









Next generation sequencing in predicting gene function in podophyllotoxin biosynthesis
下一代测序在预测鬼臼毒素生物合成基因功能中的应用
P450Rdb: CYP450数据库--地表最强系列--文献精读24-CSDN博客
P450催化的联芳基偶联反应-文献精读72_chemshell dimer优化过渡态-CSDN博客
近十年天然产物药物的生物合成研究进展-文献精读33_雌四醇合成工艺-CSDN博客
木脂素的生物合成及其微生物法生产的研究进展-文献精读47_木质素过氧化物酶在大肠杆菌中表达-CSDN博客

背景:结构复杂的植物药物的生物合成途径尚不完整或未知。
结果:通过对鬼臼属植物组织进行下一代测序/生物信息学和代谢组学分析,发现了在鬼臼毒素生物合成中的潜在未知基因。
结论:识别出特异性区域的甲基二氧桥形成的环氧化物酶(CyP450),催化了松脂酚的形成。
意义:提供了多个药用植物转录组组装和代谢谱数据,供科研界使用。
关键词:细胞色素P450,酶催化,代谢,代谢组学,转录组学,木脂素,甲基二氧桥形成,植物药物,鬼臼毒素
摘要
鬼臼属植物是(−)-鬼臼毒素的来源,这是一种用于半合成各种强效且广泛应用于癌症治疗药物的芳香四氢呋喃木脂素。然而,其生物合成途径仍然大部分未知,最后明确证明的中间体是(−)-罗汉松脂素。在本研究中,对鬼臼属植物的桃儿七和北美桃儿七的转录组进行了大规模平行测序,并对相应的组装结果进行了后续的生物信息学分析。首先通过与先前已知参与鬼臼毒素生物合成的多个基因序列以及其他候选生物合成途径基因的序列进行比对,验证了组装过程的正确性。本研究描述了其中两个基因的特征,即来自桃儿七的细胞色素P450酶CYP719A23和来自北美桃儿七的CYP719A24。这两种酶均能通过催化甲基二氧桥的形成,将(−)-罗汉松脂素转化为(−)-松脂酚,并且对其他测试的潜在底物无作用。有趣的是,这些酶与生物碱生物合成中的甲基二氧桥形成酶非常相似,而与木脂素生物合成酶更相似的候选酶在所用底物上表现出催化不活跃。因此,这一整体策略使得更容易识别可能参与(−)-鬼臼毒素生物合成的酶,并强调了下一代测序和生物信息学在探究和推断药用植物生物合成途径中的推理能力。
引言
大规模平行测序技术正在迅速发展,并日益提供前所未有的机会,显著增强对生物合成过程的理解,包括对一些重要且尚未充分了解的(非模式)药用植物的研究。一大优势是,这些技术有可能缩短新基因发现的时间框架,从而更迅速地改善我们对代谢过程的理解,例如,与更传统的方法(包括标记前体的施用、潜在中间体的鉴定、酶的纯化与表征、基因克隆、表达序列标签(EST)文库等)相比。在这种背景下,几项近期的研究已经使用这些大规模平行测序技术研究了多种非模式植物,转录组的组装主要是通过454和Illumina测序的数据生成的。其中包括人参(Panax quinquefolius L)、中国人参(Panax ginseng)、五叶参(Gynostemma pentaphyllum)、蝴蝶兰(Phalaenopsis orchids)、茶树(Camellia sinensis)、长春花(Catharanthus roseus)、罂粟(Papaver somniferum)、金合欢(Acacia auriculiformis)、花生(Cicer arietinum)以及冷杉(Abies balsamea)等。尽管通过这种方式可以获得大量的信息,但仍需要通过有见地的分析来帮助筛选候选基因,并仔细确定它们是否具有特定的生物合成功能。
鬼臼属植物产生芳香四氢呋喃木脂素(−)-鬼臼毒素(1b),由于其在半合成抗癌药物如替尼泊苷(2)、依托泊苷®(3)和依托泊苷(4)中的广泛应用(见图1),具有重要的药用价值。后者是拓扑异构酶II抑制剂,广泛用于治疗多种癌症,包括肺癌和睾丸癌。然而,作为(−)-鬼臼毒素(1b)的主要来源,桃儿七(鬼臼属植物)被大量采伐,一些报告指出,由于过度采伐,它已成为濒危物种。
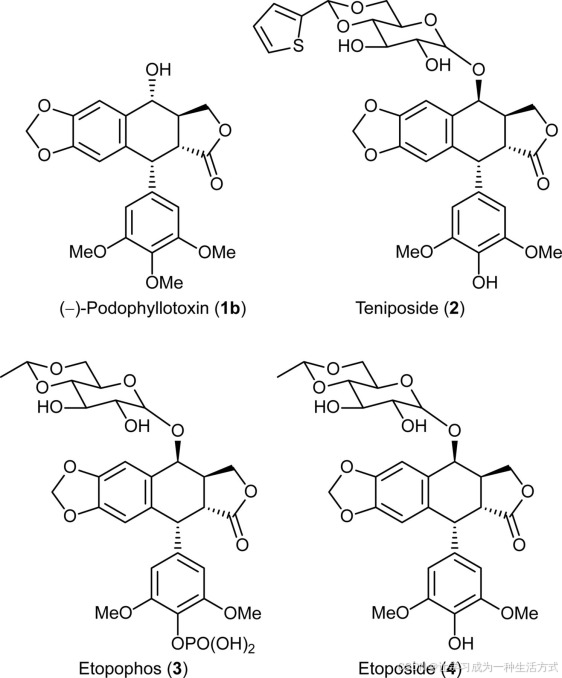
(−)-鬼臼毒素(1b)及其衍生物替尼泊苷(2)、依托泊苷(3)和依托泊苷(4)在癌症治疗中的应用。
尽管已有多种合成化学方法被描述用于合成(−)-鬼臼毒素(1b),但通过这些途径生产该化合物的经济性较差(15–17)。一种可能更具生产力的替代方法是通过生物技术手段在细胞培养或整株植物中获得更大量的(−)-鬼臼毒素。然而,目前由于我们对(−)-鬼臼毒素(1b)生物合成途径的了解尚不完整,这一方法仍不可行。尽管如此,在木质单体形成后,其生物合成途径的起始点是通过两个E-松柏醇(5)衍生的自由基的立体选择性耦合反应,涉及导向蛋白的参与,从而生成(+)-松脂素(6a)(见图2)。该化合物随后通过松脂素/松柏醇还原酶的作用进行对映选择性还原,依次生成(+)-松柏醇(7a)和(−)-去氢异松柏醇(8b)。接下来,通过立体特异性的脱氢反应,将后者转化为(−)-罗汉松脂素(9b),这是已知的(−)-鬼臼毒素(1b)生物合成的最后一个明确步骤(18,21)。随后,利用鬼臼属植物和亚麻属植物对这些已知步骤进行了验证(20,22)。另一方面,关于将(−)-罗汉松脂素(9b)转化为(−)-鬼臼毒素(1b)的下游推测步骤,仅通过粗酶法进行了报道;至今尚未鉴定出相关基因或纯化出这些酶(23–27)。
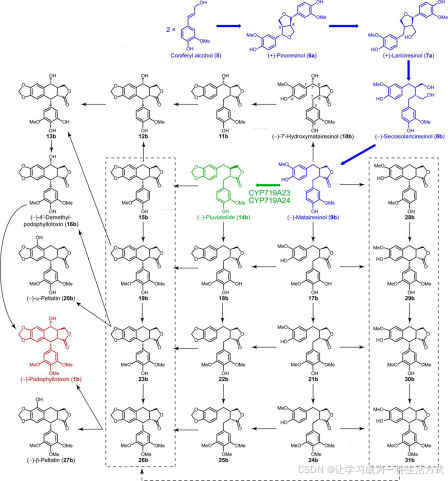
可能的(−)-鬼臼毒素(1b)及相关木脂素的生物合成途径和/或网络。已知的生物合成步骤用蓝色突出显示,而本文描述的由CYP719A23和CYP719A24催化的反应则用绿色标示。
本研究描述了使用Illumina技术进行转录组测序和生物信息学分析结合代谢组学分析的策略,旨在促进(−)-鬼臼毒素(1b)生物合成中基因的快速发现。具体而言,这一策略导致了在桃儿七和北美桃儿七中发现了两个新基因,这些基因编码能够通过将(−)-罗汉松脂素(9b)转化为(−)-松脂酚(14b)催化甲基二氧桥形成的酶。
实验程序
植物材料
桃儿七和北美桃儿七植物分别来自Digging Dog Nursery(位于加利福尼亚州Albion)和Companion Plants(位于俄亥俄州Athens),并在华盛顿州立大学的温室设施中培养。
化学品
