






来自:微生物代谢工程与合成生物学Lab

商业化生物催化剂需要能够承受高温和长时间孵育的鲁棒性酶。祖先酶的重建表明,寒武纪前的酶通常比现存形式的热稳定性更好。来自昆士兰大学的Yosephin Gumulya等人在《Nature catalysis》上发表了题为“Engineering highly functional thermostable proteins using ancestral sequence reconstruction”的研究论文,作者通过祖先酶重构,仅使用近期祖先的序列数据,设计出了获得了可以承受30°C以上高温,并且孵育时间比现存形式长100倍以上的动物细胞色素P450和细菌酮醇酸还原异构酶热稳定蛋白。与人类药物代谢的CYP3A4酶相比,脊椎动物CYP3 P450酶的祖先酶显示出60°C的60T50,具有更高的有机溶剂耐受性,并且对相似范围广泛的底物具有可比拟的活性。酮醇-酸还原异构酶的祖先酶在25°C时的比活度比同源大肠杆菌高8倍,在50°C时比活性高3.5倍。
主要内容
在二十一世纪新兴的生物经济中,酶作为生物催化剂,将在日益广泛的应用领域中发挥关键作用,包括可持续的绿色精细化学品和生物燃料的生产。然而,目前酶的应用受到其稳定性差和生产成本高的限制,这些方面与传统化学催化剂相比处于劣势。热稳定蛋白在合成生物学中也备受青睐,因为它们更易于结晶,也更适用于生物物理特性表征所需的其他程序。
人们已经尝试通过工程手段提高酶的热稳定性和溶剂耐受性,但尚未建立一种能够先验设计出稳定性大幅提升的蛋白质的通用策略。对于大多数蛋白质而言,关于序列-结构-功能关系的信息不足,无法进行有效的理性设计。特别是,当将单个稳定突变组合在一起时,很难预测会出现的上位效应。虽然已经应用了计算方法,但这些方法依赖于蛋白质结构,并且通常需要耗时和资源密集的研究来提供训练数据。祖先酶重建表明,与前寒武纪时代的酶相比,现存形式的酶可能热稳定性较低。更常见的是,定向进化,有时结合有针对性的理性设计、共识突变或祖先突变,已被用于筛选在热稳定性、溶剂耐受性和/或其他理想特性(如底物特异性和催化效率)方面有所改进的酶。然而,这通常需要多次诱变和筛选大量突变体才能实现逐步改进。
