







宿主代谢平衡微生物对胆汁酸信号的调
● 期刊:Nature (IF:50.5)
● DOI:https://doi.org/10.1038/s41586-024-08379-9
● 原文链接: https://www.nature.com/articles/s41586-024-08379-9
● 第一作者:Tae Hyung Won, Mohammad Arifuzzaman, Christopher N. Parkhurst
● 通讯作者:David Artis(dartis@med.cornell.edu) & Frank C. Schroeder(schroeder@cornell.edu)
● 发表日期:2025-1-8
● 主要单位:
美国康奈尔大学、美国杜克大学、美国密歇根州立大学、韩国嘉泉大学
摘要abstract
源自肠道微生物群的代谢物,包括胆汁酸(BA),广泛调节脊椎动物的生理机能,涵盖发育、新陈代谢、免疫反应以及认知功能。然而,我们仍不清楚宿主反应可以在多大程度上平衡微生物群衍生代谢物带来的生理影响。在此,通过对小鼠组织进行非靶向代谢组学分析,我们鉴定出一类胆汁酸-甲基半胱胺(BA-MCY)结合物,它们在肠道中含量丰富,且依赖于vanin 1(VNN1),这是一种在肠道组织中高度表达的泛酸酶。这种宿主依赖性的MCY结合作用逆转了胆汁酸在肝胆系统中的功能。微生物群衍生的游离胆汁酸作为法尼醇X受体(FXR)的激动剂,负向调节胆汁酸的生成,而BA-MCYs则作为FXR的强效拮抗剂,在体内促进胆汁酸生物合成基因的表达。补充稳定同位素标记的BA-MCY以FXR依赖的方式增加了胆汁酸的生成,并且在高胆固醇血症小鼠模型中补充BA-MCY减少了肝脏中的脂质积累,这与BA-MCYs作为肠道FXR拮抗剂的作用一致。在微生物群缺失的小鼠中,BA-MCY的水平降低,而通过移植人类粪便微生物群可使其恢复。用菊粉纤维进行饮食干预进一步提高了游离胆汁酸和BA-MCY的水平,这表明宿主产生BA-MCY受到微生物群衍生的游离胆汁酸水平的调节。我们进一步表明,多种BA-MCYs也存在于人类血清中。综上所述,我们的结果表明,宿主的BA-MCY结合作用平衡了依赖宿主和微生物群的代谢途径,而这些途径调节着FXR依赖的生理过程。
结果results
比较代谢组学揭示了胆汁酸-甲基半胱胺结合物(BA-MCYs)