(−)-罗汉松脂素(9b)(28)、(−)-牛蒡子苷元(34b,图3)和(+)-连翘脂素(38a)(29)从连翘(Forsythia intermedia)中分离得到。(±)-松脂素(6a/b)(22)、(±)-7′-羟基罗汉松脂素(10a/b)(18)、(±)-7-羟基罗汉松脂素(32a/b)、(±)-异牛蒡子苷元(33a/b)(29)和(±)-薄荷醇(37a/b)(30)按照已描述的方法合成。(−)-α-铁杉脂素(36b)和(−)-5-甲氧基罗汉松脂素(35b)由Eric P. Swan博士(Forentek)赠送,而(−)-α-和(−)-β-盾叶鬼臼素(20b和27b)则由Paul M. Dewick博士(诺丁汉大学,英国)提供。(−)-鬼臼毒素(1b)购自Sigma。
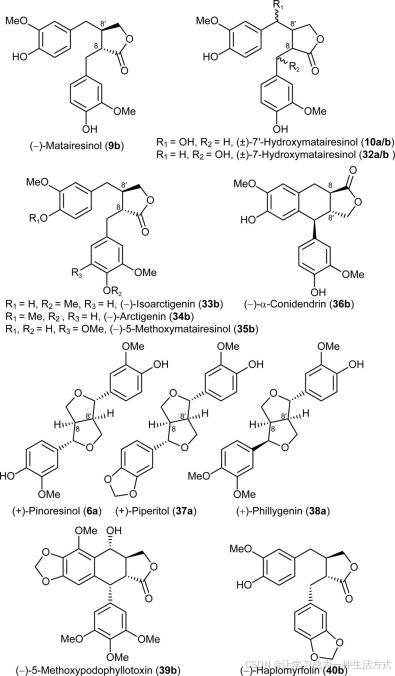
作为候选底物在CYP719A23和CYP719A24甲基二氧桥形成及(−)-haplomyrfolin(40b)分析中的木脂素测试。
代谢物提取与分析
从根茎、茎和叶(2克,鲜重)中分别采样,立即用液氮冷冻,研磨成细粉,然后进行冻干处理。每个组织经过150 μm筛网筛选后,用10 μl/mg的甲醇-水(7:3,v/v)溶液提取,所得提取液保持在−80°C,直至分析。样品通过液相色谱分析,使用Waters Acquity超高效液相色谱系统,配备Waters BEH C18柱(1.7 μm颗粒,2.1 × 50 mm),在280 nm处检测,并采用正离子模式的电喷雾质谱分析(表1)。梯度程序如下:流速为0.3 ml/min,水相中含0.1%甲酸和乙腈中含0.1%甲酸,线性梯度从95:5变为75:25,在11分钟内,接着在5分钟内变为60:40,再在4分钟内变为0:100,最后保持1.5分钟在0:100。柱温保持在25°C,样品注入体积为5 μl。质量通过Waters Xevo G2 Q-TOF质谱仪测定,使用亮氨酸-脑啡肽作为锁质量标准。
Base peaks are in bold.
| Lignan | Molecular mass | ESI-MS m/z |
|---|---|---|
| Daltons | ||
| (−)-Podophyllotoxin (1b) | 414 | 437, 432, 415, 397, 247 |
| (−)-α-Peltatin (20b) | 400 | 423, 418, 401, 247 |
| (−)-ß-Peltatin (27b) | 414 | 415, 247, 203 |
| Podophyllotoxin-glucoside (41) | 576 | 599, 594, 397 |
| α-Peltatin-glucoside (42) | 562 | 580, 563, 409, 247 |
| ß-Peltatin-glucoside (43) | 576 | 594, 577, 415, 409, 247 |
| 4′-Demethylpodophyllotoxin (16) | 400 | 423, 418, 401, 383, 247 |
(−)-鬼臼毒素(1b) m/z 437.1215 ([M + Na]+, 81%),计算值437.1207;432.1652 ([M + NH4]+, 25%),计算值432.1653;415.1391 ([M + H]+, 36%),计算值415.1387;397.1285 ([M + H − H2O]+, 100%),计算值397.1282;247.0605 (19%),计算值247.0601。
(−)-α-盾叶鬼臼素(20b) m/z 423.1059 ([M + Na]+, 60%),计算值423.1050;418.1503 ([M + NH4]+, 45%),计算值418.1496;401.1236 ([M + H]+, 71%),计算值401.1231;247.0608 (100%),计算值247.0601。
(−)-β-盾叶鬼臼素(27b) m/z 415.1397 ([M + H]+, 44%),计算值415.1387;247.0605 (100%),计算值247.0601;203.0708 (1%),计算值203.0703。
鬼臼毒素-葡萄糖苷(41) m/z 599.1739 ([M + Na]+, 61%),计算值599.1735;594.2181 ([M + NH4]+, 11%),计算值594.2182;397.1289 ([M + H − H2O − Glc]+, 100%),计算值397.1282。
α-盾叶鬼臼素-葡萄糖苷(42) m/z 580.2030 ([M + NH4]+, 3%),计算值580.2025;563.1763 ([M + H]+, 3%),计算值563.1759;m/z 409.1134 (33%),计算值409.1129;247.0603 (100%),计算值247.0601。
β-盾叶鬼臼素-葡萄糖苷(43) m/z 594.2183 ([M + NH4]+, 1%),计算值594.2182;577.1926 ([M + H]+, 11%),计算值577.1916;415.1393 ([M + H − Glc]+, 22%),计算值415.1387;409.1140 (10%),计算值409.1129;247.0599 (100%),计算值247.0601。
4′-去甲基鬼臼毒素(16) m/z 423.1057 ([M + Na]+, 53%),计算值423.1050;418.1499 ([M + NH4]+, 37%),计算值418.1496;401.1229 ([M + H]+, 14%),计算值401.1231;383.1134 ([M + H − H2O]+, 100%),计算值383.1125;247.0606 (21%),计算值247.0601。
对于代谢物的相对丰度评估,通过提取每个化合物的特定离子色谱图进行积分:m/z 397.128用于(−)-鬼臼毒素(1b),m/z 247.060用于(−)-α-和(−)-β-盾叶鬼臼素(20b和27b),m/z 594.218用于鬼臼毒素-葡萄糖苷(41),m/z 409.113用于α-盾叶鬼臼素-葡萄糖苷(42),m/z 577.193用于β-盾叶鬼臼素-葡萄糖苷(43),以及m/z 383.113用于4′-去甲基鬼臼毒素(16)(见图4中的结构)。