一个典型的例子是CYP102A1,它是细胞色素P450(P450)酶家族的代表,这是一种无处不在的含血红素的单加氧酶,可催化未活化C-H键的区域和立体选择性功能化。这仍然是生产高价值化学品中最具挑战性和因此也最令人垂涎的反应之一,而P450(尤其是CYP102A1)在生物催化方面引起了人们的极大兴趣。CYP102A1已成为开发多种定向进化方法的模型系统。工程化热稳定形式的CYP102A1需要多轮点突变,才能使变性温度比亲本形式提高约6°C。重组定向进化使变性温度进一步提高了3.4°C。最后,基于重组CYP102库筛选获得的热稳定性数据,通过高斯过程景观建模使变性温度又提高了约5.3°C。
以CYP102A1为例,酶的重组进化已证明能够产生具有多样性和新颖特性的突变体库,其探索的可行序列空间范围比随机或位点饱和诱变等方法更广。然而,当在定向进化实验中体外重组在自然进化过程中不同进化分支上产生的结构上不相容的残基时,所得突变体的适应性(即稳定性和活性)可能会受到损害。研究者假设,即使是一个相对近代的、来自酶家族的中温祖先,也应该与在不同后代进化分支中出现的突变在本质上更加兼容,因此,与任何现存形式相比,它都将是重组定向进化以开发更稳定酶的更有用支架。
作为模型系统,研究者选择了与生物催化相关的两个不相关的酶组:P450和酮醇酸还原异构酶(KARI)。虽然细菌CYP102A1对其天然底物的稳健表达和高周转率使其成为蛋白质工程的主要关注点——特别是为了改变其底物特异性,从脂肪酸转变为具有商业价值的底物——但已经代谢这些感兴趣底物的其他P450提供了定向进化的另一个起点。特别是,异源生物代谢P450,如CYP3A4和CYP2D6,对许多工业上有用的化学骨架表现出广泛的底物混杂性和催化多功能性,负责人体中至少40%的药物代谢清除。然而,热稳定性差、对溶剂敏感以及周转率低等特性阻碍了这些酶在工业上的应用,促使人们努力工程化具有更理想特性的形式。
KARI是支链氨基酸生物合成途径中的第二种酶,也是生产生物燃料的酶级联反应的关键组成部分。该酶有两种结构类:I类长约390个氨基酸,II类长约550个氨基酸。两类KARI都采用保守的催化机制,但较大的II类KARI的活性位点完全位于一个多肽链内,这有利于在无细胞环境中生产生物燃料。因此,II类KARI一直是提高热稳定性、异丁醇耐受性、催化速率和其他特性的工程化主要目标。
在这里,研究者证明,通过重建即使是近期的祖先,如细胞色素P450(P450)CYP3家族的脊椎动物祖先,也可以实现通常与前寒武纪重建相关的热稳定性和持久性的大幅增加。这种稳健的CYP3祖先比现存的人体药物代谢形式CYP3A4具有更高的溶剂耐受性,但以相似的效率代谢了相似范围的底物。此外,与现存的大肠杆菌形式相比,祖先细菌KARI酶表现出八倍增强的比活性以及更好的热稳定性。因此,祖先酶可能为生物催化中的酶进化提供了显著改善的起点。