在无菌小鼠中,由于缺乏微生物的去结合作用,导致胆汁酸-牛磺酸结合物水平升高,而游离胆汁酸的丰度则大幅降低。我们推测,尚未被注释的胆汁酸衍生物的丰度可能同样会因微生物群的缺失而受到影响。为了发现这类未被注释的化合物,我们首先通过基于高分辨率质谱(HRMS)的比较代谢组学方法,对拥有完整微生物群的无特定病原体(SPF)小鼠和无菌小鼠的样本进行分析,从而全面了解小鼠血清代谢组中依赖微生物群的变化情况(图1a)。所得数据集使用基于xcms的Metaboseek平台进行处理,该平台有助于识别在不同条件下丰度存在显著差异的质谱特征(图1b)。这种非靶向比较揭示了无菌小鼠和SPF小鼠之间的明显差异。通过对血清样本在正离子和负离子电离模式下的联合分析,我们总共检测到超过40,000个质谱特征,其中约10% 在无菌小鼠和SPF小鼠之间存在显著差异(P ≤ 0.05)。为了从大量差异特征中筛选出重点,我们聚焦于在所有重复样本中均能稳定检测到(见方法部分)且在无菌小鼠样本中相对于SPF样本差异至少达5倍的化合物。利用这些严格标准,我们在血清代谢组中检测到数百种依赖微生物群的代谢物(补充表1)。为了进一步表征这些代谢物,我们获取了所有差异代谢物的串联质谱(MS2)数据。为了专注于胆汁酸及其衍生物,我们应用了一个宽松的分子式过滤器,要求MS2碎片中存在完整的24碳骨架,这表明可能存在甾体骨架(见方法部分)。MS2网络分析揭示了三个主要的依赖微生物群的代谢物簇,其碎片模式表明它们代表胆汁酸或胆汁酸衍生物(图1c及补充图1-3)。其中两个簇分别代表胆汁酸-牛磺酸结合物和游离胆汁酸,正如预期的那样,由于缺乏微生物的牛磺酸去结合作用,在无菌小鼠中,胆汁酸-牛磺酸结合物的丰度大幅增加,而游离胆汁酸的丰度则大幅降低。相比之下,第三个簇似乎代表了一类先前未被注释的胆汁酸衍生物,与游离胆汁酸类似,它们在无菌小鼠中的丰度降低。
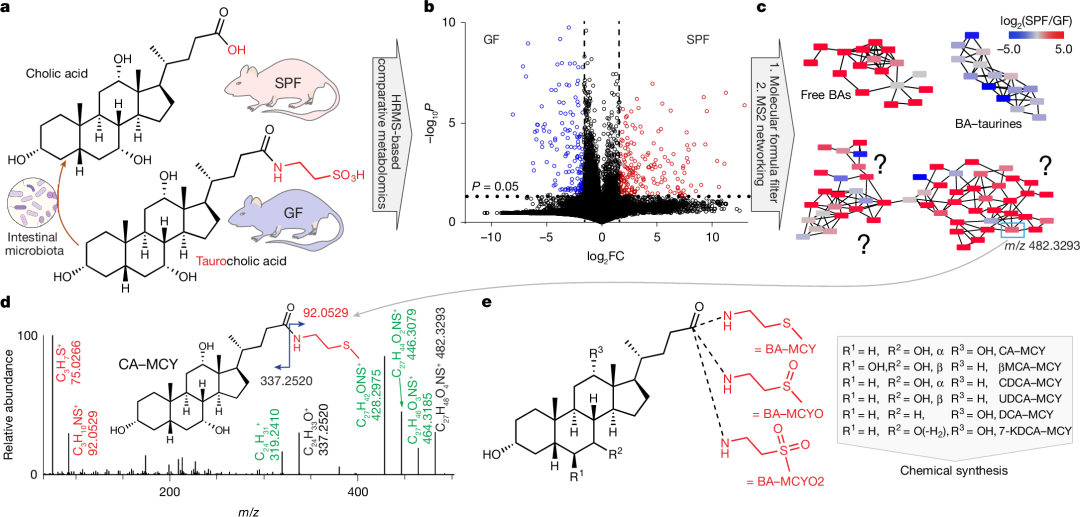
图1 | 胆汁酸甲基半胱胺结合物的鉴定
a,肠道微生物群在胆汁酸(BA)代谢中的主要作用,以及无菌(GF)小鼠和无特定病原体(SPF)小鼠比较分析策略的示意图。b,无菌(n=12)和SPF(n=9)小鼠血清中检测到的差异代谢物火山图。蓝色和红色圆点分别表示通过非配对双侧Student t检验计算得出,与SPF小鼠相比,在无菌小鼠中下调或上调5倍及以上(P<0.05)的代谢物。FC,倍数变化。c,以正离子电喷雾电离(ESI)和负离子电喷雾电离(ESI+-)模式获得的小鼠血清MS2网络(余弦值>0.7)的部分展示,显示代表游离胆汁酸、胆汁酸-牛磺酸结合物以及先前未注释的胆汁酸-甲基半胱胺(BA-MCY)结合物的簇。与SPF小鼠相比,在无菌小鼠血清中,红色节点表示下调,蓝色节点表示上调。完整的MS2网络(包括所有节点的质荷比(m/z)值)见补充图1-3。d,胆酸-甲基半胱胺(CA-MCY)的MS2谱图。红色的MS2碎片代表甲基半胱胺(MCY)基团;绿色的MS2碎片源于失水。e,在小鼠血清中鉴定出的胆汁酸-甲基半胱胺(BA-MCY)结合物的结构。图a中的示意图使用BioRender(https://biorender.com)绘制。