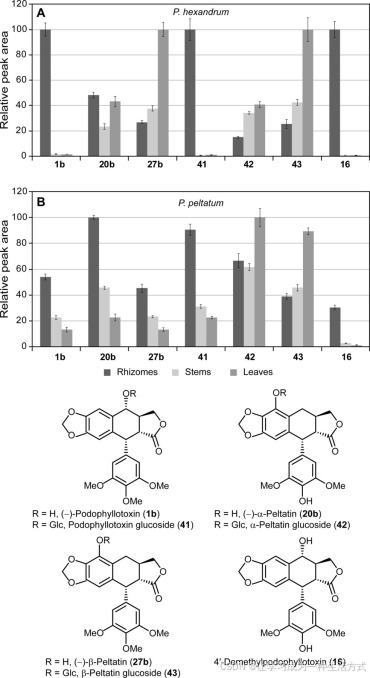
不同组织中桃儿七和北美桃儿七的相对木脂素含量 在桃儿七(A)和北美桃儿七(B)组织中鉴定的木脂素包括(−)-鬼臼毒素(1b)、(−)-α-盾叶鬼臼素(20b)、(−)-β-盾叶鬼臼素(27b)、鬼臼毒素-葡萄糖苷(41)、α-盾叶鬼臼素-葡萄糖苷(42)、β-盾叶鬼臼素-葡萄糖苷(43)和4′-去甲基鬼臼毒素(16)。每个木脂素的相对峰面积(280 nm)以最丰富的木脂素为100%,其余木脂素则以该值的百分比报告。
RNA提取与cDNA制备
总RNA分别从100 mg的快速冷冻根茎中使用Invitrogen Plant RNA Purification Reagent提取,从茎和叶中使用Qiagen(加利福尼亚州瓦伦西亚)RNeasy Mini Kit提取,按照生产商的说明,包括附加的Qiagen清理协议。每个样品提取的1 μg RNA随后用于cDNA制备。经过DNase I(Invitrogen)处理后,使用SuperScript® III First-Strand Synthesis System(Invitrogen)根据制造商的说明进行cDNA合成,并用于候选基因的扩增。
转录组测序与文库组装
总RNA样品(约25 μg)溶解于水中,通过使用Bioanalyzer 2100(Agilent,CA)评估其完整性,RNA完整性数值(RIN)>5.0的样品进一步处理。RNA样品浓度使用Qubit荧光计中的RiboGreen试剂进行估算,随后使用Illumina RNA-Seq或Illumina TruSeq RNA样品制备试剂盒,按照Illumina协议处理。简而言之,使用oligo-d(T)25磁珠(Dynabeads;Invitrogen)从总RNA样品中分离出poly-A+ RNA,然后使用提供的试剂进行片段化。随后使用随机六聚体和提供的酶混合液进行第一链cDNA合成。经过第二链cDNA合成后,使用T4 DNA聚合酶和大肠杆菌DNA聚合酶I的Klenow片段修复cDNA片段的末端,并添加单个去氧腺苷到磷酸化的平末端片段的3′末端。接着使用噬菌体T4连接酶连接测序接头,再通过琼脂糖凝胶电泳分离出约500 bp的片段。每个文库DNA通过Qiagen QiaQuick凝胶提取试剂纯化,并进行15轮PCR扩增。扩增后的文库使用Bioanalyzer 2100和Nanodrop ND-1000(Thermo Scientific)分别评估质量和数量。根据Nanodrop浓度,文库被标准化至10 nm,并通过定量PCR使用已知浓度的参考文库确定可测序分子浓度。流动池在Illumina Cluster Station上准备,使用Illumina Genome Analyzer IIx仪器获取成对的54 bp序列读数。
初始读取集被检查是否存在明显异常(如由于文库制备问题导致的过度代表性读取)和已知的测序伪影,特别是φX和已知的Illumina接头。然后,读取集被划分为大约一百万个读取的集合,并使用FASTX Toolkit在phrap Q = 10的水平上进行质量修整。
清理后的成对读取数据随后使用ABySS(32, 33)进行多次组装,使用不同kmer(k)范围(24 ≤ k ≤ 54)并行运行。由此生成的Contig被命名为合成ESTs,随后通过使用不同的kmer大小进行多次组装,汇总生成的合成ESTs,并按下述方法使用标准EST组装器进行后续组装。通常,这会产生至少20组合成ESTs。
每个ABySS组装的Contig被使用ABySS scaffolder进行框架构建,利用读取配对约束。通过SOAPdenovo套件(34)的GapCloser解决插入的NNN间隙间隔,然后根据得到的至少80核苷酸的框架构建每个kmer的合成EST集合。所有读取数据都被纳入组装,最终生成整体转录本参考。
最终组装
合成的EST集合通过MIRA在EST组装模式下进行组装(35)。为了明确控制冗余(98%序列相似性),组装结果使用cd-hit进行处理(36)。最终,至少100个核苷酸的contig被报告为构建的最终contig集合。通过使用Tablet(37)检查堆积视图,对最长的contig以及读取数异常低或高的contig进行了手动评估。所有研究的组织和物种的最终组装可以通过Medplants网站访问。
生物信息学分析
用于香草酸、苯丙烯类和木脂素生物合成途径的参考基因的氨基酸序列来自NCBI数据库。大部分基因来自拟南芥(Arabidopsis thaliana),但以下基因除外:前香草酸氨基转移酶来自四季海棠(Petunia × hybrida),水杨酰CoA:香草酸水杨酰转移酶来自烟草(Nicotiana tabacum),导向蛋白来自连翘(Forsythia × intermedia),松脂素/松柏醇还原酶来自四季海棠(北美桃儿七),以及去氢异松柏醇还原酶和导向蛋白也分别来自北美桃儿七(表2和附表S1)。每个参考基因的氨基酸序列被应用于BioEdit(版本7.0.5.3,2005年10月28日)中的tblastn(38)进行同源基因搜索,数据库为桃儿七或北美桃儿七的contig数据库。选择了与每个参考基因具有较高同源性的contig(相似度≥30%,E值≤5 × 10−23),并使用NCBI的ORF Finder确定每个contig的开放阅读框(ORF)。每个ORF被单独转译成氨基酸序列,使用EMBOSS Transeq(欧洲生物信息学研究所)进行转译,然后将每个氨基酸序列应用于tblastn(NCBI),以确认其是否与目标基因相对应。上述过程旨在避免嵌合contig的歧义(39,40)。此外,使用blastp(NCBI)计算每个候选基因的E值和相似度(%),并与相应参考基因的氨基酸序列进行比较。基于已知序列的肉桂酸4-羟化酶(CYP73A1)、对香豆酰CoA 3-羟化酶(CYP98A44)、阿魏酸5-羟化酶(CYP84A3)、黄酮类6-羟化酶(CYP71D9)、黄酮类3′-羟化酶(CYP75A1)和可可托贝林合酶(CYP80G2),以及甲基二氧桥形成酶(例如薄荷醇/紫杉醇合酶CYP81Q1(41)和(S)-可可托贝林合酶CYP719(42)选择了(−)-鬼臼毒素(1b)生物合成的未知候选基因(附表S2)。
BLAST result for known protein sequences from *北美桃儿七* and *桃儿七*
Pp, 北美桃儿七; Ph, 桃儿七.
