






任务执行中拖延的认知神经机制
引言
【执行意愿---->行为】虽然动机的产生往往是人们行动的前提(Ajzen, 2011;林琳&白新文,2014),但动机的产生却并不必然地导致随后的行为(Rhodes&deBruijn,2013;Sniehotta, Scho1z, & Schwarzer, 2005)。动机向行为的转换依赖于一系列自我控制、情绪调节和动机的管理的作用(Adam & Fayolle, 2015; Araujo Soares,M cIntyre, & Sniehotta, 2008; Reyes Fernandezet al, 2015)。其中任何一个因素的作用不足,个体的动机都可能无法引起随后的行为,从而引起拖延行为(Rozental & C arlbring, 2014; van Hooft, Bom, Taris, Van der Flier, & Blonk, 2005)。【研究表明高低拖延者之间的差异更多地来自于任务的执行阶段】此外,研究2的结果也显示,高低拖延者之间在决策阶段并没有展示出显著差异。这也间接地显示高低拖延者的差异更多地来自于任务的执行阶段。
【任务厌恶在拖延中的作用】如何应对任务引起的负性情绪是避免拖延所需要解决的关键问题。情绪调节理论最早开始强调应对负性情绪在拖延中的重要作用(FSirois&Pychyl,2013, D.M.Ticeetal,2001)。同样地,时间决策模型也强调只有当任务厌恶的作用强于结果效用的作用时个体才会选择拖延(S. Zhang & Feng, 2019; S. Zhang, Liu, et al,2019)。例如,个体应对负性情绪的能力越差,其拖延的可能也越高(Mirzae1,Gharraee, & Birashk, 2014, Rebetez, Rochat, & Van der Linden, 2015)。相反,如果个体能够接纳自己的负性情绪则能够减少自己的拖延行为(FM.Sirois,2014c,Wohl, Pychyl, & Bennett, 2010)。同时,通过训练提升个体的情绪调控能力也能够长久地减少拖延倾向(Eckert, Ebert, Lehr, Sieland, & Berking, 2016)。然而,个体是如何在任务执行的过程中调控负性情绪的仍不清楚。根据时间决策模型,个体执行任务时执行任务时所体验到的负性情绪要远远高出做决策时的预期。更强的负性情绪也需要更强的情绪调节能力来应对,也更容易导致个体的拖延。因此,相对于决策阶段,高低拖延者的差异也更有可能来自于任务的执行阶段。为了检验这一可能,实验5在保留实验3的决策阶段的基础上,进一步探索个体在兑现决策的执行过程中的差异。相对于实验3,实验5通过自适应程序根据不同个体自身的能力将任务难度始终控制在困难程度。同时,实验5将每次任务的奖赏固定。因此,任务的难度和奖赏在个体间能够完全保持一致,而个体在任务选择或执行上的差异也能够完全归结于个体特质间的差异。
生活中,人们往往需要在逆境中坚持很久才能实现自己的目标。因此,理解这--过程的认知神经机制对人们达成目标至关重要。虽然已有大量研究探索了个体在面对困难有奖赏的任务时的选择(Wunderlich,Rangel,&O'Doherty,2009, Yanget al, 2014), 但个体执行任务时的神经机制却仍不清楚。导致这一 研究现状的原因是缺乏合适的实验在实验室中模拟这一过程。例如,大部分的研究都通过增加要求的按键次数或者推动操作感所需的力度来充当困难任务(Chang et al, 2019; Culbreth, Moran, & Barch, 2018, Fillaetal, 2018)。因此难以模拟现实生活中的心理逆境。另一方面,先前研究还显示当人们不愿意执行某任务时,往往会选择从事更多的娱乐活动(Schouwenburg & Groenewoud, 2001;Tsukayama,Duckworth,&Kim,2012)。为了系统地模拟生活中个体面对厌恶任务时的选择,实验6同时探讨个体在面对任务厌恶时在娱乐活动和任务执行中的差异。
实验5 高低拖延者在任务执行中的差异
实验方法
被试
本实验招募了26名(14名女,平均年龄20.34)西南大学在校学生以验证不同拖延特质的个体在任务选择时的差异。该研究获得了西南大学伦理委员会的授权,并且所有被试都签署了知情同意书。
实验设计
本实验将个体的拖延特质作为自变量,将个体在拖延决策上的差异以及个体执行任务的差异作为因变量。从而探索具有不同拖延特质的个体是否会在拖延决策以及任务执行中表现出差异。本实验同样使用纯拖延量表作为拖延特质的测量工具。同时,本实验改编了实验4的拖延决策任务,使之不仅能够衡量个体在做决策的差异还更方便测查决策后执行任务的差异。因此,本实验既能够重复检测实验4中拖延决策与拖延特质之间的关系,又能够在此基础上进一步检验拖延特质与拖延决策以及任务执行的关系。
实验任务
与实验4相似,本实验同样要求被试在简单任务(数图片任务)与困难任务(数点任务)之间进行选择。而与实验4的区别在于,本实验只关注个体在高难度任务中的表现。因此实验中的数点任务将始终维持在高难度水平,并以此探究高低拖延者在任务执行中可能的差异。同样地,本实验也使用了纯拖延量表(PPS)来测量个体的拖延特质。
为了更好地衡量个体在对简单任务与困难任务.上的偏好差异,本实验操纵了简单任务与困难任务之间奖励的差异以探测个体选择的变化。本实验固定了困难任务的难度和奖赏大小,而不断地变化简单任务的奖励大小。当简单任务的奖励从0不断增大时,个体选择困难任务的概率也随之增大。当个体选择简单任务和困难任务的概率--致时(即概率各为50%),此时简单任务的积分便被称为无差比点。如果个体的无差别点越大,那么该个体便越能够坚持选择困难任务。
本实验的简单任务与实验4的设置相同,依然要求被试数出一定时间内呈现过的图片数量。正确完成该任务便可获得对应的积分奖励。错误完成该任务不会受到任务惩罚。简单任务的初始积分的空间为[0204060],四种奖赏梯度各有一个试次。四种奖赏出现的顺序随机,完成一个循环(四个试次)后程序将根据被试的选择变化积分空间中积分的大小。具体而言,如果四种奖赏中被试有一半以上选择了简单任务,则积分空间中所有积分都将被减少20;相反,如果简单任务的选择不足一半,其奖励积分都将被增加20;当选择比例恰好是一半时,积分空间不做改变。简单任务的奖励积分最大可达180,最少则为0.
