







Notch 信号通路在各种癌症类型中既是致癌因子又是肿瘤抑制因子。该通路的失调促进恶性肿瘤的上皮-间质转化和血管生成,与癌症增殖、侵袭和转移密切相关。此外,Notch 信号通路有助于维持癌细胞中的干细胞样特性,从而增强癌症侵袭性。Notch 信号通路在癌症代谢重编程和肿瘤微环境中的调节作用表明其在平衡致癌和肿瘤抑制作用中起着关键作用。
一、NOTCH信号通路组成部分
Notch 通路由四部分组成:Notch 受体 (Notch1/2/3/4),Notch 配体 (Jagged ligands, JAG1/2;Delta-like ligands, DLL1/3/4),CSL-DNA 结合蛋白,下游靶基因 (Hes 家族,MYC 等)。配体激活的 Notch 受体通过与 DNA 结合的 CSL 辅阻遏蛋白复合体相互作用来启动下游靶基因的转录,形成经典的 Notch 信号通路。
Notch 受体是一种跨膜蛋白,具有三个主要部分:Notch 胞外结构域 (NECD)、跨膜结构域 (TMD) 和 Notch 胞内结构域 (NICD)。NECD 包含多个 EGF(表皮生长因子)样重复序列和一个负调节区 (NRR),由 O-糖修饰以调节 Notch 受体对不同配体的亲和力。NRR 由三个富含半胱氨酸的 Lin12-Notch 重复序列组成,稳定 NECD 和膜结合的 NICD 相互作用,这对受体切割至关重要。TMD 包括一个细胞外短区域和形成异二聚体的保守半胱氨酸残基。NICD 由一个 RBPJ [重组信号结合蛋白-J] 关联模块 (RAM) 结构域、七个锚蛋白重复序列 (ANK) 结构域和 ANK 结构域两侧的两个核定位序列 (NLS) 组成。Notch1 和 Notch2 在 ANK 序列后具有转录激活结构域 (TAD),而 Notch3 和 Notch4 缺乏 TAD。NICD 的 C 端有一个“PEST”序列,富含脯氨酸、谷氨酸、丝氨酸和苏氨酸,这对 NICD 的稳定性至关重要。
人类和小鼠具有 5 个与细胞外 Notch 受体片段结合的配体。根据是否存在富含半胱氨酸的区域,Notch 配体分为锯齿样配体 Jagged1 (JAG1) 和 JAG2,以及 δ 样配体 DLL1、DLL3 和 DLL4。JAG1/2 的胞外结构域由 DSL 结构域、EGF 样重复序列和富含半胱氨酸的区域组成。DLL1/3/4 的胞外结构域类似于 JAG1/2,但缺乏富含半胱氨酸的区域。
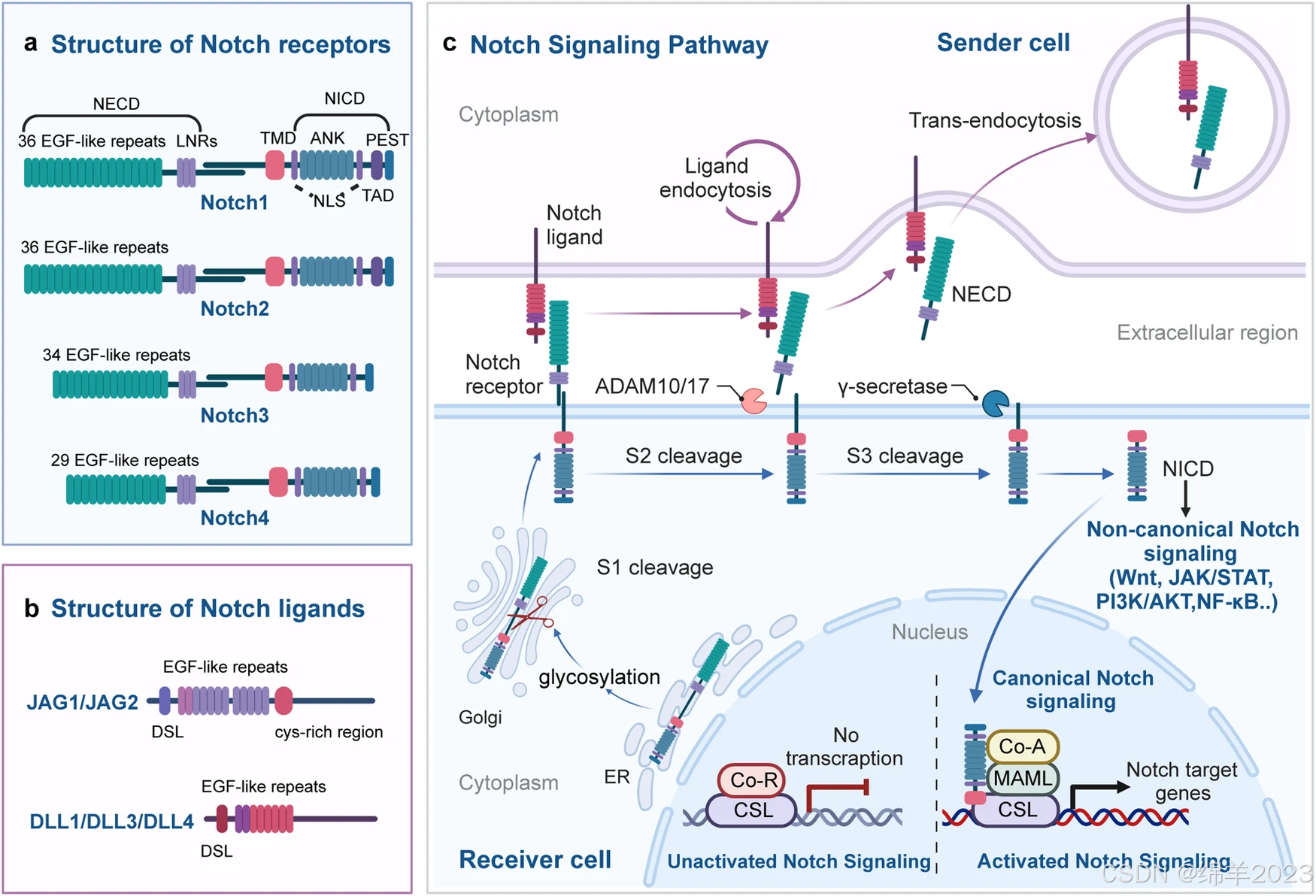
Notch 信号概述图
二、经典 Notch 信号通路
最初,Notch 蛋白作为无活性的单链前体转运到内质网。在内质网内,Notch 受体的 EGF 样结构域发生糖基化。然后将糖基化的 Notch 单链转运到高尔基体。在高尔基体中,转化酶裂解 Notch 跨膜区细胞外片段中的 S1 位点,导致形成两个不同的片段:NECD 和 TMD。这些片段随后形成异二聚体形状的成熟 Notch 受体。成熟的 Notch 受体随后被转运到细胞表面。到达细胞表面后,Notch 异二聚体跨膜受体与相邻细胞上的 Notch 跨膜配体结合。然后,Notch 受体的 S2 切割位点被 ADAM(一种解整合素和金属蛋白酶)金属蛋白酶家族的成员,特别是 ADAM10 或 ADAM17 切割。这种切割释放出部分细胞外片段,产生一种称为"NeXT"[Notch 细胞外截断]的瞬时中间肽,它由 TMD 和 NICD 组成。