| Protein | Query accession number/species of origin | Species/transcript | Identity | E value |
|---|---|---|---|---|
| % | ||||
| Dirigent protein | AAK38666.1/北美桃儿七 | PpDir1_Pp27246 | 99.0 | 2 E−115 |
| PhDir1_Ph08051 | 92.0 | 4 E−93 | ||
| Pinoresinol/lariciresinol reductase | ACF71492.1/桃儿七 | PhPLR2_Ph140193 | 99.4 | 0 |
| PpPLR1_Pp37193 | 95.5 | 0 | ||
| Secoisolariciresinol dehydrogenase | AAK38664.1/北美桃儿七 | PpSDH1_Pp12640 | 99.3 | 3 E−163 |
| PhSDH1_Ph12248 | 97.5 | 2 E−152 |
基因克隆与酵母表达
从桃儿七的cDNA中使用附表S2中描述的引物扩增CyP450基因候选基因。扩增使用PfuTurbo DNA聚合酶(Agilent PCR)在热循环仪中进行,循环条件为:94°C变性30秒,55°C退火30秒,70°C延伸3分钟,最后延伸10分钟。PCR产物在1%的琼脂糖凝胶中分离,获得约1500 bp的单一条带。序列已提交至GenBank™数据库,登录号为KC110988–KC110998(附表S2)。
产物被克隆到pENTR/d-TOPO(Invitrogen)中,并根据制造商的说明转移到酵母表达载体pYES-DEST52(Invitrogen)中。从桃儿七中获得了酵母表达克隆pYES-DEST52::CYP719A23和pYES-DEST52::CYP73A107,从北美桃儿七中获得了pYES-DEST52::CYP719A24。每个克隆随后使用锂乙酸盐法按照载体制造商的说明,分别转化到Saccharomyces cerevisiae株系WAT11(43)中。空载体pYES-DEST52也被转化到WAT11中,用作阴性对照。
转化酵母的筛选
转化后的酵母使用缺乏尿嘧啶的合成完全培养基(SC-U)平板(2%琼脂和2%葡萄糖)进行筛选。转化酵母的单个菌落被接种到含2%葡萄糖的液体SC-U(10 ml)中,培养至A600达到3–4,然后接种到含有诱导培养基(200 ml SC-U,含1%鼠李糖和2%半乳糖)的1升锥形瓶中,最终A600为0.05。诱导反应在30°C的振荡器中以300 rpm的速度进行24小时,直到细胞被收集用于即刻的微体准备。
微体准备
诱导后,收获每个候选重组酶的细胞,通过3900 × g离心10分钟,重悬于3 ml/g细胞重量的Tris-HCl缓冲液(50 mm,pH 7.4),其中含有EDTA(1 mm)、山梨醇(600 mm)、DTT(0.1 mm)和PMSF(0.4 mm)。使用0.5 mm的玻璃珠(Biospec Products, Inc.)破碎细胞,将总量的一半加入到Falcon管(50 ml)中的细胞悬浮液中,通过高速振荡,每次30秒、间隔30秒,共10次,过程中保持冰上。细胞裂解液通过倾析与玻璃珠分离,玻璃珠用初始重悬细胞时所用缓冲液的半量洗涤两次。通过15,000 × g离心10分钟去除细胞残骸,并在91,000 × g下进行超离心75分钟,收集微体作为凝胶状沉淀。每个微体分级被重悬于Tris-HCl缓冲液(50 mm,pH 7.4)中,含有EDTA(1 mm)和30%甘油(500 μl/g新鲜细胞重),并使用Dounce匀浆器进行匀浆。微体准备可在−80°C保存,最长可保存8周,且无可检测到的活性损失。
酶学分析
在含有10 μl底物的甲醇溶液(浓度从0.1到25 mm不等)的磷酸钠缓冲液(200 μl,100 mm,pH 7.5)中进行反应,然后加入10 μl的NADPH(50 mm)溶液和30 μl的微体制备(蛋白浓度约为60 μg/μl)。每次加入微体制备后,反应混合物进行振荡,并在25°C下震荡孵育5分钟。反应通过加入冰醋酸(10 μl)终止,然后在16,000 × g下离心30分钟。上清液的样品随后使用前述的代谢物分析方法,通过超高效液相色谱进行直接分析。所有动力学数据均在三次独立实验中进行。动力学参数(Km和kcat)通过非线性最小二乘法数据拟合(44)估算,微体中的细胞色素P450(CyP450)含量通过还原CO差异光谱法(45)使用Lambda 20紫外-可见分光光度计(PerkinElmer Life Sciences)进行测定。
酶产物的分离
使用表达CYP719A23的酵母微体制备,并以(−)-罗汉松脂素(9b)为底物的150个酶学反应被汇集在一起,整个(约37.5 ml)的制备液用氯仿(40 ml)提取3次。合并的有机溶解物在真空中蒸发至干,并重悬于甲醇(1 ml),然后通过高效液相色谱(HPLC)纯化酶生成的(−)-松脂酚(14b)。使用SymmetryShield RP18柱(Waters,粒径5 μm,3.9 × 150 mm)进行洗脱,流速为1 ml/min,并采用水和乙腈的线性梯度,从9:1到4:6在25分钟内变化,再在2.5分钟内变为1:0,最后在4.5分钟内保持1:0。含有(−)-松脂酚(14b)的分馏液被汇集,冻干后,使用氘代氯仿作为溶剂,四甲基硅烷作为内标,在Varian VNMRS 600 MHz核磁共振仪上进行1H、13C和13C,1H异核单量子相干(HSQC)核磁共振分析(附表S3和图S1–S3)。
(−)-松脂酚(14b)
