






母乳中的补体可以改变后代肠道微生物群,促进婴儿健康
Complement in breast milk modifies offspring gut microbiota to promote infant health

Article,2024-1-18,Cell, [IF 64.5]
DOI:https://doi.org/10.1016/j.cell.2023.12.019
原文链接:https://www.cell.com/cell/fulltext/S0092-8674(23)01384-3#%20
通讯作者:Fengyi Wan
- 重点 -
乳汁缺乏补体的断奶小鼠易发生肠道感染
乳汁中的补体选择性地消灭婴儿肠道中的某些革兰氏阳性微生物
母乳补体通过 C1 依赖、抗体独立途径激活
生命早期肠道微生物群调节新生儿对肠道感染的易感性
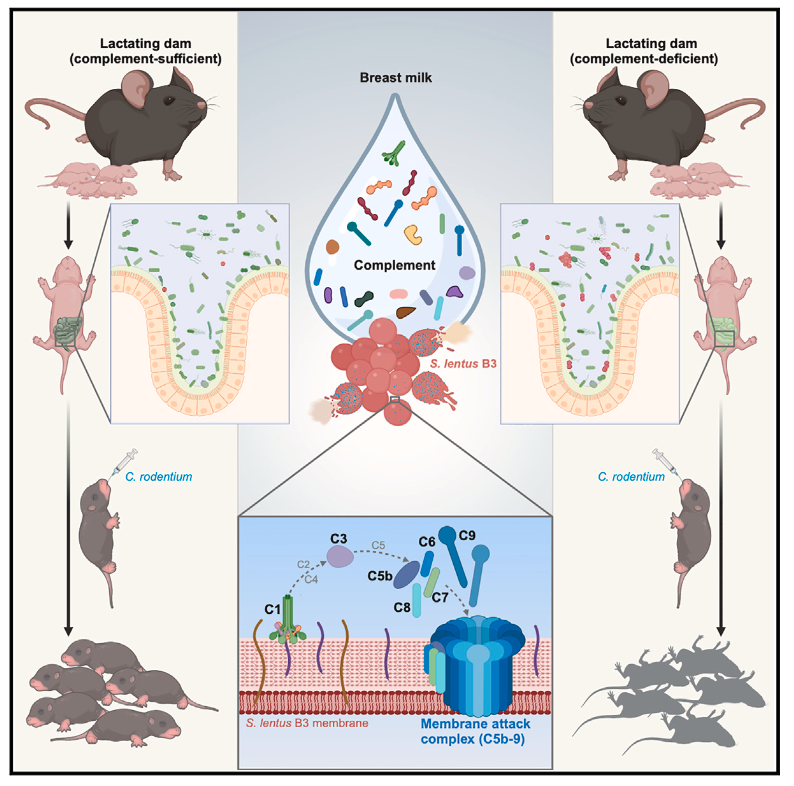
图形概要
- 摘要 -
母乳喂养通过提供营养和免疫保护以及塑造肠道共生微生物群,为新生儿和婴儿带来明显的好处。尽管几十年来人们一直认识到母乳中含有补体成分,但母乳中补体的生理相关性仍然不明确。