与实验4相似,本实验仍然使用数点任务作为困难任务。数点任务的积分奖励固定为200。当被试正确完成任务时可以获得对应的积分奖励。由于本实验的数点任务错误时并不会伴随惩罚,因此个体可以在执行数点任务中主动放弃任务,使本实验更能够检测个体在任务执行中的差异。为了确保数点任务始终处于高难度水平,实验程序将每隔四个试次根据被试数点的正确率更改点图的范围。初始的点图范围为67~82,先前数据显示多数被试在这一范围内的正确率都为30%左右,因此保证了初始时任务的难度。如果四个试次中,被试正确地数出了2次及2次以上。实验将自动增加点图的难度(点图范围下限+7,点图范围.上限+5)。相反,如果被试正确的次数不足一次,实验则将自动减少点图的难度(点图范围的下限-5,点图范围的上限-7)。由于实验程序能够根据被试的表现调整点图的难度,本实验能够在考虑个体数点能力的同时保证数点任务始终处于高难度的水平。
本实验仍然一共含有48个试次,被试最后的报酬将由其在实验中获得的积分总和转换而成。
结果
由于实验中简单任务的积分奖励会根据被试的选择更改,本实验只运用个体简单任务的无差别点作为其选择偏好的衡量指标。为了探索个体简单任务的无差别点是否与PPS存在关联,本使用计算了两者之间的皮尔逊相关值。结果显示,两者的相关系数并不显著(r=-0.03,p>025)。 为了减少极端数据的影响,本研究也计算了spearman's ρ作为参照。结果显示,简单任务的无差别点与PPS之间仍然没有显著相关(r=0.01,p> 0.25)。这一结果重复了实验3的结果,意味着高低拖延被试在简单和困难任务的选择上并没有表现出显著的差异。
接着,本实验继续探索了高低拖延被试在困难任务的表现上是否有差异。本实验使用了个体在数点任务中的正确率作为衡量个体任务表现的指标。为了探测其与拖延特质的关系,本实验计算了两者的皮尔逊相关值。结果显示,个体在PPS上的得分与其在困难任务中的正确率呈显著的负相关(r=-0.60, p <0.01)(如下图)。这意味着,高拖延特质的个体在执行任务时表现地比低拖延特质的差。
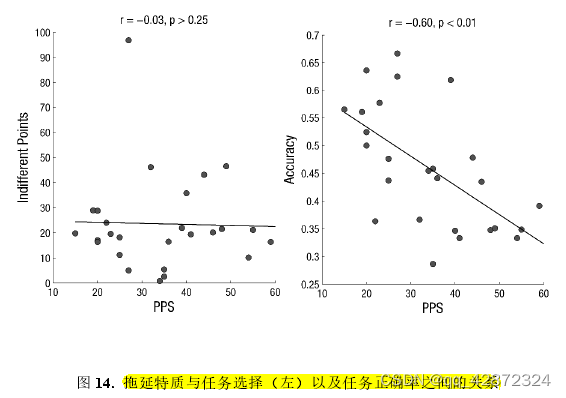
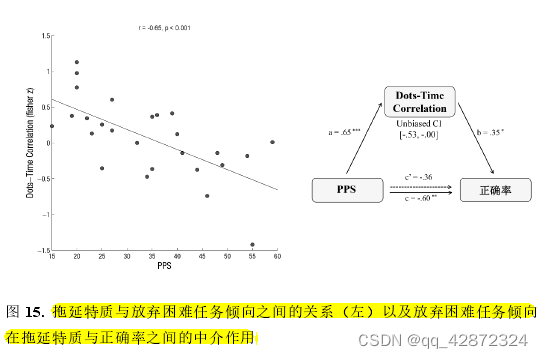
由于本实验会根据个体的正确率对任务难度进行动态的调整,因此高低拖延者之间的正确率差异不应该是由个体数点能力导致的。例如,正确完成数点任务时高低拖延所需的平均时间并没有显著差异(r=-0.03,p > 0.25)。 为了寻找导致高低拖延者正确率差异的原因,本实验将探索高低拖延者随着点数增多时在点图上所用时间的差异。针对每个被试,本实验计算了点图点数与所花费时间的皮尔逊相关值作为衡量两者关系的指标。按常理而言,点图点数越多,被试若想正确完成数点任务所需要花费的时间也应该相应增多。因此,点图点数与数点所用时间理应为正值。然而,出人意料的是,部分高拖延个体中点数与时间的相关为负值,并且个体的PPS得分越高,相关值(fisher z-transfromed)越小(r= -0.65,p< 0.001)。这意味着随着点数的增加,高拖延个体相对于低拖延个体更早地放弃了数点任务。由此可见,高拖延随点数增加更早放弃这一倾向很可能是导致高拖延个体正确率较低的原因。为了对此验证,本研究检验了点数和用时的相关值对PPS与正确率的中介作用。结果显示,点数与用时的相关值的中介作用显著(Biased Corrected CI= [-0.53,-0.00],如下图)。这一结果显示,高拖延个体在任务上较差的表现是由于其更容易放弃任务导致的。
讨论
该研究显示,高低拖延者在任务的选择上仍然没有显著差异。这一结果重复了实验4的发现,-定程度上说明了个体在决策上的差异对拖延特质贡献并不显著。此外,研究发现随着任务难度的增加,高拖延个体投入到任务中的时间比低拖延个体减少的更快。这-结果说明,高拖延个体较低拖延个体更倾向于在任务执行过程中放弃,从而在任务中表现也更差。进一步的中介分析显示,个体放弃的速度能够显著地中介拖延特质对正确率的作用。综上可知,高低拖延者都能够正确选择远期利益最大化的任务,但高拖延者会比低拖延更快地在任务中放弃,从而导致更差的任务表现。
本实验重复了拖延特质与拖延决策之间没有显著关系这一结果。然而,需要注意本实验仍然无法排除研究犯II型错误可能。虽然如此,从相关值大小而言,实验4和5所发现的拖延特质与拖延决策之间的皮尔逊相关值都不足0.2.因此,两个实验都未能为拖延决策与拖延特质之间存在关联提供有力的证据。与之相比,个体在任务执行中的差异则更能够解释个体拖延特质上的差异。这一结果一方面支持时间决策模型对实时任务厌恶和延迟任务厌恶的区分。即实际执行任务时个体所能够体会到的任务厌恶一定会强于个体在执行前的决策中所体验到的。因此,更能够使不同拖延特质的个体表现出差异。另一方面,这一结果也为设计干预拖延的训练方案提供了有益的参考。由于拖延特质更多地表现在执行任务中对任务厌恶的调节能力,因此以此为训练核心干预措施应该会比以增加执行意愿的干预措施更有效果。这也能够进--步解释为何以增强个体执行动机的干预措施都未能减少个体的拖延行为(Gustavson&Miyake,2017,Morin,2013),而以情绪调节为核心的干预措施反而能更有效地减少拖延行为(Eckert et al , 2016; Scent & Boes, 2014)。