δH (CDCl3): 2.45–2.62 (4H, m); 2.89 (1H, dd, J = 7.0 and 14.1); 2.96 (1H, dd, J = 5.2 and 14.0); 3.85 (3H, s); 3.86 (1H, dd, J = 7.4 and 9.1); 4.11 (1H, dd, J = 7.1 and 9.2); 5.93 (1H, d, J = 1.4); 5.94 (1H, d, J = 1.4); 6.44–6.47 (2H, m); 6.63 (1H, dd, J = 1.8 and 7.9); 6.67 (1H, d, J = 1.8); 6.69 (1H, d, J = 7.7); 6.84 (1H, d, J = 8). δC (CDCl3) 178.64, 147.85, 146.65, 146.32, 144.52, 131.59, 129.43, 122.07, 121.55, 114.22, 111.48, 108.79, 108.31, 101.04, 71.19, 55.87, 46.59, 41.00, 38.30, 34.62. MS: m/z 379.1155 ([M + Na]+, 计算值379.1157), 357.1337 ([M + H]+, 计算值357.1338), 339.1230 ([M + H − H2O]+, 计算值339.1232), 161.0604 (计算值161.0603), 137.0604 (计算值137.0603) 和135.0445 (计算值135.0446)。
结果与讨论
代谢物谱分析
首先,进行了代谢物谱分析,以确保目标代谢物和生化相关代谢物在不同的鬼臼属植物组织中存在。因此,使用超高效液相色谱-电喷雾电离-质谱(UPLC-ESI-MS)对根茎、茎和叶的提取物进行了检测。根据代谢物的紫外吸收、保留时间和质谱,可以轻松检测并确认目标代谢物(−)-鬼臼毒素(1b)的存在。在桃儿七中,它在根茎中积累较多,但在叶子和茎中几乎无法检测到(图4A)。在北美桃儿七中也观察到相同的趋势,根茎中的积累较高,尽管茎和叶中的(−)-鬼臼毒素(1b)含量也接近根茎的水平(图4B)。通过与标准品进行比较,确认了该代谢物,其保留时间和质谱一致(表1和“实验方法”)。此外,在两个物种的不同组织中检测到了另外两个相关木脂素(−)-α-盾叶鬼臼素(20b)和(−)-β-盾叶鬼臼素(27b),并通过标准品进行了确认(表1和“实验方法”)。有趣的是,在桃儿七中,这两种木脂素的积累模式与(−)-鬼臼毒素(1b)的积累模式截然不同,其中(−)-α-盾叶鬼臼素(20b)在所有组织中都有检测到,而(−)-β-盾叶鬼臼素(27b)则在地上部分,特别是在叶片中积累较多(图4A)。而在北美桃儿七中,这两种木脂素的积累模式与(−)-鬼臼毒素(1b)类似,从叶片到茎的积累逐渐增加,根茎中的含量最高(图4B)。
根据质谱分析(表1和“实验方法”),还能够识别已知的(46–48)木脂素糖苷形式:鬼臼毒素-葡萄糖苷(41)、α-盾叶鬼臼素-葡萄糖苷(42)和β-盾叶鬼臼素-葡萄糖苷(43)。同时,还可以检测到4′-去甲基鬼臼毒素(16),其质量与同分异构体(−)-α-盾叶鬼臼素(20b)相同,但具有由于水分子丢失所导致的额外基峰[M + H − H2O]+,m/z为383.1134(计算值383.1125)。鬼臼毒素-葡萄糖苷(41)和4′-去甲基鬼臼毒素(16)的积累模式与鬼臼毒素(1)的模式相似,在地下组织中的含量较高,尤其是在桃儿七中。而对于(−)-α-和(−)-β-盾叶鬼臼素糖苷(42b和43b),它们在所有组织中都有积累,叶片中的水平略高(图4)。
根据观察到的(−)-鬼臼毒素(1b)及上述其他相关木脂素的代谢物谱,可以认为,这些木脂素的整体生物合成功能途径可能在两个物种的所有组织中存在;另一方面,特定的羟基化酶可能在不同组织中有选择性地加羟基,分别导致(−)-鬼臼毒素(1b)或(−)-α-和(−)-β-盾叶鬼臼素(20b和27b)的合成,这可能是组织特异性的。
转录组组装与分析
在确认并分析代谢物与标准品的比对后,RNA从组织中提取,并按“实验方法”描述生成转录组数据。从粗RNA中分离出poly-A+ RNA,进行片段化,并使用随机六聚体转化为cDNA,进行端配对,以提高数据质量以便后期组装。添加接头并对产品进行质量控制后,使用Illumina Genome Analyzer IIX进行测序,生成读取集。随后,检查这些初始读取集是否存在已知异常,如与φX基因组或Illumina接头参考集的显著序列相似性,发现的异常被丢弃,因此初始数据略微减少,通常为1–5%。然后,将读取数据分配到配对数据块,以利用工作流程中的并行处理,随后使用ABySS进行组装,这使得可以将读取数据的输入文件与其来源库关联。在此步骤中,kmer大小往往是构建和分析de Bruijn图中最敏感的因素,选择不同的kmer大小常常导致类似但不同的组装。来自这些组装的contig长度通常在100–500 bp之间,并被视为“合成ESTs”,然后将这些EST组装成最终的contig集合。值得强调的是,以这种方式处理大量数据会面临许多挑战,因此上述每个阶段都包括多个完整性检查(例如由系统故障引起的处理不完整)、基本数据质量测量(例如测序污染物的丰度)以及生物学意义。为桃儿七和北美桃儿七组装的转录组数据库分别生成了227,885和147,960个contig,包括若干完整和不完整的ORF。获得的转录组数据可与其他重要药用植物的数据一起访问,用户可以通过Medplants网站获取。
接下来,进行了代谢物谱和转录组组装数据的比较分析,分析了各个组织(根茎、茎和叶)中的数据,以:(a) 验证之前从北美桃儿七克隆的已知基因是否正确组装;(b) 识别在香草酸-香叶酸途径到苯丙氨酸、苯丙烯类途径入口点以及通向单木酚类的核心苯丙烯途径中的候选基因,例如松柏醇(5)(后者也是(−)-鬼臼毒素(1b)生物合成途径的入口代谢物);(c) 进行生物信息学分析,识别可能的候选基因,这些基因编码在(−)-罗汉松脂素(9b)之后的步骤,最终形成目标化合物(−)-鬼臼毒素(1b)。