在这里,我们证明,由补体缺乏的母鼠饲养的断奶小鼠在暴露于鼠类病原体啮齿枸橼酸杆菌(Citrobacter rodentium,CR)时会迅速死亡,而母乳中含有补体的乳补体成分饲养的幼鼠则被证明可以通过 C1 依赖性、抗体非依赖性机制直接裂解革兰氏阳性肠道共生微生物群的特定成员,导致膜攻击复合物的沉积和随后的细菌裂解。母乳中的补体成分通过选择性消除共生肠道群落的成员,塑造新生儿和婴儿肠道微生物组成,以抵御CR 等环境病原体。
- 结果 -
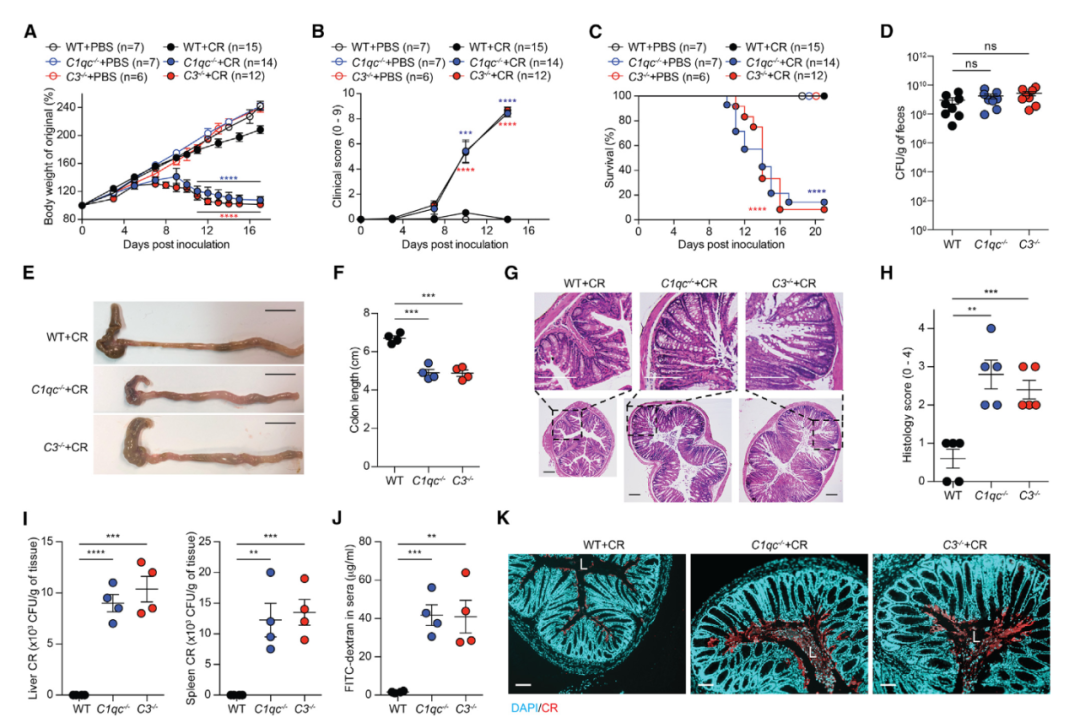
图1 补体缺乏的幼崽容易受到CR感染。
A-C. 2 x 109 CFUs的CR处理或PBS对照的21天大的野生型(WT)、C1qc-/-和C3-/-小鼠在特定接种后天数(dpi)的体重变化(A),临床打分(B)和存活(C)。
D. 7 dpi,从WT、C1qc-/-和C3-/-幼崽粪便样品中复苏的活CR。
E和F. 9 dpi时来自CR感染后的幼崽结肠代表性肉眼图(E)和长度(F)。
G和H. 9 dpi时来自CR攻击的幼崽的结肠切片的苏木精和伊红染色(G)和组织病理学评分(H)。
I. 9 dpi时来自CR攻击的幼崽肝脏(左)和脾脏(右)中的CR负荷。
J. 口服FITC右旋糖酐后4小时CR感染的幼崽(9 dpi)血清中的FITC右旋糖酐浓度。
K. 在9 dpi时来源于CR感染的幼崽的结肠中CR的代表性免疫荧光显微照片,细胞核用DAPI复染。L、 结肠腔。
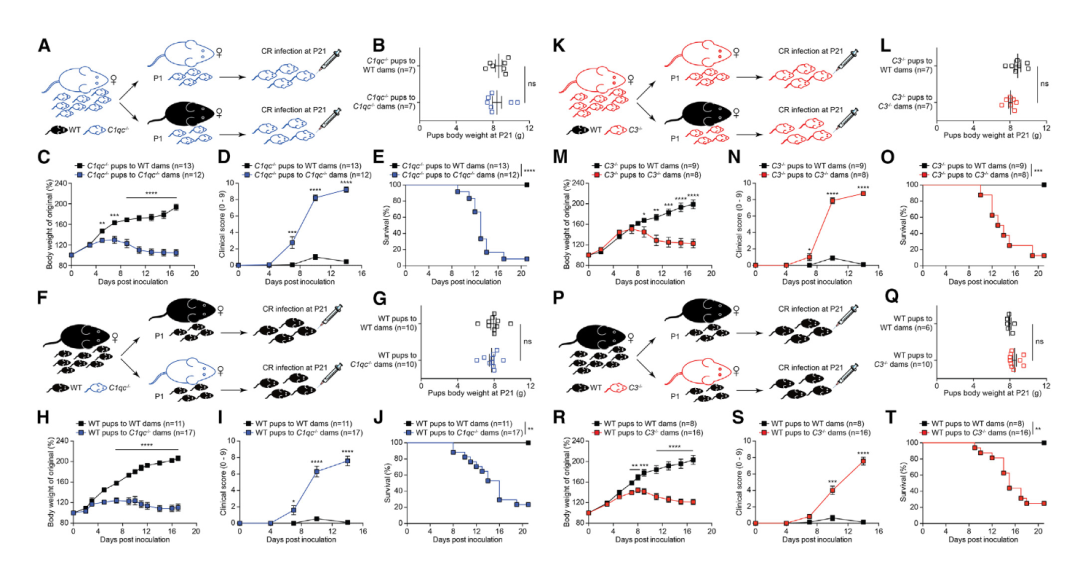