此外,实验4和5中所检测的决策阶段是在任务执行之前所做的决策。虽然执行任务前的决策对随后的行为有着-定的预测作用,但并不意味着人们在随后的任务执行中便不再做出决策。现实情况下,人们极可能会在任务执行任务的过程中重新考虑是否继续执行任务,从而做出新的选择。这说明,虽然实验4和5并没有发现拖延特质与拖延决策之间存在显著关系,但这一结果并不否定时间决策模型用拖延决策这一概念解释拖延的观点。时间决策模型中用于解释拖延行为的决策是更为广义的概念上的决策。根据动机和意图是产生决策和行为的前提(Ajzen,2011),我们有理由认为拖延行为是伴随着做出拖延决策而产生的。因此,时间决策模型更倾向于将人们的决策看作是-一个动态的过程。即个体能够感觉当前的感受动态地变化自己的决策。从现实应用而言,在时间上距离行为越近,个体的决策也就越能够解释其行为的产生。而这一点也在时间决策模型中有着充分地表现:个体对未来的决策往往会随着时间的逼近而改变(S. Zhang & Feng, 2019)。
实验5清楚地揭示了高拖延个体为何会比低拖延个体在困难任务有着更差的表现。通过个体在任务执行中所用的时间可见,高拖延个体更早地结束了任务的执行。值得注意的是,高拖延个体更早地结束任务并不是由于其能够更快地完成任务所致。正常而言,任务量的增加会使个体花费更多的时间来完成任务。这一点在低拖延个体中得到了印证,即任务量上升其对应的执行任务的时间也相应地上升。然而,结果显示高拖延个体在任务量上升的情况下所用的时间反而会更少。据此可以推测,高拖延个体结束任务时并非顺利完成了任务,而是主动地提前放弃了任务。而且,高低拖延者的这一差异都是在同样选择执行任务的情况下发生的。这更印证了任务执行对于保证个体避免拖延行为的重要作用。因此,探明高拖延者放弃任务时的认知机制如何也十分必要。
实验6 拖延特质影响任务执行的神经机制
实验方法
被试
本实验招募了43名(30名女,平均年龄21.32)西南大学在校学生以验证不同拖延特质的个体在任务选择时的差异。该研究获得了西南大学伦理委员会的授权,并且所有被试都签署了知情同意书。
实验设计
在探测行为变量之间的关系时,本实验将任务的难度作为自变量将个体选择娱乐项目的时间及个体的任务表现以作为因变量,而将个体的拖延特质作为调节变量。从而检测当任务难度,上升时,高低拖延个体在选择娱乐项目的时间以及任务表现上的差异。而在探测行为变量与神经活动之间关系时,本实验预计将任务难度、娛乐活动的时间以及任务表现等作为自变量,而将神经活动作用因变量。同样以拖延特质作为调节变量,从而探索行为变量在高低拖延个体之间引起的神经活动的差异。本实验将拖延特质作为调节变量而非自变量的原因在于个体的任务表现以及神经活动既受到任务难度又受到个体特质的影响。因此,将拖延特质作为调节变量能够方便检验当行为变量--定时拖延特与因变量之间的关系。
实验任务
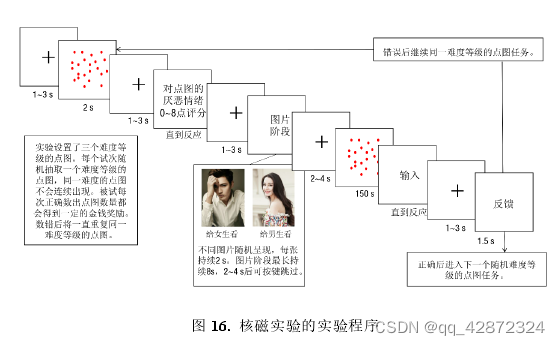
本研究旨在考察高低拖延者面对令人厌恶但却不应该回避的任务时在娱乐事件选择和任务表现上的差异(如下图)。该实验利用数点任务作为令人厌恶但却不应该回避的任务。为了使数点任务能够引起厌恶感,本研究为点图设置了高中低三个等级,并保证对每个被试而言完成高难度数点任务都是十分吃力。此外,本研究为每次正确数出点图设置了一定额度的金钱奖励。完成不同难度的点图的奖励额度相同,被试最后的报酬取决于其累计正确数出点图的个数。为了使被试认为即使数点任务很难也不应该回避,实验被设置为一旦在某一 难度错误后,被试将重复数同一难度的点图直到正确。由于实验的总时长是一定的(1小时),因此即使高难度的点图令人厌烦,被试也应该尽快完成,否则将无法继续在实验中获取金钱收益。其次,本研究使用欣赏人物照片作为娱乐事件。为了使欣赏人物图片对于大部分被试都有一-定的吸引力,本研究根据质性调查从网络和收集了大量外貌具有高吸引力的人物照片。并且在实验前邀请了6名异性恋志愿者(3男,3女)对图片进行了筛选。最后,实验中保留了332张女性图片,314张男性图片。其中女性图片作为男性被试的刺激材料,而男性图片作为女性被试的刺激材料。为了弱化个体间偏好差异的影响,欣赏图片时不同照片随机呈现,每张图片持续2s。这一设置保证了即使当前呈现的图片对被试没有吸引力,被试也有理由相信下一张可能出现自己喜欢的图片。
每个试次包含点图呈现阶段、欣赏图片阶段和数点阶段。在点图呈现阶段,被试只需要对点图的厌恶情绪进行评估即可。在数点阶段,被试有150s的时间数点。数点期间被试可以在任意时刻按键进入作答界面。数点任务设置了三个不同的难度水平。为了给每个被试设置合理的点图。每个被试在扫描前都进行了预实验。难度水平的设置是根据个体在预实验中数出接近正确答案(输入答案与正确答案差值的绝对值≤3个)的时间来确定的。这保证了不同个体在不同难度下正确输出点图点数所需的时间是大致相同的。在点图呈现阶段和数点阶段之间,被试有机会选择停留一会观看一些人物图片。欣赏图片阶段最多持续8s,经过一个最少图片时间(2-4随机抽取)后被试可按键跳过该阶段。因此在欣赏图片阶段时个体按键的频率(按键次数除以欣赏图片阶段的持续时间)可以反应个体在图片阶段停留的意愿。在该阶段个体的按键频率越高,停留的意愿就越低。