因此,首先对组装结果进行了检查,以评估与先前描述的基因相对应的contig是否存在,并获得组装质量的初步衡量。在我们之前关于(−)-鬼臼毒素(1b)生物合成的研究中,我们已经克隆并表征了一个导向蛋白,该蛋白负责介导E-松柏醇(5)的立体选择性耦合,生成(+)-松脂素(6a)(18),以及一个去氢异松柏醇还原酶,负责将(−)-去氢异松柏醇(8b)转化为(−)-罗汉松脂素(9b)(22)。当使用这些已知序列对北美桃儿七的组装结果进行查询时,观察到与先前描述的基因具有非常高的相似性(>98%)和低的E值(<10−110)(表2)。这表明,组装过程提供了与已知基因相关的高质量数据;然而,是否完全正确(例如,是否有点突变等)并未进一步探讨,即没有通过克隆、蛋白质表达等进行确认。
在香草酸/苯丙烯类和单木酚类形成途径中进行同源基因搜索也取得了成功,识别出了在两个物种中与每个基因相似的多个同源基因。这包括初始香草酸/香叶酸途径中的10个酶和核心苯丙烯途径中的9个酶(附表S1),它们的相似度≥30%,E值≤5 × 10−23,如“实验方法”所述。总体而言,对于香草酸/苯丙烯类途径中所有已知基因的成功搜索,这些基因导致目标木脂素及其他重要化合物(如黄酮、木质素等)的合成,表明获得的组装具有满意的物种转录组覆盖度。另一方面,对于非CyP450s,克隆并确认组装的绝对准确性以及相应基因的实际功能并未进行,因为这超出了当前研究的范围。
尽管从初步评估的组装数据集中得到了非常令人鼓舞的结果,但仍需考虑这一整体方法的潜在局限性。这是因为组装协议是在组装严格性和获得的转录本长度之间的权衡。因此,总是有可能发现嵌合contig,尤其是在高度相似的转录本(近亲同源基因)或包含重复元素的情况下。另一方面,如果增加组装程序的严格性以最小化嵌合contig的存在,结果可能会增加不完全和冗余的contig数量(即同一转录本对应多个contig)。因此,在组装过程中,优化了多个参数以达到平衡,但在最终组装中仍然可以观察到这两种情况。
CyP450s在鬼臼毒素生物合成中的作用
一旦评估并验证了组装数据的质量,便开始了对可能参与(−)-鬼臼毒素(1b)生物合成最后步骤的未知推测基因的搜索。转换(−)-罗汉松脂素(9b)为(−)-鬼臼毒素(1b)所需的生化修饰可以容易推断(见图2),但这些反应的顺序则存在几种可能的排列方式。实际上,这些反应可能并行发生在一个生物合成“网格”中,或者通过一系列特定的转换反应(见图2)。为了验证可能底物的作用,需要测试多种化合物,其中大部分是商业上无法获得的。然而,从(−)-罗汉松脂素(9b)开始,预计生物合成途径将涉及多个CyP450,包括两次羟基化、一次碳-碳(C-C)耦合/环化和甲基二氧桥形成(见图2)。此外,尽管C-C耦合和7位的羟基化可能由非CyP450酶进行(例如,在第一种情况下可能由漆酶进行,/或者在后者情况下由依赖α-酮戊二酸的双加氧酶进行),但甲基二氧桥功能的形成应由CyP450酶催化。因此,首先选择用于研究的基因是CyP450s。
CyP450s是一个非常庞大(例如拟南芥中有超过200个基因)且多样的家族,在植物次级代谢中具有关键作用(49–51)。其中许多参与苯丙烯类代谢,并已从多个植物物种中被识别、克隆和表征。更具体地说,它们可用于单木酚类的生物合成,其中香草酸-4-羟化酶、对香豆酰CoA 3-羟化酶和阿魏酸5-羟化酶负责C6C3核心的连续羟基化(52, 53),以及在许多下游产物的生物合成途径中起作用,如黄酮(54)和木脂素(41, 55)的合成。
然而,预测CyP450生理功能通常是一个困难的任务。这是因为不同基因家族的成员可以执行相似的功能,在某些情况下,同一家族的成员可以催化不同的反应(51)。为了增加获得可能负责甲基二氧桥形成的CyP450基因的概率,使用了几种已知的CyP450作为模板,搜索可能参与(−)-鬼臼毒素(1b)及相关苯丙烯类生物合成的同源基因。最初,集中在桃儿七,按“实验方法”描述挖掘其转录组,寻找与香草酸-4-羟化酶(CYP73A1)、对香豆酰CoA 3-羟化酶(CYP98A)、阿魏酸5-羟化酶(CYP84A3)、黄酮类6-羟化酶(CYP71D9)、黄酮类3′-羟化酶(CYP75A1)、可可托贝林合酶(CYP80G2)、(+)-δ-可可托贝林8′-羟化酶(CYP706B1)、(S)-可可托贝林合酶(CYP719)和紫杉醇/紫杉醇合酶(CYP81Q)相似的基因。然后,选择组装转录组中同源性最高的contig进行克隆和进一步分析(附表S2)。所有克隆的基因与转录组contig序列完全匹配(100%),因此再次验证了所采用的测序和组装程序。
按照“实验方法”中描述的方式,每个上述克隆的候选基因都被异源表达,并对一系列可能的底物进行了酶活性测试。首先测试的基因编码已知酶的同源物CYP73A107(香草酸-4-羟化酶同源物)、CYP98A68(对香豆酰CoA 3-羟化酶同源物)、CYP84A52(阿魏酸5-羟化酶同源物)和CYP71BE30(黄酮类6-羟化酶同源物);这些CyP450s经过测试,发现它们能够执行预期的酶促转化(数据未显示)。CYP73A107表现出最高的活性,因此在所有测试中作为阳性对照。接下来,我们开始研究(−)-鬼臼毒素(1b)和相关木脂素的甲基二氧桥形成步骤。
鬼臼毒素生物合成中的甲基二氧桥形成
CyP450催化的甲基二氧桥形成已在异黄酮(56)、木脂素(41, 55)和生物碱(42, 57–59)生物合成中有所描述,并且已克隆并表征了编码基因。对转录组数据进行了查询,寻找与CyP450家族相似的序列,并成功识别了与木脂素(CYP81Q1和CYP81Q2)(41)和生物碱(CYP719A1和AY610513)(42, 57)生物合成相关的甲基二氧桥形成酶同源物。在组装的桃儿七转录组中,发现一个完整的转录本,其与前者有约50%的同一性,编码为CYP81B57(附表S2),另一个完整的候选基因与后者的同一性约为68%,编码为CYP719A23(见图5)。随后,北美桃儿七的组装转录组也进行了查询,选择了转录本CYP719A24,显示与CYP719A1的同一性约为68%,与CYP719A23的同一性约为96%(见图5)。

甲基二氧桥形成细胞色素P450的序列比对 从桃儿七和北美桃儿七克隆的CYP719A23和CYP719A24分别显示与Coptis japonica的(S)-可可托贝林合酶(CYP719A1)具有约68%的同一性。