图2 母乳中的补体保护断奶小鼠免受CR感染引起的生长迟缓和致死。
A和F. 交叉喂养策略实验方案。
B和G. 出生21天后,CR感染之前,交叉喂养的C1qc-/-(B)和WT(G)幼崽的体重。
C-E. 交叉喂养的C1qc-/-幼崽在特定CR接种后天数(dpi)的体重变化(C)、临床打分(D)和存活(E)。
H-J. 交叉喂养的WT幼崽在特定dpi的体重变化(H)、临床打分(I)和存活(J)。
K和P. 交叉喂养策略实验方案。
L和Q. 出生21天后,CR感染之前,交叉喂养的C3-/-(L)和WT(Q)幼崽的体重。
M-O. 交叉喂养的C3-/-幼崽在特定dpi的体重变化(M)、临床打分(N)和存活(O)。
R-T. 交叉喂养的WT幼崽在特定dpi的体重变化(R)、临床打分(S)和存活(T)。
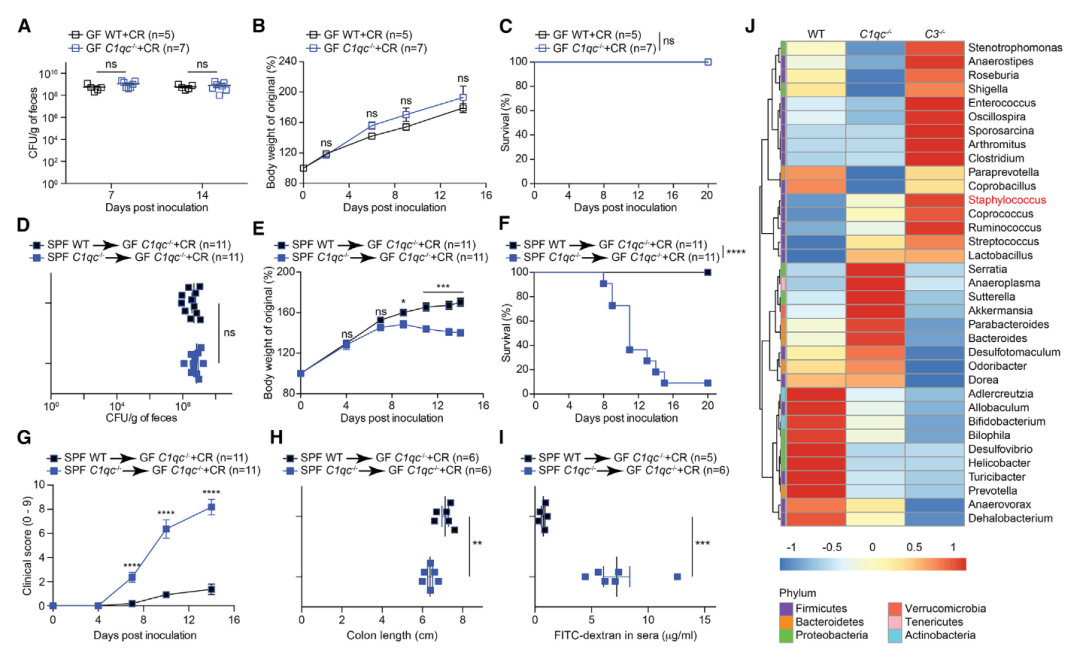
图3 乳鼠肠道微生物群在CR感染致死中起着关键作用。
A. 在7 dpi和14 dpi,从2 x 109 CFUs CR处理的21天大的无菌WT和C1qc-/-小鼠粪便样品中复苏的活CR。
B和C. 2 x 109 CFUs CR处理的21天大的无菌WT和C1qc-/-幼崽在特定dpi的体重变化(B)和存活(C)。