fMRI数据采集
本实验采用SiemensTrio3T磁共振扫描仪进行数据采集。使用圆形的极化头线圈( circularly pol arized head coil), 并在线圈中放入泡沫软垫以尽量减少被试的头动。使用T加权的平面回波成像序列( echo-planar imaging,EPI) 获得功能像,详细扫描参数如下: Repetton time= 2000 ms, Echo time= 30 ms, FOV= 200x200 mm2, fip angle=90°, matrix size= 64 x 64, voxel size: 3.1 x 3.1 x 3mm3,层数为33,层厚为3mm,层间距为0 mm。3D像(无间隔三维全脑)的扫描序列是梯度回波脉冲系列(DPGR), 扫描参数如下: Repetion time=2530 ms, Echo time=3.39 ms, FOV=256 x 256 mm2, fip angle=9o, matriz size=256 x 256, voxel size: 1.3x 1.0x 1.3 mm',层数为128, 层厚为1.33 mm,层间距为0mm。
fMRI数据分析
为了减少头动、低信噪比等问题的影响,本研究在进行正式的数据分析之前对数据进行了标准化的预处理。具体包括时间校正(Slice Timing)、头动校正(Realign,剔除前5个volume以保证数据的稳定性,然后以第一个volume为基准对齐减少头动)、图像标准化( N ormalization).空间平滑( Smooth, FWHM为8 mm).
由行为结果可知,拖延特质主要显著调节个体面临厌恶任务时的表现(例如:数点任务中的错误量)。因此脑成像的分析主要集中于探讨厌恶任务导致个体在数点任务中犯更大错的脑机制。以及拖延特质对这些脑机制的影响。为此,本研究以个体数点结束的时刻为感兴趣时刻,因为这一时刻的脑活动直接与个体形成的答案相关联。通过建立一-般线性模型( General linear model, GLM)探究(1)形成正确答案和错误答案的脑活动差异: (2)答案形成时刻,对点图点数有关联的脑活动:(3)答案形成时刻,对随后的错误量有关联的脑活动。
在个体间层面,本研究利用已经确认的个体在数点结束时刻的神经活动。利用Mediation Effect Parametric M apping (MEPM) (Wager, Davidson, Hughes,Lindquist; & Ochsner, 2008) (1) 搜索能够中介厌恶任务(点图点数)和任务表现(数点错误量)的脑活动; (2)以及这一大脑区域如何与其他区域互动最后决定个体的任务表现(数点错误量)。最后,为了探索受到拖延特质影响的脑活动,本研究检验了拖延特质对这些互动脑区之间的脑活动的调节作用。
结果
事后检查
为了检查数点任务是否能够成功地引起个体的厌恶情绪,本研究利用混合线性模型(mixedlinearmodel)预测个体在各试次中对点图厌恶情绪的评分。其中点图的个数作为固定效应(fixedeffect),而不同的个体则作为随机效应(random effect) 以控制不同个体之间的差异。结果显示,点图的个数能够正向地预测厌恶情绪的评分(b=0.05,t=41 .49,p< 0.001, R2=0.70)。这一结果显示,操纵点图的个数能够有效地引起厌恶情绪。点图中点的个数越多,点图所引起的厌恶情绪也就越强。
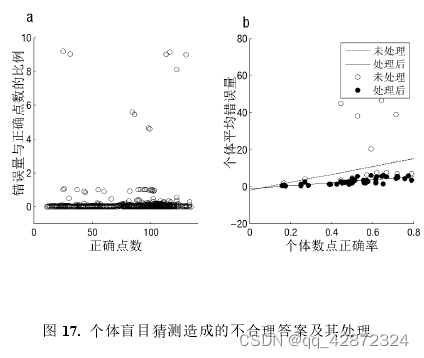
为了检验个体作答时是否有乱猜的情况,本研究检验了个体在不同试次中数点错误量的变化。数点的错误量是个体输入答案与正确答案差值的绝对值。数点错误量越大表示个体距离正确答案越远。虽然个体数点答案的错误量理应与点图的难度成正比。但按常理而言,个体的错误量与正确答案的比例应该在一定范围内。例如,行为结果显示95%以上的个体错误量与正确答案的比例都在20%以内。然而,部分被试的答案与正确答案比例却相去甚远。有的答案的错误量与正确答案的比例甚至在一倍以上(如下图a,例如:正确答案为100,输入答案为201)。由此可见,被试在输入答案时存在胡乱猜测的可能。为了减小这些乱猜数据的影响,本研究将所有错误量与正确答案的比例在20%以上的试次全部按照正确答案的20%取值。实践显示,处理后个体的平均错误量能比原始数据的平均错误量更好地拟合个体数点的正确率(如图b)。因此,对原始数据的错误量进行合理的处理更能很好地减少极端数据的影响,并且更好地描述个体的行为表现。
行为结果
为了探究不同程度的拖延个体在数点任务中表现的差异。本研究将个体的拖延程度作为调节变量,利用混合线性模型预测了个体数点任务错误量的大小。在该模型中,个体的拖延得分、点图的点数以及拖延得分与点数的交互项作为固定效应。而不同个体作为随机效应以控制个体间的差异。结果显示,拖延特质对点数和错误量之间的关系存在显著的正向调节作用(t= 3.28,p< 0.01)。即当点图中点数一-定时,高拖延的个体较低拖延的个体产生了更大的数点错误量。