对推定的胆汁酸(BA)衍生物的质谱同位素模式和二级质谱(MS2)图谱进行详细分析后发现,其中存在甲基半胱胺(MCY)部分(图1d及扩展数据图1)。MS2图谱进一步表明,这些MCY衍生物可归为三个不同系列,分别代表推定的胆汁酸-甲基半胱胺结合物(BA-MCYs)、相应的亚砜(BA-MCYO)和砜(BA-MCYO2;扩展数据图1)。基于所分析样本中游离胆汁酸的相对丰度和保留时间,我们提出了这些BA-MCYs的结构,并通过合成标准品予以确认。最终共鉴定出18种胆汁酸衍生物,包括胆酸、β-鼠胆酸(βMCA)、鹅去氧胆酸(CDCA)、熊去氧胆酸(UDCA)、脱氧胆酸(DCA)和7-酮基脱氧胆酸(7-KDCA)的MCY、MCYO和MCYO2衍生物(图1e、补充表2及补充图4;合成步骤及核磁共振数据见补充信息)。综上所述,通过对无特定病原体(SPF)小鼠和无菌小鼠进行非靶向代谢组学比较,我们发现BA-MCYs是一类此前未被注释、依赖微生物群的胆汁酸衍生物。
胆汁酸-甲基半胱胺(BA-MCY)水平对微生物群的依赖性
与无特定病原体(SPF)小鼠的血清样本相比,无菌小鼠血清中胆汁酸-甲基半胱胺(BA-MCYs)的丰度显著降低,但并未完全消失(图2a、b,扩展数据图2a及补充表3)。同样,与SPF小鼠相比,无菌小鼠粪便中的BA-MCY水平也有所下降(扩展数据图2b-d)。为了更好地理解BA-MCY水平与微生物群存在之间的关系,接下来我们测试了将人类微生物群引入无菌小鼠体内是否会影响BA-MCY的产生。为此,我们将健康个体的粪便微生物群移植(FMT)到无菌小鼠体内,然后分析血清和粪便中的胆汁酸(BAs)及胆汁酸结合物。我们发现,接受人类粪便微生物群移植的小鼠,其血清和粪便中游离胆汁酸和BA-MCY结合物的丰度均大幅增加(图2a、b及扩展数据图2a-g)。
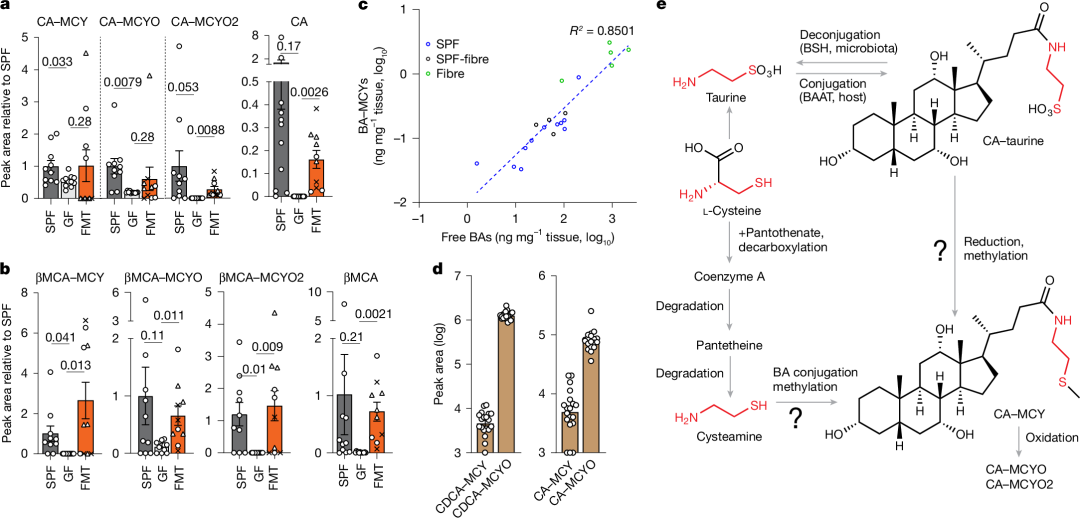
图2 | BA-MCY对微生物群的依赖性及其生物合成
a、b,无特定病原体(SPF)小鼠(n=11)、无菌小鼠(n=12)以及接受粪便微生物群移植(FMT)的小鼠(n=10;三位供体分别用三角形、圆形和十字表示,每位供体的样本数n=3-4)血清中胆-甲基半胱胺(CA-MCY)结合物(a)和β-鼠胆酸-甲基半胱胺(βMCA-MCY)结合物(b)的相对丰度,以及相应游离胆汁酸的相对丰度。数据以平均值±标准误表示。P值采用未配对双侧Student t检验并经Welch校正计算得出。
c,SPF小鼠(n=11)、菊粉纤维饮食研究中的SPF对照小鼠(n=4)以及菊粉纤维饮食喂养的SPF小鼠(n=5)肝脏中游离胆汁酸与胆汁酸-甲基半胱胺(BA-MCY)结合物丰度之间的关系。
d,人血清(n=19)中CA-MCY结合物或鹅去氧胆酸-甲基半胱胺(CDCA-MCY)结合物的相对丰度。数据以平均值±标准误表示。