D. 用2 x 109 CFUs CR处理的,7 dpi的21天大的无菌C1qc-/-幼崽(在出生后第17天用来源于出生后21天的无特异性病原体(SPF)WT或C1qc-/-幼崽的盲肠和结肠微生物群重建)的粪便样本中复苏的活CR。
E-G. 在特定dpi,如D中重建并感染的21天大的无菌C1qc-/-幼崽的体重变化(E)、存活(F)和临床打分(G)。
H. 12 dpi,如D中重建并感染的无菌C1qc-/-幼崽的结肠长度。
I. 无菌C1qc-/-幼崽如D中重建并感染。口服FITC右旋糖酐4 h后无菌C1qc-/-幼崽(12 dpi)血清中FITC右旋糖酐浓度。
J. 21天大的WT(n = 5)、C1qc-/-(n = 4)和C3-/-(n = 3)幼崽盲肠和结肠内容物中,用16S rRNA基因为基础的高通量测序检测到的主要的35个属的物种丰度热图。
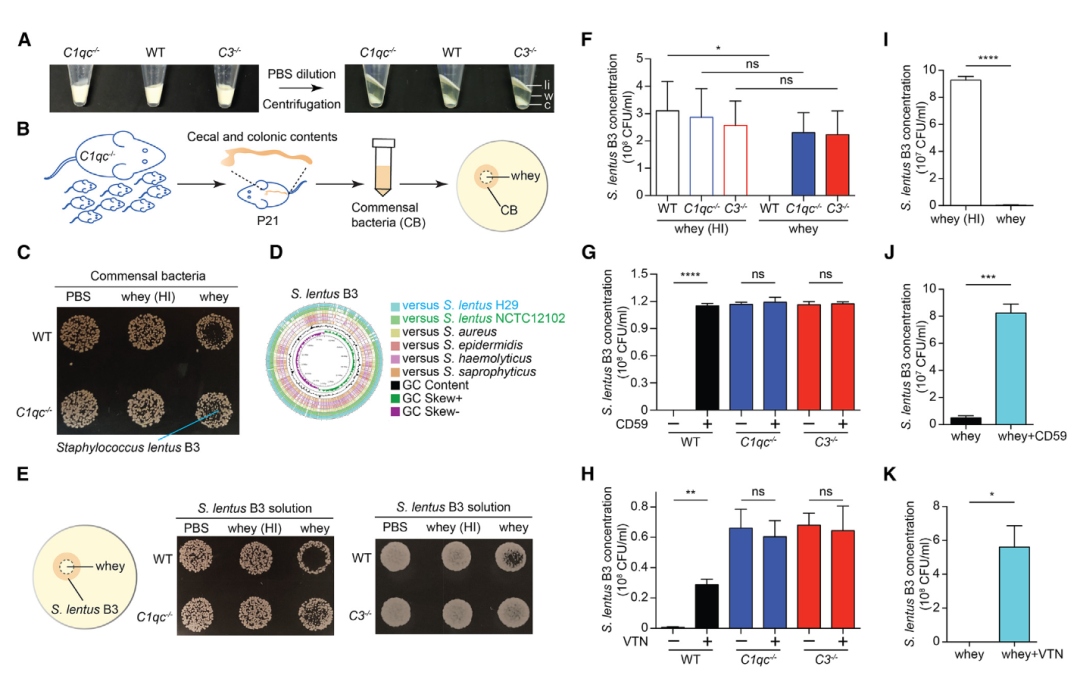
图4 母乳补体杀死革兰氏阳性共生菌慢速葡萄球菌B3。
A. 源自野生型(WT)、C1qc-/-和C3-/-母鼠的全脂牛奶和乳清(经过PBS稀释和离心)的代表性肉眼图。li:脂质;W:乳清;C:酪蛋白。
B. 使用源自出生后第21天C1qc-/-幼崽的可培养共生菌(CB)进行小鼠乳清杀菌试验的实验方案。
C. 使用WT和C1qc-/-小鼠乳清杀死来源于C1qc-/-幼崽的CB,PBS和热灭活(HI)乳清作为阴性对照的LB琼脂平板上杀菌试验的代表性肉眼图。指示的是从C1qc-/-幼崽的可培养CB中分离出的一种,慢速葡萄球菌B3菌株。