结果
研究者聚焦于现存P450家族中最具混杂性的一个家族,推断并复活了CYP3家族的祖先(图1a),命名为CYP3_N1,据推测,它存在于约4.5亿年前的第一批脊椎动物中。这种祖先蛋白可以在大肠杆菌中表达为血红素蛋白,并显示出Fe(ii)·CO与Fe(ii)之间的特征差异光谱,这表明它是一种正确折叠的P450酶。该酶的表达水平超过500 nmol l⁻¹(约25 mg l⁻¹)23。令人惊讶的是,即使在没有进行任何定向进化之前,与几种现存的CYP3A酶(包括人类CYP3A4和CYP3A5,以及鱼类形式的CYP3A27和鸟类形式的CYP3A37)相比,CYP3_N1已经表现出高度的内在热稳定性。其60T50(即在60分钟后仍有50%蛋白质保持折叠状态的温度)为66 ± 2 °C,比现存CYP3酶高约30 °C(图1b)。与CYP3A4相比,CYP3_N1的最适温度也提高了约20 °C(图1c)。
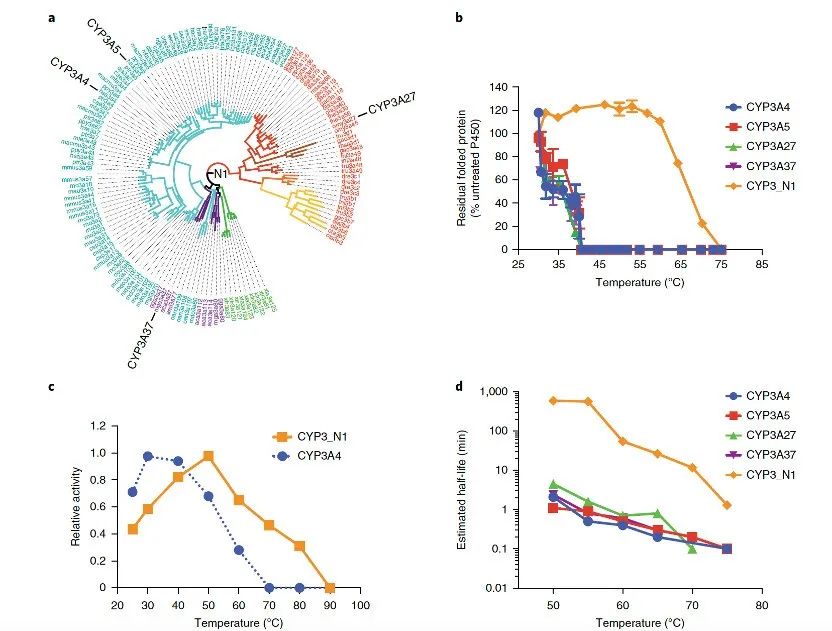

生物催化过程的经济可行性取决于能否最大化酶反应的体积-时间产率;较高的温度稳定性意味着在更温和的工艺条件下具有更长的半衰期。在光谱测定的灵敏度范围内,无法在37 °C下连续数天内检测到CYP3_N1的任何损失;然而,如特征光谱保留所示,其在50 °C和60 °C下的半衰期相较于CYP3A4和其他现存的CYP3形式,从1-5分钟延长至约10小时(图1d)。在45 °C至80 °C之间,祖先酶和现存CYP3酶均表现出一级失活动力学,根据阿伦尼乌斯图,CYP3_N1的失活能为251 kJ mol⁻¹,是现存P450酶(91–151 kJ mol⁻¹)的1.7至3倍。
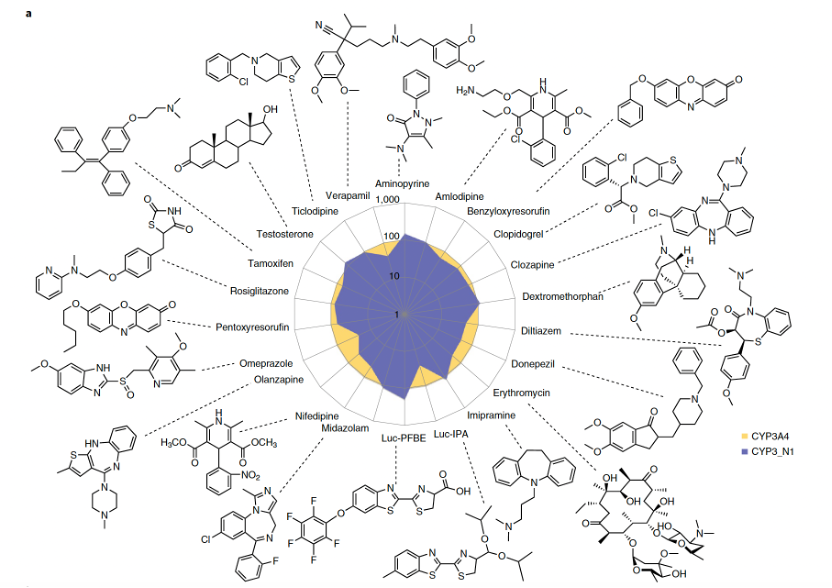
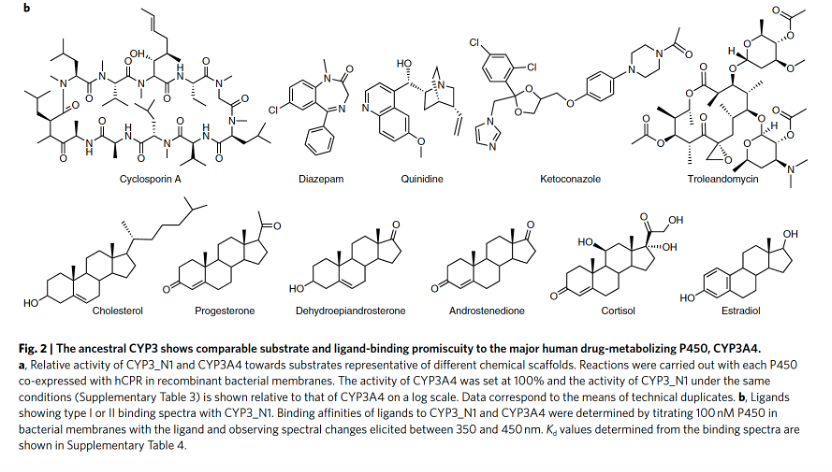
CYP3_N1表现出与人CYP3A4相似的催化活性
当将CYP3_N1的活性与特征明确的人类形式CYP3A4进行比较时,祖先酶对已知CYP3A4底物表现出相当或更高的活性(图2a)。基于配体结合到活性位点附近从而改变血红素辅基吸收光谱的能力的光谱结合研究表明,CYP3_N1与多种结构的配体相互作用,包括几种类固醇和大环内酯类抗生素(图2b),其亲和力与CYP3A4相当。祖先酶对睾酮的6β-羟基化和尼非地平的氧化(CYP3A4的标记底物)表现出更高的催化效率。因此,尽管对不同底物的相对活性(包括区域选择性)有所变化,但CYP3_N1显示出与人主要药物代谢P450相似的底物混杂性。