为了探测高低拖延个体在点图点数一定时错误量的差异是否有可能源自个体所体验到的厌恶情绪或者所面临的点图点数不同,本研究继续验证了个体的拖延得分对点数和厌恶情绪的关系,以及点数和难度等级的关系之间是否有显著调节作用。为了验证当点图一定时,高低拖延个体所感受到的厌恶情绪是否有显著差异。本研究还探究了拖延特质是否能显著调节点图点数与厌恶情绪之间的关系。混合线性模型的结果显示:在预测厌恶情绪时,拖延特质与点图点数的交互项不显著(t=0.50, p=0.62)。这意味着当点数一定时,高拖延的个体所感受到的厌恶情绪并没有显著高于低拖延的个体。此外,为了验证高低拖延个体所面对的点图是否有差异。本研究使用混合线性模型利用个体的拖延得分、难度等级、拖延得分与难度等级的交互项预测了点图的点数。结果显示个体的拖延得分(t=-0.10,p=0.92) 以及拖延得分与难度等级的交互项(t= 1.51,p=0.13)对点图点数的预测作用均不显著。这一结果意味着高低拖延个体在不同难度所面临的点图并没有显著的差异。
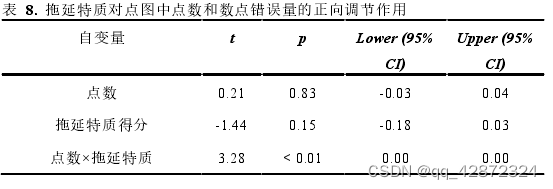
此外,为了探究不同拖延特质的个体在面对令人厌烦的任务时观看人物图片意愿的差异,本研究利用个体的拖延得分、点图点数以及两者的交互项预测个体在图片阶段的按键频率。其中,拖延得分、点图点数以及两者的交互项被纳入作为混合线性模型的固定效应,而不同个体被纳入作为随机效应以控制个体间的差异。结果显示,拖延特质对点数和按键频率有着正向调节的趋势。这显示,当点数一定时,高拖延特质的个体比低拖延特质的个体按键次数更高,在欣赏图片阶段的停留意愿越低。
脑成像结果
本研究探索了个体在形成正确答案和错误答案时的神经活动差异。结果显示,个体在形成错误答案时比形成正确答案时在的双侧前脑岛(anteriorinsula)、左侧杏仁核(amygdala)、扣带回前部及中部(antenior, middle cingulate cortex, ACC,MCC)、以及背外侧前额叶( dorsal lateral prefrontal cortex, DLPFC)等脑活动有着显著升高(p < 0.05, FWE-corrected)。其中前脑岛及杏仁核往往负责情绪的加工和调节(Shah, Klumpp, Angstadt, Nathan, & Phan, 2009; Stein, Simmons, F einstein,&Paulus,2007),而背外侧前额叶区域则更多地负责认知控制(Soltaninejad,Nejati,& Ekhti ari, 2019; Steinbeis, Bemnhardt, & Singer, 2012)。 因此,这- -结果预示着在个体在形成错误答案时可能既经历着厌恶情绪又进行着对厌恶情绪的认知控制过程。
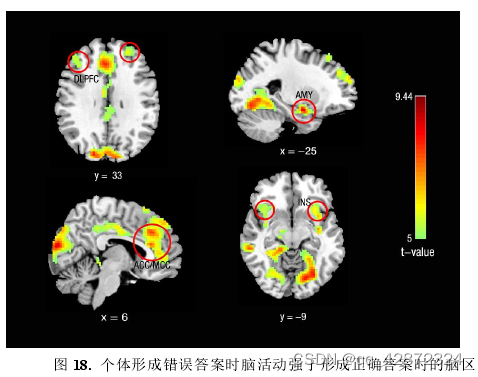
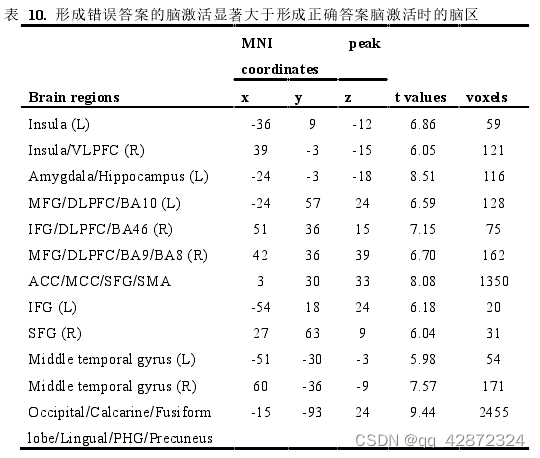
为了进一步确认答案形成时刻,脑活动受到厌恶任务的难度(点图点数)的脑区以及与随后的任务表现(错误量)存在关联的脑区。本研究利用parametric modulation分析,分别以点图点数以及错误量作为调节参数。结果显示,脑活动受到点图点数调节的脑区只有左侧前脑岛(p < 0.005, cluster size > 40; uncorrected)。而脑信号受到错误量调节的脑区则包括前脑岛、杏仁核、海马、中扣带回以及背外侧前额叶等(q < 0.05, FDR-corrected)。
为了进一步确认厌恶任务(点图点数)引起个体任务错误(数点错误量)的脑区,本研究采用Medi ation Effect Par ametic M apping (MEPM)对形成错误答案与正确答案的contrast进行了全脑搜索,找寻答案形成时刻神经活动能够显著中介二者关系的脑区。这一结果显示,脑活动能够显著中介厌恶任务和任务错误的脑区有:双侧前脑岛、双侧梭状回(fusiformgyrus).双侧枕小叶(occipitallobe)、双侧缘.上回( supramarginal gyrus, SMG)以及大量的双侧中央前回( precentral gyrus)、中央后回(postcentral gyrus)和辅助运动区(supplem entary motor area, SMA)(p < 0.05, cluster size > 20, uncorrected).其中,可能由双侧前脑岛负责表征任务所诱发的厌恶情绪,从而引起随后的任务错误。这一推测的理由有:(1)以往研究中显示前脑岛能够表征恶心感、厌恶情绪以及疼痛(Huetteletal,2006;P1oghaus etal,1999;Wickeretal,2003)等多种个体期望回避的刺激:(2)本研究的parametric modulation 分析显示与脑活动受到厌恶任务的难度(点图点数)调节的区域只有前脑岛部分: (3)而受到任务错误(数点错误量)调节的区域也包含了前脑岛部分。因此本研究将右侧前脑岛(如下图,a*b/c=0.37) 作为任务厌恶感的表征区域,继续探讨其他脑区间是如何与前脑岛互动从而产生任务错误的。