D. 慢速葡萄球菌B3与其他代表性葡萄球菌基因组的圆形比较基因组图谱。
E. 使用所示的小鼠乳清杀死慢速葡萄球菌B3,PBS和乳清(HI)作为阴性对照的LB琼脂平板上杀菌试验的代表性肉眼图。左边:实验方案。
F-H. 使用在 LB 培养基中培养的慢速葡萄球菌进行乳清杀菌测定,补充来自WT、C1qc-/-和C3-/-母鼠以及HI对照(F)的乳清,或在存在CD59(G)或玻连蛋白(VTN)的情况下进行(H)。
I-K. 使用在 LB 培养基中培养的慢速葡萄球菌B3进行人乳清杀菌测定,补充常规或热灭活(HI)人乳清(I),或在存在CD59(J)或VTN (K)的情况下进行。
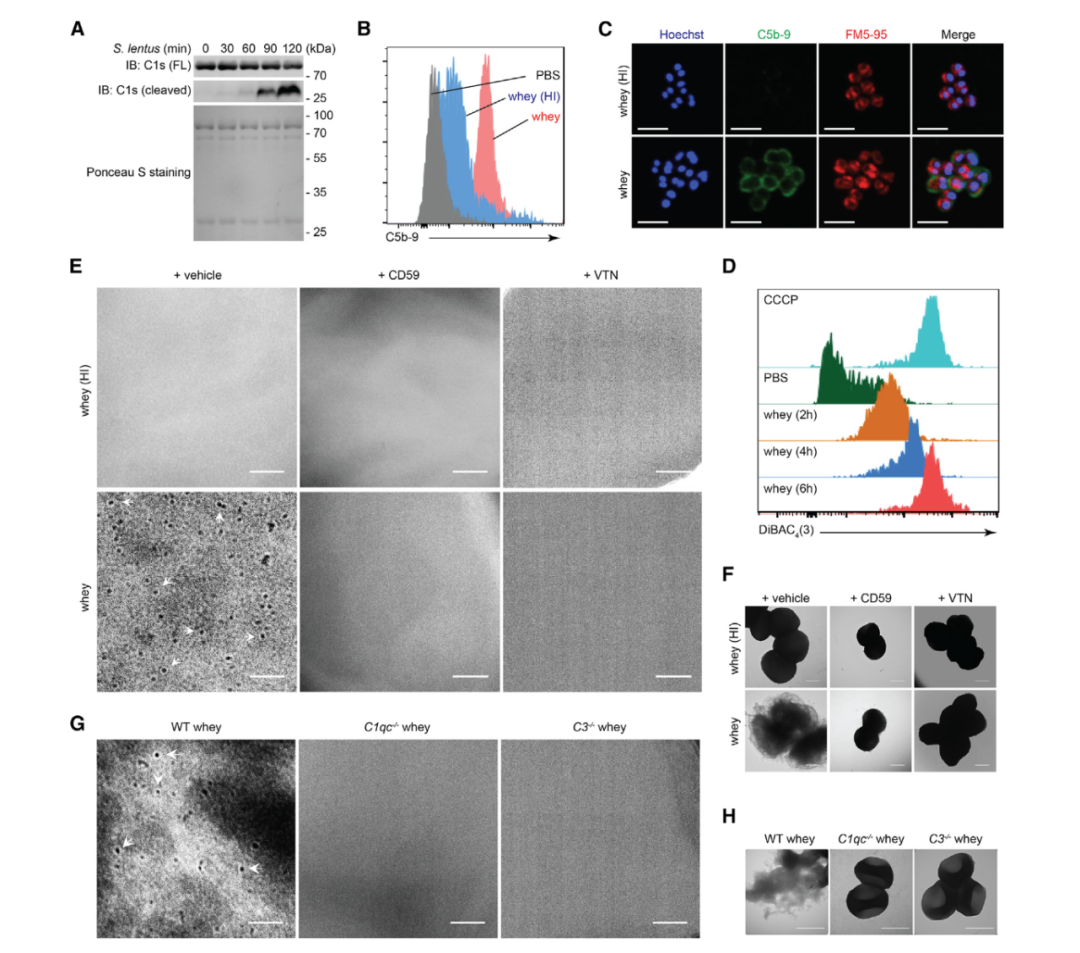
图5 母乳中的补体通过C1激活和MAC沉积裂解慢速葡萄球菌B3。
A. 将人乳清和慢速葡萄球菌B3孵育指定的时间段,然后进行SDS-PAGE分离。
B. 与PBS、常规或热灭活(HI)人乳清孵育后,通过流式细胞术分析慢速葡萄球菌B3上的C5b-9水平。
C. 与指定的人乳清孵育后,分别用Hoechst和FM5-95对DNA和膜进行复染,在慢速葡萄球菌B3上C5b-9的代表性免疫荧光显微照片。