下一步 γ-分泌酶在 S3 切割位点裂解 NeXT。这个过程导致 Notch 的可溶性 NICD 的释放。随后,NICD 易位到细胞核,在那里其 RAM 结构域与转录因子 CSL(也称为 RBPJ) 相互作用。这种相互作用促进了共激活因子复合物向 CSL 的募集,包括 MAML1-3 蛋白。这些复合物的组装将原始的“共阻遏复合物”转化为“共激活因子复合物”,从而形成多蛋白质-DNA 复合物,促进 Notch 靶基因的转录。在没有 NICD 结合的情况下,CSL 通过募集各种共阻遏蛋白来下调靶基因的表达。
非经典 Notch 信号通路
Notch 信号通路除了与 CSL 相互作用外,还可以通过非 CSL 依赖性调控通路影响相关基因的表达,构成非经典 Notch 信号通路。该通路可能由不依赖配体的机制启动,并且可能不需要 Notch 受体切割。研究表明 Notch 可以调节 Wnt/β-catenin 信号通路,JAK/STAT 通路,PI3K/AKT 通路,和 NF-κB 通路在翻译后水平,从而发挥其非经典生物学功能。
三、NOTCH信号通路与癌症
Notch 信号通路在生理条件下调节细胞命运决策中起着至关重要的作用,影响细胞增殖、分化、发育和体内平衡。然而,在各种人类恶性肿瘤中越来越多地观察到 Notch 信号通路的失调。
在 HCC 中已观察到 Notch 受体及其配体表达失调。Ahn 及其同事在 288 例 HCC 病例中观察到 Notch1、Notch3 和 Notch4 的细胞质表达分别为 50.3%、20.8% 和 59.7%。Notch1 表达和 Notch4 过表达可能独立预测 HCC 患者的不良生存率。另一项研究表明,在 HBV 相关的 HCC 组织中,Notch1 或 Notch4 的表达与 HBx 相关,表明 HBx 可能通过调节 Notch 通路在致癌作用中发挥作用。Notch2 与肝癌的发生密切相关。Hayashi 等人通过免疫组化发现 19% 的人原发性 HCC 的 Notch2 核染色呈阳性。与此一致,在人 HCC 细胞系中观察到 Notch2 的上调。在功能上,Michael 等人发现肝脏中的组成型 Notch2 信号通过促进增殖和分化程度较低的 HCC 加速了二乙基亚硝胺诱导的肿瘤发生。与正常肝组织相比,Notch3 在 HCC 中过表达,并且与侵袭性增加和生存期缩短呈正相关。另一项研究报告了 Notch3 和 Notch4 在 78% 和 68% 的 HCC 组织中的异常积累。JAG1 在 HCC 中高表达,JAG1 和 DLL4 在 HCC 细胞中的表达分别为 57.2% 和 88.9%。然而,尚未观察到 DLL4 表达与临床特征之间的相关性。在 HCC 组织中也观察到 JAG2 表达上调,并且与不良的临床病理特征相关。靶向抑制 JAG1 和 JAG2 有望在 HCC 中起肿瘤抑制作用。总之,这些研究表明 Notch 家族成员可作为 HCC 患者的潜在预后指标。
参考文献:Notch signaling pathway in cancer: from mechanistic insights to targeted therapies

















 196
196

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