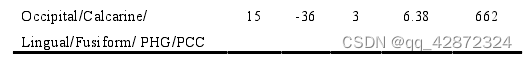
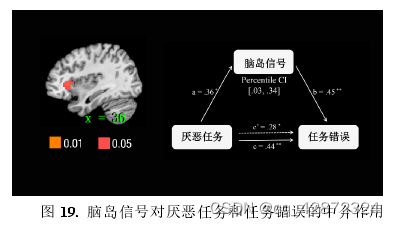
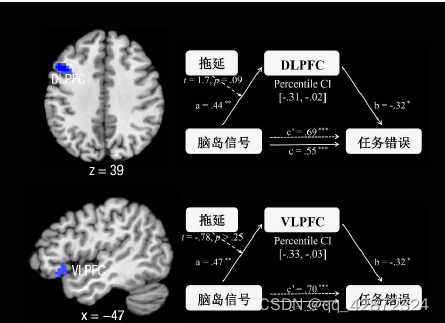
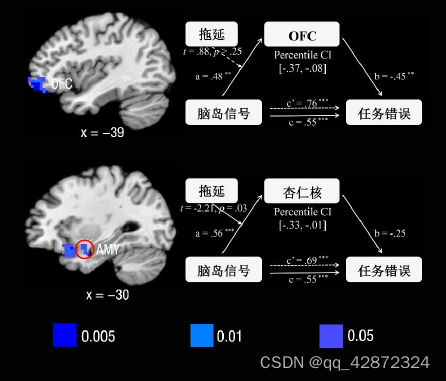
为了探究前脑岛如何与其他脑区相互作用从而引起任务错误(数点错误量),本研究继续利用MediationEffectParametricMapping技术对全脑进行搜索寻找能够中介前脑岛信号与任务错误的脑活动。结果发现一个包含额中回(middle frontal gyrus, MFG)和背外侧前额叶(DLPFC) 能够显著地负向中介脑岛信号对任务错误的作用。--个包含左侧杏仁核、腹侧前脑岛延申至腹外侧前额叶(ventral lateral prefrontal cortex, VLPFC)的团块也起着显著地负向中介作用。此外,左外侧眶额叶( orbitofrontal cortex, OFC)、双侧缘上回(SMG)、双侧中央旁小叶(paracentrallobule)及其延伸至辅助运动区域(SMA)也都起着显著的负向中介作用。另--方面,双侧的中央前回(precentralgyrus)及部分的辅助运动区域则起着显著的正向中介作用。
由于背外侧前额叶(DLPFC).腹外侧前额叶(VLPFC).眶额叶皮层(OFC)以及杏仁核都是进行情绪调节的核心区域(Kadosh et al, 2016; M orawetz, Bode,Derntl,&Heekeren,2017)。因此这些脑区所起到的负向中介作用很可能是对前脑岛信号进行情绪调节的过程,从而降低了数点任务的错误量。为了寻找与拖延特质存在关联的脑区间的相互作用,本研究分别检验了拖延特质在这些脑区与前脑岛关系之间的调节作用。结果发现拖延特质只对脑岛与杏仁核之间的关系存在负向调节作用(t=-2.21, p=0.03)。

讨论
该研究显示,高拖延个体较低拖延个体在面对令人厌恶的任务时,确实有着更差的表现,这一结果也与实验5的发现吻合。然而,在面对更加令人厌恶的任务时,高拖延个体并没有比低拖延个体更久地停留在娱乐活动中。甚至从趋势而言,当面对更加令人厌恶的任务时,高拖延个体相对于低拖延个体会更倾向于跳过娱乐阶段。从神经机制而言,当个体产生任务错误时较没有产生错误时在双侧前脑岛、左侧杏仁核、扣带回前部及中部、以及背外侧前额叶等脑区有着更多的脑活动。并且,其中只有前脑岛的脑信号既受到任务错误量也受到任务难度的调节。进一步的中介搜索也显示,前脑岛的信号能够中介任务难度对任务错误量的作用。探索脑区间的相互作用发现,任务执行时有两个神经系统影响着个体任务的表现。其中一个是一前脑岛为中心的厌恶情绪系统,负责表征任务厌恶,从而加剧任务中的错误量。另一个系统包括DLPFC, OFC, VLPFC以及杏仁核的情绪调节系统,负责抑制负性情绪,从而减弱前脑岛对任务错误的加剧作用。并且,在这些厌恶情绪脑区和情绪调控脑区的相互作用中,个体的拖延特质会负向调节前脑岛和杏仁核之间的相互作用。换言之,相较于低拖延个体,高拖延个体的前脑岛和杏仁核的互动效率更低,从而增加了任务的错误量。
本研究首次揭示了任务执行中的神经机制。由于实验范式的局限,先前的研究只能通过简单的厌恶刺激来探究对应的神经机制(deJongetal,2019;Vander Weele et al, 2018)。这些刺激的形式相对于真实的任务执行过于简单,因此往往难以模拟执行任务的场景。另外,虽然许多研究使用了厌恶图片和视频来探究个体情绪调节的神经机制(Kanske Heissler, Schonfelder, Bongers, & Wessa, 2010,Kohn et al, 2014), 但却又缺乏任务执行过程和指标。相比之下,实验6则很好地创设了一个令人厌恶却又应该去做的任务。相比于简单的厌恶刺激,本实验不仅能够引起足够的厌恶情绪,也能够更多地还原个体执行任务的过程。此外,与简单的情绪调节任务相比,本研究能够进一步地揭示情绪调节脑区与任务表现的关系。虽然先前的情绪调节能够揭示情绪调节的脑区如何减少杏仁核等情绪脑区的神经活动(Banks, Eddy, Angstadt, Nathan, & Phan, 2007; Zotev, Phillips, Young,Drevets,&Bodurka,2013),但这一发现还不足以描述人们执行令人厌恶的任务时的复杂情景。例如本研究的发现并不简单地表现为调节脑区对情绪脑区的减弱作用。具体而言,在执行厌恶任务时,升高的前脑岛信号会伴随着更多的情绪调节脑区的活动。然而,虽然两个脑区的信号同时升高,但他们对任务表现的作用却完全相反。情绪脑区的活动增加任务错误,而调控脑区则减弱这一作用。