CYP3祖先酶通过疏水相互作用保持稳定
与每种现存的CYP形式相比,CYP3_N1在残基上差异为139–169个。尽管在整体氨基酸组成上没有出现主要趋势,但在CYP3_N1中不同疏水残基的比例发生了轻微变化。因此,我们假设增强的热稳定性是由于疏水核心包装得更好。在CYP3_N1中确定了9个替换,这些替换似乎增加了折叠特定区域中的有利非共价相互作用。当将CYP3_N1中的每个候选残基突变为降低疏水性的残基时,每个变化都显著降低了热稳定性。当这9个残基全部组合突变(在SDM8中)时,10T50降低到了一个与CYP3A4无显著差异的值。然而,将反向突变引入CYP3A4并未提高热稳定性,并且在几种情况下,消除了突变体的异源表达。这一结果强调了在具有内在不同上位性景观的蛋白质中理性设计热稳定性的难度。
CYP3_N1是进一步定向进化的稳健模板
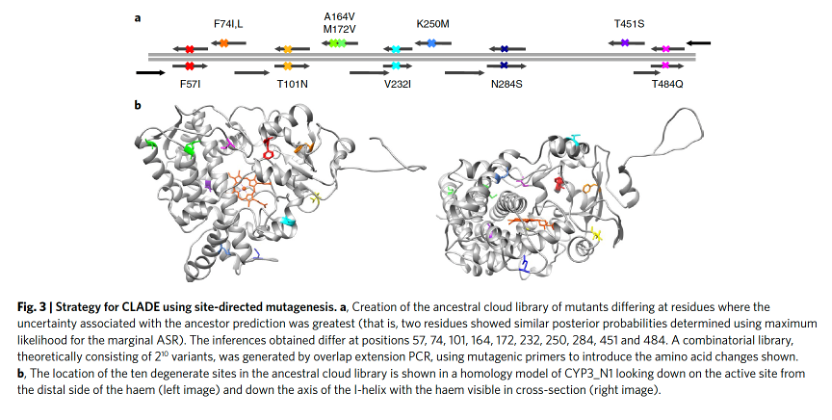
通过祖先序列重建(ASR),无法确定哪个序列是历史上正确的,而只能推断出最可能的祖先。因此,为了确定推断祖先CYP3具有热稳定性的稳健性,我们突变了10个位点(图3a、b),这些位点与祖先预测相关的不确定性最大(即,两个最可能的残基具有相似的后验概率)。实际上,我们生成了一群可能存在于第一批脊椎动物中的祖先蛋白质(以下简称CYP3祖先云库)。这群可能的祖先也代表了一组突变体,它们可能具有不同的表型,从而可以从中选择出最适合给定定向进化实验的祖先。对于生物技术目的而言,表型的稳健性不太重要,因为更多的表型多样性可能对蛋白质工程有用。
突变体被设计为编码一个羧基(C)端融合的增强型黄色荧光蛋白(eYFP)。在所采样的1023个突变体中,77%能够正确翻译,这由细胞中eYFP的荧光所证实。这些翻译的突变体在大肠杆菌中均表达了可检测的P450,平均产量为每升培养物550 ± 270 nmol(平均值 ± 标准差)。虽然大多数突变体的热稳定性相当,但有222个突变体的热稳定性显著高于CYP3_N1,其10T50值高达75.8 °C。对一组突变体针对典型CYP3A4底物的筛选揭示了不同的活性特征,表明该文库具有功能多样性。
祖先CYP3酶表现出增强的溶剂耐受性
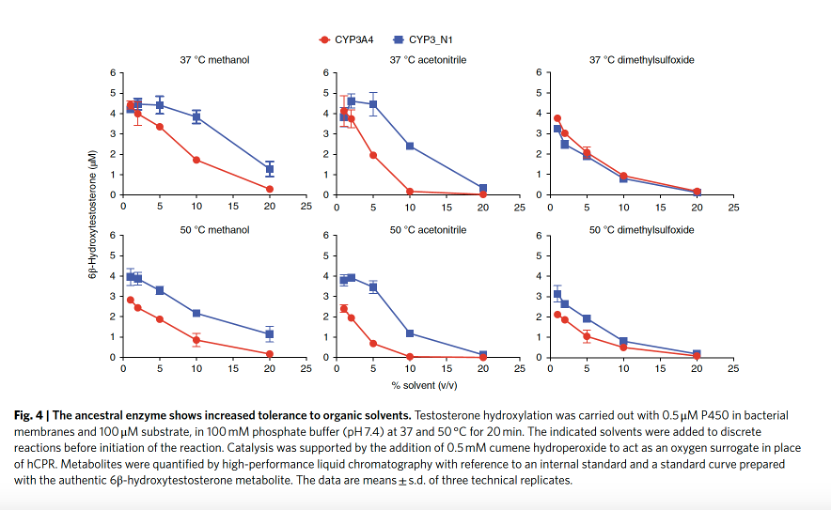