这三种候选基因被克隆,并通过传统的Sanger测序(60)确认其序列。随后,每个基因单独在酵母中表达,得到的重组蛋白用于检测甲基二氧桥的形成,使用了一系列可能的底物及其类似物,包括(−)-罗汉松脂素(9b),这是在鬼臼属植物中确认的(−)-鬼臼毒素(1b)生物合成的最后中间体,(±)-7′-羟基罗汉松脂素(10a/b),(±)-7-羟基罗汉松脂素(32a/b),(−)-5-甲氧基罗汉松脂素(35b),(±)-异牛蒡子苷元(33a/b),(−)-牛蒡子苷元(34b),(±)-松脂素(6a/b),(±)-薄荷醇(37a/b),(+)-连翘脂素(38a)和(−)-α-铁杉脂素(36b)(图3)。其中,(−)-7′-羟基罗汉松脂素(10b)已被证明是Linum flavum中(−)-5-甲氧基鬼臼毒素(39b)形成的中间体(18),而异牛蒡子苷元(33)如果(−)-罗汉松脂素(9b)在进行其他修饰之前首先在4′位进行甲基化,也可能是(−)-鬼臼毒素(1b)生物合成的中间体。其他类似物(±)-松脂素(6a/b)、(−)-牛蒡子苷元(34b)、(−)-α-铁杉脂素(36b)、(±)-薄荷醇(37a/b)和(+)-连翘脂素(38a)具有类似的整体结构,可能是中间体,因此进行了测试以评估酶的底物多样性。
首先评估的候选基因是紫杉醇/紫杉醇合酶同源物,即CYP81B57。在我们的实验中,未观察到催化活性,包括对紫杉醇/紫杉醇合酶的天然底物(+)-松脂素(6a)和(+)-薄荷醇(37a)的活性(数据未显示)。
接下来,我们评估了与生物碱生物合成相关的酶同源物,假设这些酶参与了甲基二氧桥的形成。这些(S)-可可托贝林合酶同源物在NADPH存在的条件下对(−)-罗汉松脂素(9b)进行了催化,形成了一个在超高效液相色谱分析中具有较长保留时间的产物(图6A),其质子化分子离子在m/z 357(图6C)处,较(−)-罗汉松脂素(9b)丢失了两个氢原子(图6B)。然而,对于其他测试的假定底物,并未检测到任何活性和/或产物形成(数据未显示),表明(S)-可可托贝林合酶同源物对(−)-罗汉松脂素(9b)具有较强的底物特异性。从通过甲基二氧桥形成酶形成的三个可能产物(每个芳香环中的一个或两个甲基二氧桥)中,预计的生物合成途径反应产物是(−)-松脂酚(14b),而不是(−)-haplomyrfolin(40b)。在这方面,酶产物的裂解模式与文献中描述的(−)-松脂酚(14b)一致(61, 62)(图7)。产物在负离子模式下的基峰为m/z 355.1183, correspond to [M − H]− (计算值355.1182)(数据未显示)。正离子模式的质谱更加信息丰富(图6C),显示基峰为m/z 339.1230,对应于[M + H − H2O]+(计算值339.1232),以及m/z 357.1337和m/z 379.1155,分别对应于[M + H]+(计算值357.1338)和[M + Na]+(计算值379.1157)。更重要的是,可以观察到预测的裂解图谱,明确指示了甲基二氧桥的形成;即,特征性替代的环戊烯阳离子对应甲基二氧桥基团,m/z 135.0445(计算值135.0446),以及带有羟基甲氧基取代的片段,m/z 137.0604(计算值137.0603)。最后,还观察到m/z 161.0605的碎片,这有助于区分两个潜在的同分异构产物(−)-松脂酚(14b)和(−)-haplomyrfolin(40b)(图7);这对应于烯丙基苯并二噁烯片段(计算值161.0603),并表明甲基二氧桥是在预期的环中形成的(−)-松脂酚(14b),而替代产物(−)-haplomyrfolin(40b)将产生m/z 163.0754的4-烯丙基-2-甲氧基苯酚片段(图7)。因此,获得的产物在正确的芳香环中引入了甲基二氧桥。对酶生成的产物的1H、13C和异核单量子相关核磁共振(NMR)光谱的分析也清楚地支持了产物的形成,符合文献中的数值(63),例如,甲基二氧桥特征性峰值分别出现在1H和13C NMR光谱中的5.9和101 ppm(附表S3和图S1–S3)。
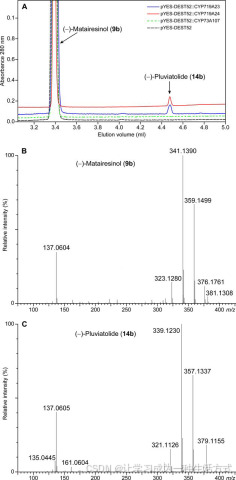
酶学分析的超高效液相色谱-质谱分析 A、超高效液相色谱色谱图显示了CYP719A24和CYP719A23酶反应中的产物形成,并与负对照(空载体和香草酸-4-羟化酶,CYP73A107)进行了比较。 B、底物(−)-罗汉松脂素(9b)和 C、产物(−)-松脂酚(14b)的正离子质谱显示了失去了两个质量单位。
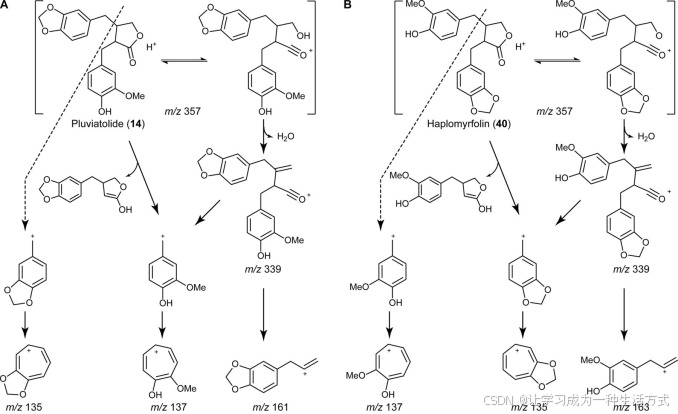
(−)-松脂酚(14b)和(−)-haplomyrfolin(40b)的裂解图谱 通过LC-ESI-MS分析生成的由这两种同分异构化合物14b(A)和40b(B)产生的预期碎片如图所示(改编自Schmidt et al.(61))。m/z 161处的碎片和m/z 163处碎片的缺失指示(−)-松脂酚(14b)是CYP719A23和CYP719A24的产物。
假定的松脂酚合成酶的动力学数据
使用(−)-罗汉松脂素(9b)作为底物,来自表达桃儿七 CYP719A23的酵母微体(其浓度为72 pmol,通过分析还原CO结合光谱确定,见图8B)显示出与米氏-门捷列夫动力学一致的饱和曲线,Km为9.7 ± 2.2 μm(见图9A),kcat为14.9 ± 1.0 min−1。此外,从北美桃儿七克隆的同源物CYP719A24,浓度为26 pmol(通过还原CO结合光谱确定,见图8C),显示出表观Km为5.8 ± 1.4 μm(见图9B)和kcat为7.8 ± 0.4 min−1。这种对(−)-罗汉松脂素(9b)的紧密结合表明,这些来自两个物种的CyP450可以暂时假定为参与甲基二氧桥形成,进而导致(−)-鬼臼毒素(1b)和相关木脂素的形成。然而,最终确认(−)-松脂酚(14b)是否为真正的生物合成中间体还需要通过体内验证,例如通过下调或敲除该基因并识别代谢物谱中的相应变化。然而,至今尚无用于转化鬼臼属植物的系统。
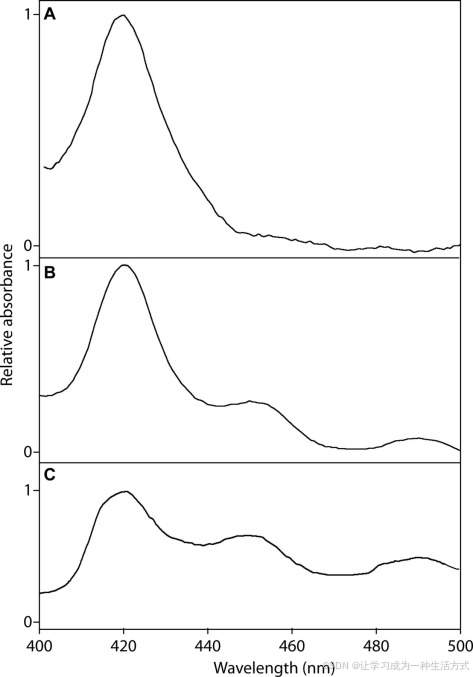