e,胆汁酸-甲基半胱胺(BA-MCY)结合物来源的相关途径。MCY结合物可能源自肝脏酶BAAT产生的胆汁酸-牛磺酸结合物的还原,或者胆汁酸与来自辅酶A和泛酰巯基乙胺降解产生的半胱胺或其他半胱胺衍生物的结合。MCY结合物的氧化产生相应的MCYO和MCYO2结合物。
我们对无菌小鼠、无特定病原体(SPF)小鼠和接受人类粪便微生物群移植(FMT)小鼠进行比较的结果表明,胆汁酸-甲基半胱胺(BA-MCY)的水平在一定程度上受相应游离胆汁酸(BA)水平的调控。在先前的研究中,我们及其他研究人员已证实,补充菊粉纤维引起的微生物群变化可显著提高SPF小鼠血清中游离胆汁酸的水平。我们利用这一点来测试基于饮食干预导致的游离胆汁酸水平升高是否也会影响BA-MCY的水平。我们观察到,喂食菊粉基高纤维饮食的SPF小鼠中BA-MCY水平大幅升高(扩展数据图3a-c),这表明游离胆汁酸库的扩充会促使BA与MCY的结合增加。此外,我们发现喂食不同饮食的SPF小鼠中游离胆汁酸和BA-MCY结合物的水平通常具有相关性(图2c和扩展数据图3d、e)。最后,为了确定BA-MCYs是否也存在于人体中,我们分析了人类血清样本,结果显示人体中常见的所有主要胆汁酸均存在MCY衍生物(图2d和扩展数据图3f、g)。总之,这些数据表明,BA-MCY结合物存在于小鼠和人体中,并且在将人类粪便微生物群移植到无菌小鼠体内或因补充膳食纤维而导致游离胆汁酸水平升高时,相应的BA-MCY结合物也会平行增加。
BA-MCY的生化来源
为了进一步阐明宿主和微生物群在胆汁酸-甲基半胱胺(BA-MCY)结合物生成过程中的作用,我们接下来研究了体内半胱胺部分的来源。半胱胺主要通过辅酶A(CoA)的降解产生,半胱胺在肝脏及其他组织中氧化生成牛磺酸,随后牛磺酸在肝脏中与胆汁酸结合,生成胆汁酸-牛磺酸结合物,并分泌至肠道。因此,我们考虑了BA-MCYs中MCY部分来源的两种不同模型(图2e)。其一,这些化合物可能源于胆汁酸与半胱胺或由CoA分解产生的半胱胺衍生物的结合。其二,BA-MCY结合物可能源于宿主或肠道微生物群对相应胆汁酸-牛磺酸衍生物的还原(图2e)。为了区分这两种情况,我们在无特定病原体(SPF)小鼠中使用氘代牛磺酸(taurine-d4)和氘代L-半胱氨酸(L-cysteine-3,3-d2)进行了一系列稳定同位素标记实验(扩展数据图4a-c)。对补充了taurine-d4的SPF小鼠血清进行高效液相色谱(HPLC)-高分辨率质谱(HRMS)分析,如预期所示,发现胆汁酸的牛磺酸结合物广泛标记(扩展数据图4a)。然而,MCY结合物均未被标记(扩展数据图4a),这表明牛磺酸结合物和MCY结合物的生物合成途径不同。