地点包括许多可能影响两个地点大脑和行为关联的额外调节因素(如社会经济地位,见Chan等人2018)。虽然多地点研究增加的统计能力允许更高的敏感性来检测大脑-行为关联,但它也可能带来与人口差异相关的结构化噪声的潜在成本。理解这些差异也将是未来研究的重要方向。
结论
我们采用了多方法数据采集和分析协议来研究多个空间尺度上的功能性大脑老化,特别强调了内在网络去分化的年龄相关模式。BOLD信号维度性的降低表明,与年轻成年人相比,老年人的网络组织向去分化方向全局性地转变。横断面研究设计的局限性限制了对生命周期大脑功能转变的解释。然而,我们推测整个成年生命周期的网络整合可能包括中年期的一个拐点,在此之后,早期成年的网络整合转变为网络去分化的模式,以及分段和模块化网络架构的解体。
这里采用的方法和分析方法被选择用来至少部分克服生命周期网络神经科学中最持久和普遍的几个挑战。这些包括BOLD信号中噪音概况的年龄相关变异性,以及通过组间空间对齐到标准化模板引入的扭曲。当然,这些方法无法解决所有复杂化RSFC分析的混淆因素。其中最关键的是解决,或至少准确建模年龄差异在神经血管耦合方面的影响。随年龄变化的神经血管耦合可能引入难以用标准成像协议检测到的虚假RSFC差异(Tsvetanov et al. 2020)。虽然将神经和非神经来源在BOLD信号中分离的ME-ICA方法是一个重大进步,但实施多模态方法,如同时动脉自旋标记和回波平面成像可能是解决这个问题所必需的(Tsvetanov et al. 2020)。此外,在BOLD信号中观察到残余的与运动相关的噪音,这可能归因于呼吸(例如Power et al. 2018; Lynch et al. 2020)。虽然这种噪音没有混淆我们的年龄效应,但其持续存在需要额外考虑,并指出了需要进一步改进以提高ME-fMRI数据信噪比的必要性。尽管存在这些局限性,我们认为这里采用的多方面方法为大脑功能网络架构的年龄差异提供了全面的解释,包括新颖的和先前观察到的网络去分化和整合模式。总的来说,这些发现进一步明确和精确了当前对功能网络如何形成、塑造和转变到老年期的理解。

















 2565
2565

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