本研究有利于揭示高低拖延个体在神经机制上的差异。由于缺乏实验室实验,大多先前的研究只能通过rest-statefMRI或者大脑灰质体积等指标来探讨高低拖延个体的神经差异(Chen, Liu, Zhang, & Feng, 2019; Huetal, 2018, W. Zhang et al, 2016)。由于无法直接检测这些脑区在引起拖延时起到的功能,这一方法只能通过寻找其他具有中介作用的人格特质来解释这些脑区的作用(PLiu&Feng,2017, 2018,C:.Zhangetal,2017)。相比之下,本研究则能够直接模拟人们在任务中拖延的情景,也能够因此直接检验拖延行为中发挥作用的脑区。例如,先前的研究虽然发现个体的拖延特质与DLPFC以及VLPFC等脑区有关(Chenetal,2019,Y Wu et al, 2016),却无法知道这些脑区直接的相互作用及功能。本研究则发现DLPFC及VLPFC等脑区能够直接与情绪相关脑区(如脑岛)发生拮抗作用,从而决定个体的任务表现。同时,本研究还进一步揭示了拖延特质在脑区间的关系,以及这一-关系与任务表现的关联。例如,本研究首次发现拖延的特质会影响脑岛与杏仁核之间的互动效率。
研究总结
研究3则进--步探讨高低拖延个体在做出执行任务的决策后.执行任务时认知神经机制的差异。首先.实验5探究了高低拖延个体在任务选择以及执行中的差异。结果仍然显示,高低拖延个体都能够更多地选择困难但有奖赏的任务。但是在任务执行的过程中.高拖延个体却比低拖延个体有着更差的表现。由于实验程序能够通过自适应的方式向每个个体提供难度相当的任务.因此他们任务表现的差异可以归结于任务中的重新选择。与该推测一致.结果显示,相对于低拖延个体.高拖延个体在任务执行的过程中会更早地放弃任务的执行。并且个体放弃任务的快慢能够显著地中介掉高低拖延个体在任务表现上的差异。实验6则在这一发现上进一步探讨了高低拖延个体在任务执行上的神经差异.以及他们是否在娱乐活动的选择上也存在差异。实验6的结果重复了实验5的发现.即随着任务难度的上升.高拖延个体会比低拖延个体在任务中表现得更差。但高拖延个体并没有比低拖延个体更倾向于停留在娱乐活动之中。在神经机制上而言,实验6首次揭示了个体在坚持执行令人厌恶却又有必要做的任务时的神经机制。简言之,实验6发现有两个相互拮抗的神经系统影响着个体是否能够正确地完成任务。其中一个是负贵表征任务厌恶的以前脑岛为中心的情绪系统。另一个系统则负责抑制负性情绪。其中包括DLPFC,OFC,VLPFC以及杏仁核等脑区。情绪系统的活动上升.会引发更强的自我控制情绪的需求.从而引起自我控制脑区的信号上升。总体而言,自我控制系统和情绪系统在影响任务表现上存在着拮抗作用。情绪系统的活动增加会使个体在任务中有更差的表现.而自我控制系统的激活则会尽可能地缓和情络系统的这一作用。此外,在情绪系统和自我控制系统的作用中拖延的特质会影响杏仁核与脑岛的作用效率。具体而言.相对于低拖延者。高拖延者的脑岛会引起更少的杏仁核的活动。从而导致更差的任务表现。
综合而言,本论文以时间决策模型的拖延决策为核心。从决策产生前的任务评估、拖延决策以及任务执行三个阶段系统地考察了对拖延行为存在作用的重要因素。结果发现:(1)在决策前的任务评估阶段,任务厌恶和结果效用是构成任务主观价值的主要任务因素,并且以各自独立的神经环路影响着拖延行为;(2)在拖延决策的决策阶段.自我控制特质能够调节任务厌恶或者结果效用的权重来影响决策和拖延行为。然而。拖延特质却并没有对任务执行前的拖延决策产生显著的影响; (3) 在决策后的执行阶段.尽管高拖延者和低拖延同样选择了执行任务,但却更倾向于提前放弃任务的执行,从而呈现出更差的任务表现。并且存在着包含DLPFC、VLPFC、OFC以及杏仁核自我控制系统通过抑制前脑岛的活动从而避免高拖延个体产生更差的任务表现。此外.高拖延个体较低拖延个体的杏仁核调控前脑岛的效率更低,因而导致了高拖延者的任务表现更差。综合而言,这些结果不仅在理论上为时间决策模型提供了系统的支持。也能够从应用上为开发减少拖延行为以及个体的拖延倾向的干预手段提供指导。
讨论
任务执行中拖延的认知神经机制
研究3在研究2的基础上进-步探讨任务执行的具体过程中产生拖延行为的认知神经机制。虽然执行动机是行为产生的前提(Montano&Kaspzyi,2015).但并不是所有的动机和决策都会转化为完整的行动。研究2的实验3也证实了这-点.即高拖延的个体在决策时并没有比低拖延个体更多地回避任务的执行。因此.探讨个体在任务执行中认知神经机制是减少拖延行为的必要环节。为此,研究3的实验5探讨了高低拖延者在任务选择和随后的执行中的差异。结果显示,高低拖延者在对任务进行选择时仍然没有表现出个体间的差异.但是高拖延个体却比低拖延个体更早地放弃了任务的执行。并且这一差异也使得高拖延个体在任务中有着更差的表现。这一结果不仅重复了实验3的发现。也进一步地揭示出高低拖延者的个体差异更多地表现在任务的执行中。实验6则进一步地探索了高低拖延者在执行任务时的神经机制.以及他们的差异。结果显示,有两个相互拮抗的神经系统影响着个体任务执行的好坏。其中一个是以前脑岛为核心的情绪系统。这一系统的激活会使个体更想要回避任务。从而阻碍任务的完成。另一个是包括DLPFC、VLPFC、 OFC以及杏仁核的情绪调控系统。这一系统负责约束由前脑岛负责表征的负性情绪,从而使任务更好地完成。从个体差异的角度而言,结果发现个体的拖延得分越高,其杏仁核与脑岛的相互作用的效率也越低。