通过增加底物负载也可以提高时空产率。对于许多药物等亲脂性底物而言,能够在有机溶剂存在下进行生物转化反应是有益的。为了在不受人类NADPH-细胞色素P450还原酶(hCPR)干扰的情况下测试溶剂对CYP3_N1的影响,反应由氧替代物枯烯过氧化氢支持。与CYP3A4相比,CYP3_N1对甲醇和乙腈的耐受性显著增加,但对二甲基亚砜的耐受性没有增加(图4)。在37 °C下,与甲醇和乙腈分别以10%(v/v)的浓度处理时,CYP3_N1分别保留了91 ± 7%和63 ± 6%的活性,而CYP3A4仅保留了39 ± 2%和4 ± 2%的活性。
该方法适用于其他蛋白质家族
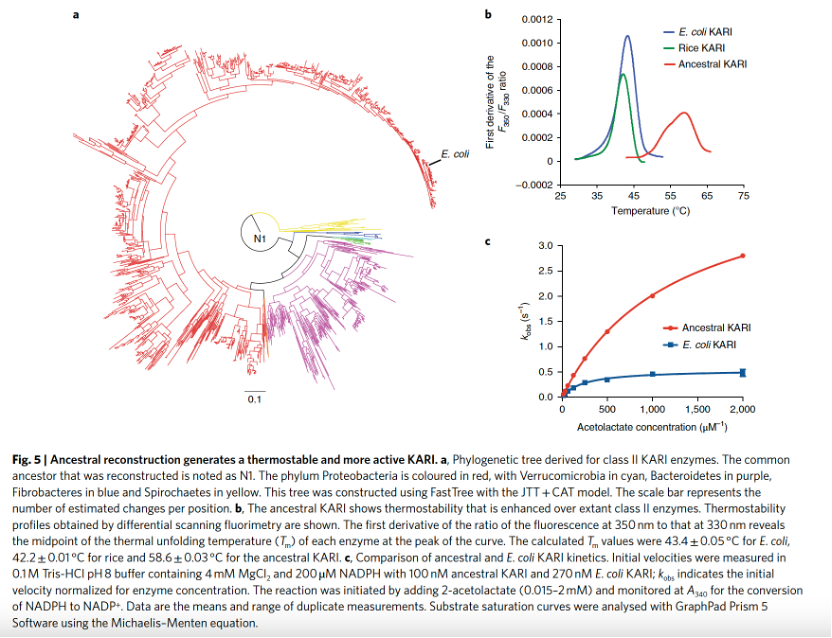
为了证明该方法的通用性,我们接下来研究了一种功能上不相关的酶——酮醇酸还原异构酶(KARI),该酶可将2-乙酰乳酸转化为2,3-二羟基异戊酸或将2-乙酰-2-羟基丁酸转化为2,3-二羟基-3-甲基戊酸(EC 1.1.1.86)。现有数据库中大多数II类KARI序列来自真细菌(特别是变形菌门和拟杆菌门),而来自疣微菌门、纤维杆菌门和螺旋体门的序列较少,因此我们推测了这些类群的共同祖先(图5a)。从该分析中选择的祖先II类KARI通过差示扫描荧光法测得的熔解温度(Tm)为59 °C,而大肠杆菌和水稻中现存的II类KARI的熔解温度分别为42–44 °C(图5b)。此外,该酶对2-乙酰乳酸的转换数(kcat)和比活性比大肠杆菌中的现有形式高8倍,尽管其米氏常数(Km)比大肠杆菌中的形式高5倍(图5c)。在不同温度下孵育10分钟后测定活性所得的半失活温度(10T50)也有所提高,然而,与大肠杆菌中的酶相比,祖先酶的最适温度和异丁醇耐受性并未增强。
(李俊锋摘译)
文章链接:https://www.nature.com/articles/s41929-018-0159-5#Sec2
高颜值免费 SCI 在线绘图(点击图片直达)
最全植物基因组数据库IMP (点击图片直达)
往期精品(点击图片直达文字对应教程)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|















































 1966
1966

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









