







Wnt 基因最初来源于小鼠乳腺癌中的整合酶-1 和果蝇的无翼基因。由于这两个基因和功能蛋白相似,研究人员将这些术语组合为 Wnt 基因。Wnt 信号通路包括非经典和经典通路。非经典 Wnt 通路独立于 β-catenin-T 细胞因子/淋巴增强子结合因子 (TCF/LEF),例如 Wnt/Ca2+通路和非经典 Wnt 平面细胞极性。经典 Wnt 通路,也称为 Wnt/β-catenin 通路,涉及 β-catenin 的核转位和通过 TCF/LEF 转录因子激活靶基因。经典 Wnt 通路主要控制细胞增殖,而非经典 Wnt 通路调节细胞极性和迁移,两种主要通路形成相互调控网络。Wnt 信号通路与肝脏代谢和再生、肺组织修复和代谢、毛囊更新、造血系统发育以及成骨细胞成熟和活性密切相关。
Wnt/β-catenin 通路包括四个片段:细胞外信号、膜片段、细胞质片段和核片段。细胞外信号主要由 Wnt 蛋白介导,包括 Wnt3a、Wnt1 和 Wnt5a。细胞膜段主要包含 Wnt 受体 Frizzled(特异性七重跨膜受体 Frizzled 蛋白)和 LRP5/6。细胞质段主要包括 β-catenin、DVL、糖原合成酶激酶-3β (GSK-3β)、AXIN、APC 和酪蛋白激酶 I (CK1)。核段主要包括 β-catenin,它易位到细胞核、TCF/LEF 家族成员和 β-catenin 下游靶基因,如 MMPs 和 c-Myc。
经典 Wnt 通路通常高度保守,并通过自分泌/旁分泌方法将细胞外 Wnt 配体与膜受体结合而被激活。一旦被激活,典型的 Wnt 通路会诱导 β-catenin 的稳定性并将其转移到细胞核,最终促进参与细胞增殖、存活、分化和迁移的基因的表达。
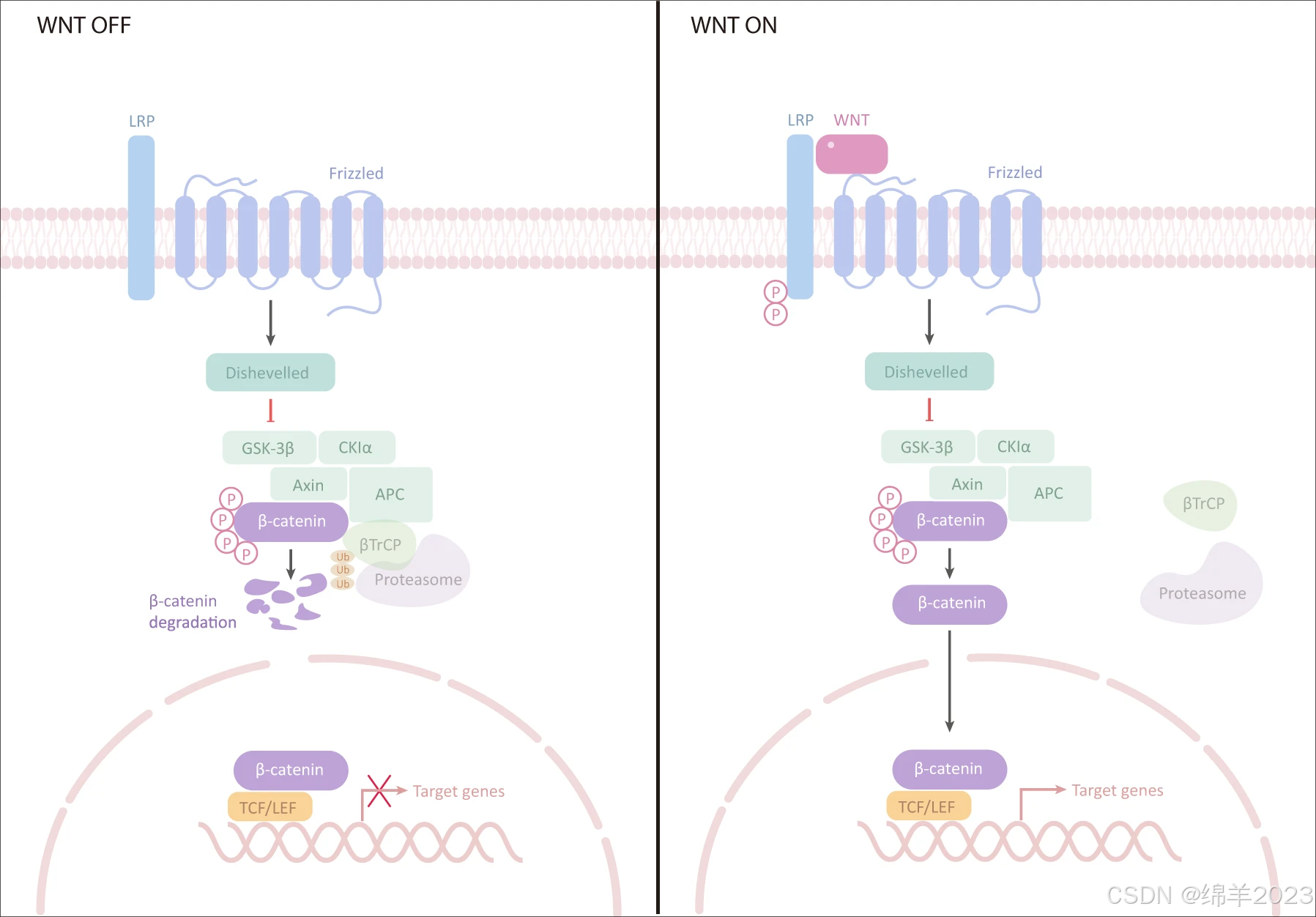
在没有 Wnt 的情况下,跨膜受体 FZD 和 LRP5/6 分别位于质膜上。在细胞质中,由 APC、AXIN、CK1α 和 GSK-3β 等蛋白组成的“破坏复合物”通过捕获 β-catenin 并降解。
当 Wnts 被 FZD 和 LRP5/6 识别时,“破坏复合物”无法形成,从而失去降解 β-catenin 的能力。β-catenin 易位到细胞核,并通过与 TCF/LEF 相互作用激活靶基因的转录。
参考文献:
Wnt/β-catenin signalling: function, biological mechanisms, and therapeutic opportunities

















 327
327

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