实验5首次系统地揭示了高低拖延个体在决策和任务执行中的差异。由于拖延行为的复杂性。先前的研究都未能设计出实验室实验来模拟拖延的过程(Steel & Kingseck, 2016; Steel, Svartdal, Thundiyil & Brothen, 2018)。 相比之下。实验5不仅区分了任务的选择和执行阶段.还进-步地揭示了高低拖延者在任务执行中的差异。与实验3相似。实验5也未能发现高低拖延者在决策上的差异,而只发现高低拖延者的差异体现在任务的执行中。这一结果重复了原来研究所发现的高低拖延者在意图层面没有显著差异的结果(Stee,2007)。另外,这一结果也体现了时间决策模型中对实时效用与任务延迟效应进行区别的意义(S. Zhang, Lin etal, 2019)。由于延迟的任务厌恶会受到时间折扣的影响,因此总是小于实时的任务厌恶。这解释了为何个体预期将会执行某事,但却仍在那一时刻到来时放弃。
另外,由于实验5的任务执行阶段能够区分高低拖延者.因此该实验有望成为一个拖延行为的客观测量方法。如今流行的拖延测量方法主要还是基于问卷的自我报告法,例如Lay (1986)的一般拖延量表(Ge ne ral Procrastination scale . GPS)、McCown and Johuson (1989)的成人拖延问卷、Tuckma (1991)拖延问卷(Tuckman proc rastination scale)以及Steel (2010)的- - 套纯拖延量表(Pure Procrastination Scale, PPS)。 此外,虽然也有研究利用个体完成论文写作的时间、阅读完指定书籍的时间以及完成间卷并通过邮件返回的时间(Anely&Wertenbroch 2002; Glick & Orsllo, 2015; McCrea etal, 2008)作为拖延的测量指标。这些实验又无法排除生活中多种因素的影响。相对于先前的实验和指标,实验5通过实验模拟创造了一个独立的实验环境.因此可以排除日常环境中复杂的干扰因素。同时,实验5还能够通过自适应的方式为被试提供合适难度的任务。这一优势一方面方便了实验的操作。另一方面能够完全匹配个体间的难度。可以更具纯粹地探索个体间差异的影响。
实验6则在实验5的基础上进一步探索了任务执行过程中的神经机制。实验6揭示了脑区直接是如何互动以使个体能够坚持执行令人厌恶但却具有长远利益的任务。一方面,实验6重复了实验5的结果,即当任务的难度上升时,高拖延的个体比低拖延的个体更倾向于犯更大错误。而在神经机制上,个体犯错误时比正常完成任务时在许多脑区都有更强的活动,其中包括。双侧前脑岛(antenior insula)、左侧杏仁核(amygdala). 扣带回前部及中部( anterior,middle cingulate cortex, ACC, MCC)、以及背外侧前额叶( dorsal lateral prefrontal cortex, DLPFC)等。进一步的Mediaton Effect Parametric Mapping分析则更清晰地展示了这些脑区在影响任务表现时的互动机制。简言之,结果显示确实存在一个厌恶情绪脑区和一个情绪调控脑区相互拮抗来影响个体任务的表现。这一结果也为时间决策理论的思想提供了证据。即拖延与否是受到短时情绪引起的回避需求和远期利益引起的控制需求向竞争所决定的。
实验6的结果不仅有助于理解拖延的神经机制,还能为拖延的理论建构提供启发。近些年来.研究主要应用静息态功能性磁共振(functio nal mag netic re sonance imaging, fMRI)以及脑形态学指标(voxel-based mophonetry,VBM)来探索与个体的拖延特质存在关联的大脑自发性代谢以及大脑灰质体积。这些研究表明与个体的拖延特质存在关联的脑区主要集中在海马旁回(prahippcampalgyrus,PHG)和前额叶皮层(prefrontal cortex, PFC) (Hu Lin, Guo, & Feng,2018;P. Liu & Feng,2017; W. Zhang, Wang, & Feng, 2016)。例如。Chenetal. (2019)根据大脑形态学的结果将与拖延有关的脑区分成了自我控制、情络调节以及预期想象三个大的网络。由于缺乏实验室实验对拖延情最的模拟.这些研究都无法从认知机制上验证这些脑区的功能。而本研究则首次揭示了拖延行为至少与情绪调控系统以及情绪系统的拮抗作用有关。这--定程度上为脑网络的划分以及神经模型的建立提供了依据。其次。该结果对拖延的理论思考也有着启发。从个体差异的层面而言,实验6只发现高拖延与负责情绪调控的杏仁核抑制脑岛信号的效率有关.而并没有发现负贵认知控制的DLPFC等脑区与脑岛之间的作用存在关联。这一定程度上说明。高拖延个体的特质差异可能来自于情绪调节网络的功能减弱而非认知控制的能力低下。
综上可述。任务的执行阶段确实能够更多地反映和揭示高低拖延者之间的差异。一方面,这一结果体现了任务执行阶段在减少和干预拖延行为的重要作用。另一方面.这个阶段中拖延个体所展现出的差异有助于研究者更深入的理解拖延产生的认知神经机制。















 446
446











 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