D. 与PBS或人乳清一起孵育指定时间段的通过流式细胞术进行分析慢速葡萄球菌B3上的DiBAC4(3)荧光的代表性直方图,质子载体CCCP作为阳性对照。
E和F. 在存在和不存在CD59或玻连蛋白(VTN)的情况下,与指示的人乳清一起孵育后,慢速葡萄球菌B3的代表性细胞膜(E)和细菌形态(F)的显微照片。白色箭头表示组装的环状结构的膜攻击复合体(MAC)孔。
G和H. 与来源于野生型(WT)、C1qc-/-和C3-/-母鼠的乳清孵育后,慢速葡萄球菌B3的代表性细胞膜(G)和细菌形态(H)的显微照片。白色箭头表示组装的环状结构的膜攻击复合体(MAC)孔。
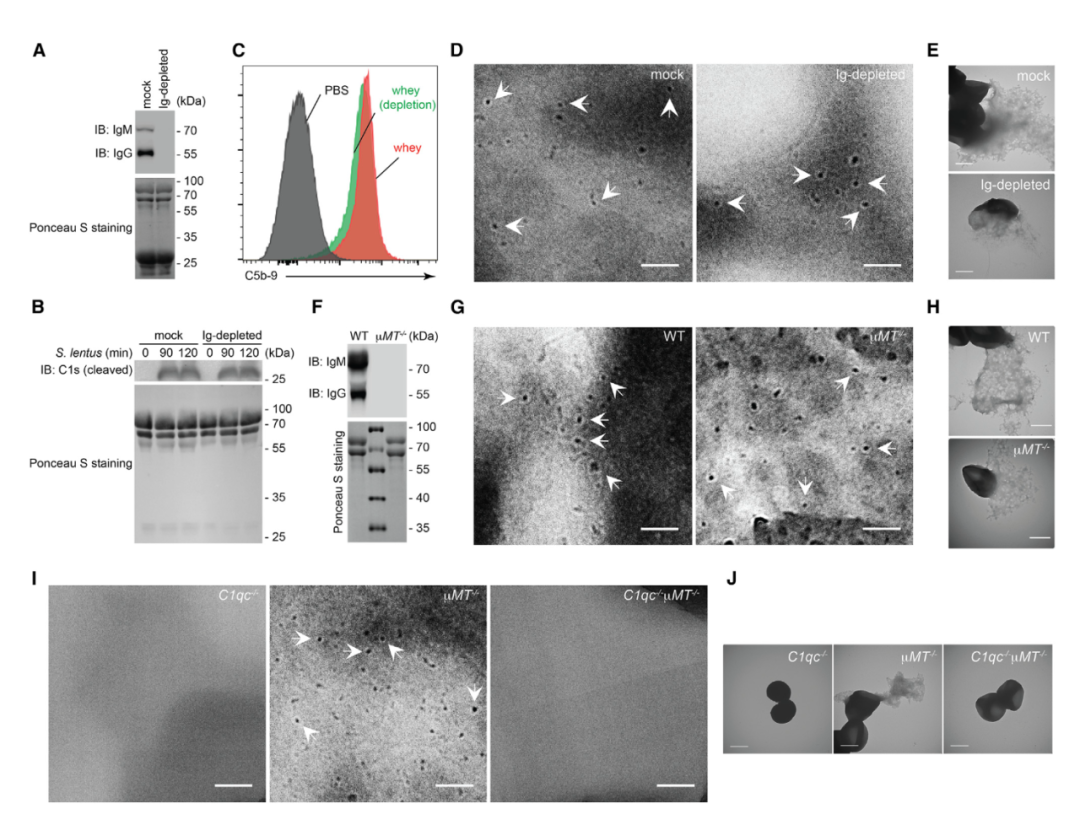
图6 母乳中的补体以不依赖抗体的方式杀死慢速葡萄球菌B3。
A. 将免疫球蛋白G(IgG)和IgM耗尽(Ig耗尽)和模拟处理的人乳清样品进行SDS-PAGE分离,然后进行IgM和IgG的Ponceau S染色或免疫印迹(IB)。
B. 将模拟处理的和Ig耗尽的人乳清与慢速葡萄球菌B3孵育指定的时间段,进行SDS-PAGE分离,然后对活化的C1s蛋白进行Ponceau S染色或IB。
C. 用PBS或指定的人乳清孵育后,通过流式细胞术分析慢速葡萄球菌B3上的C5b-9水平。
D和E. 与特定的人乳清孵育后的慢速葡萄球菌B3的代表性细胞膜(D)和细菌形态的显微照片(E)。
F. 将来源于野生型(WT)C57BL/6J和µMT-/-母鼠的乳清进行SDS-PAGE分离,然后进行IgM和IgG的Ponceau S染色或IB。
G-J. 与来源于特定母鼠的乳清孵育后,慢速葡萄球菌B3的代表性细胞膜(G)和(I)以及细菌形态(H)和(J)的显微照片。
参考文献
Xu D, Zhou S, Liu Y, Scott AL, Yang J, Wan F. Complement in breast milk modifies offspring gut microbiota to promote infant health. Cell, 2024.
- 通讯作者简介 -

约翰霍普金斯大学
Fengyi Wan
教授
解析宿主细胞、微生物群落和病原体之间的互作,来理解结肠的健康和疾病。
研究兴趣:信号转导;病原体-宿主-微生物群相互作用;RNA结合蛋白;结肠炎;结肠肿瘤发生;自身免疫性疾病。
宏基因组推荐
猜你喜欢
iMeta高引文章 fastp 复杂热图 ggtree 绘图imageGP 网络iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读

















 1051
1051

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