(−)-松脂酚合成酶的还原CO结合光谱 光谱数据通过从转化了pYES-DEST52(空载体作为阴性对照)(A)、pYES-DEST52::CYP719A23(B)和pYES-DEST52::CYP719A24(C)的大肠杆菌(株WAT11)中分离的微体获得。
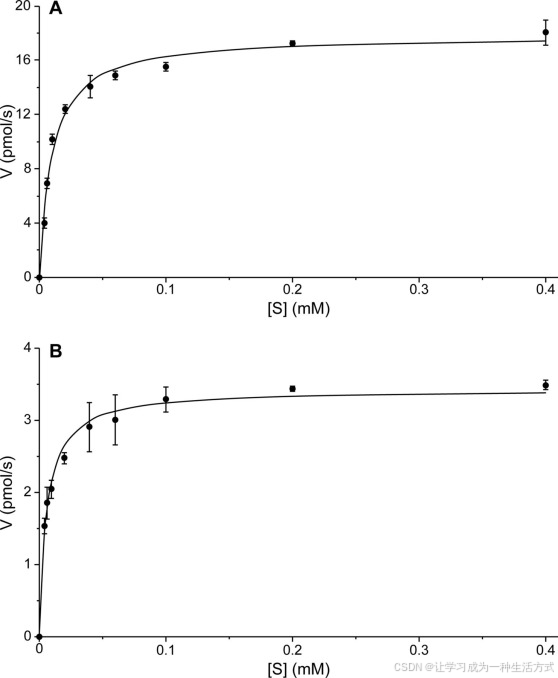
CYP719A23和CYP719A24的动力学参数 来自CYP719A23(A)和CYP719A24(B)的富集微体的稳态米氏-门捷列夫动力学数据,底物为(−)-罗汉松脂素(9b)。所有实验均进行了三次重复。
非常有趣的是,与生物碱生物合成酶CYP719A1(S)-可可托贝林合酶(图10)具有最高同源性的候选基因,发现它们能够在木脂素代谢中执行相同的反应,而与木脂素紫杉醇/紫杉醇合酶(来自胡麻科Sesamum indicum)密切相关的候选基因CYP81B57则未能检测到活性。然而,鬼臼属植物属于毛茛科(Berberidaceae)和毛茛目(Ranunculales),这是迄今为止唯一描述了CYP719基因的家族(64)。鬼臼属植物还与其他已知生物合成类似芳香四氢呋喃木脂素的物种(如Dysosma(66, 67)和Diphylleia(68)物种)形成了一个密切相关的系统发育群体,但到目前为止并未报告其中含有生物碱。这一分支与许多生产苯基异喹啉生物碱的物种(69)形成单系群体,包括其他已描述的CYP719基因的物种。因此,可以初步推测,鬼臼属植物群体可能没有或只有极少的生物碱合成水平,同时“招募”了其中的一些基因用于鬼臼毒素的生物合成途径。
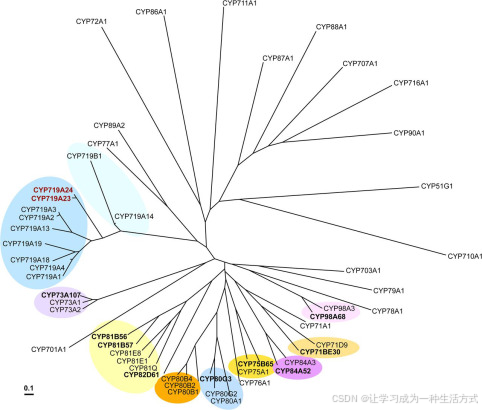
克隆(加粗)和已知细胞色素P450酶的系统发育分析 通过使用ClustalW(版本1.4)进行序列比对,并使用邻接法聚类算法(版本3.5c)构建的系统发育树,并通过Tree View(版本1.6.6)进行可视化。氨基酸序列来自UniProtKB、SwissProt或GenBankTM,相关的登录号如下:AB014459, CYP51G1, A. thaliana;AF212990, CYP701A1, Cucurbita maxima;AB006790, CYP703A1, Petunia × hybrida;NM_202845, CYP707A1, A. thaliana;M32885, CYP71A1, Persea americana;O81971, CYP71D9, Glycine max;NM_129002, CYP710A1, A. thaliana;NP_850074, CYP711A1, A. thaliana;NM_123002, CYP716A1, A. thaliana;Q948Y1, CYP719A1, Coptis japonica;EU882969, CYP719A2 和 AB126256, CYP719A3, Eschscholzia californica;EU883001, CYP719A4, Thalictrum flavum;EF451151, CYP719A13, Argemone mexicana;EF451152, CYP719A14, A. mexicana;AB374407, CYP719A18, C. japonica;AB374408, CYP719A19, C. japonica;EF451150, CYP719B1, P. somniferum;L10081, CYP72A1, Catharanthus roseus;Z17369, CYP73A1, Helianthus tuberosus;NP_180607, CYP73A2, A. thaliana;Z22545, CYP75A1, Petunia × hybrida;X71658, CYP76A1, Solanum melongena;X71656, CYP77A1, S. melongena;P48420, CYP78A1, Zea mays;U32624, CYP79A1, Sorghum bicolor;U09610, CYP80A1, Berberis stolonifera;AF014801, CYP80B1, E. californica;AB025030, CYP80B2, C. japonica;AY610509, CYP80B4, T. flavum;AB288053, CYP80G2, C. japonica;P93147, CYP81E1, Glycyrrhiza echinata;AY278229, CYP81E8, Medicago truncatula;BAE48234, CYP81Q, Sesamum indicum;NP_195345, CYP84A3, A. thaliana;P48422, CYP86A1, A. thaliana;AF216313, CYP87A1, H. annuus;U32579, CYP88A1, Z. mays;U61231, CYP89A2, A. thaliana;Q42569, CYP90A1, A. thaliana;NP_850337, CYP98A3, A. thaliana。比例尺表示10%的变化。
结论
本研究展示了大规模平行测序技术在非模式鬼臼属药用植物研究中的应用。基于Illumina的技术生成了大量高质量数据,与通过传统Sanger测序获得的克隆转录本的序列完全一致(100%)。除了验证通向(−)-鬼臼毒素(1b)的木脂素途径中已知序列的有效性外,我们研究的来自桃儿七和北美桃儿七的两种CyP450酶被认为是可能的松脂酚合成酶,即能够催化(−)-鬼臼毒素(1b)生物合成途径中的甲基二氧桥形成。作为本研究的延伸,并为科学界提供资源,我们所研究物种的转录组数据和代谢谱数据,以及其他几种重要药用植物的数据,已在开放网站MedPlTranscriptome和Medplants中提供,供科研人员使用。

















 3357
3357

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