接下来,为了测试MCY结合物是否源于半胱氨酸衍生的半胱胺部分的掺入,我们分析了补充了L-cysteine-3,3-d2的SPF小鼠的血清样本(扩展数据图4b、c)。HPLC - HRMS分析显示,氘掺入了胆汁酸-牛磺酸结合物和胆汁酸-MCY结合物中(扩展数据图4b、c),这与牛磺酸和MCY部分均来源于半胱氨酸的情况相符。如预期一样,我们还观察到氘掺入了CoA的分解产物泛酰巯基乙胺中(扩展数据图4d)。
这些结果支持这样一种模型:胆汁酸-甲基半胱胺(BA-MCY)结合物是由除牛磺酸以外的一种半胱胺衍生物的酰化作用产生的。接下来,我们考虑了这种结合是可能由微生物群还是由宿主介导的。尽管BA-MCY结合物的丰度强烈依赖于微生物群,但在无菌动物中它们的产生并未完全消失(图2a、b和扩展数据图2a-d),这表明结合反应本身并不需要微生物群。因此,我们接下来研究了BA-MCY的产生是否与胆汁酸-牛磺酸结合物的生物合成有关。在肝脏中,胆汁酸与牛磺酸和氨基酸的结合是由BAAT(一种I型酰基辅酶A硫酯酶(ACOT))介导的。该基因家族的成员能催化广泛的酰基转移反应;据最近报道,在Baat−/−小鼠中胆汁酸与半胱胺和甲基半胱胺的推定结合物会积累。我们证实了在Baat−/−小鼠中积累的胆汁酸半胱胺衍生物与我们鉴定出的化合物相同(扩展数据图5a-d),这表明BA-MCY的生物合成与胆汁酸-牛磺酸结合是不同的。在Baat−/−小鼠中,由于缺乏牛磺酸结合,导致游离胆汁酸和BA-MCY结合物的水平大幅增加(扩展数据图5e、f),这与喂食高纤维饮食的小鼠中观察到的游离胆汁酸和BA-MCY结合物的平行增加情况一致。在Baat−/−小鼠中抑制微生物群并不会降低该突变株中升高的BA-MCY水平。总之,我们的结果表明,BA-MCYs源自一种与胆汁酸和牛磺酸结合并行存在的宿主依赖性结合途径。
接下来,我们考虑了胆汁酸-甲基半胱胺(BA-MCY)生物合成的潜在位点。虽然胆汁酸-牛磺酸结合物是在肝脏中产生的,但BA-MCY水平主要受游离胆汁酸丰度的调控,而在无特定病原体(SPF)小鼠中,游离胆汁酸主要源于肠道微生物群对胆汁酸-牛磺酸结合物和胆汁酸-甘氨酸结合物的去结合作用,这一事实表明肠道可能参与了BA-MCY的产生。因此,我们另外分析了小肠和盲肠中的BA-MCY水平,结果显示BA-MCY结合物在肠道组织中的含量比在肝脏中丰富得多(图3a和扩展数据图6)。此外,BA-MCY结合物与其氧化产物BA-MCYO和BA-MCYO2的比例在肠道组织中高于在肝脏、血清和粪便中(图3a和补充表3),这表明BA-MCY结合物是在肠道中对游离胆汁酸重吸收后由游离胆汁酸生成的。肠道产生BA-MCYs依赖于游离胆汁酸重吸收的这一模型进一步与我们的观察结果相符,即在不同饮食和条件下,游离胆汁酸和BA-MCYs的丰度是相关的(扩展数据图3a-c)。
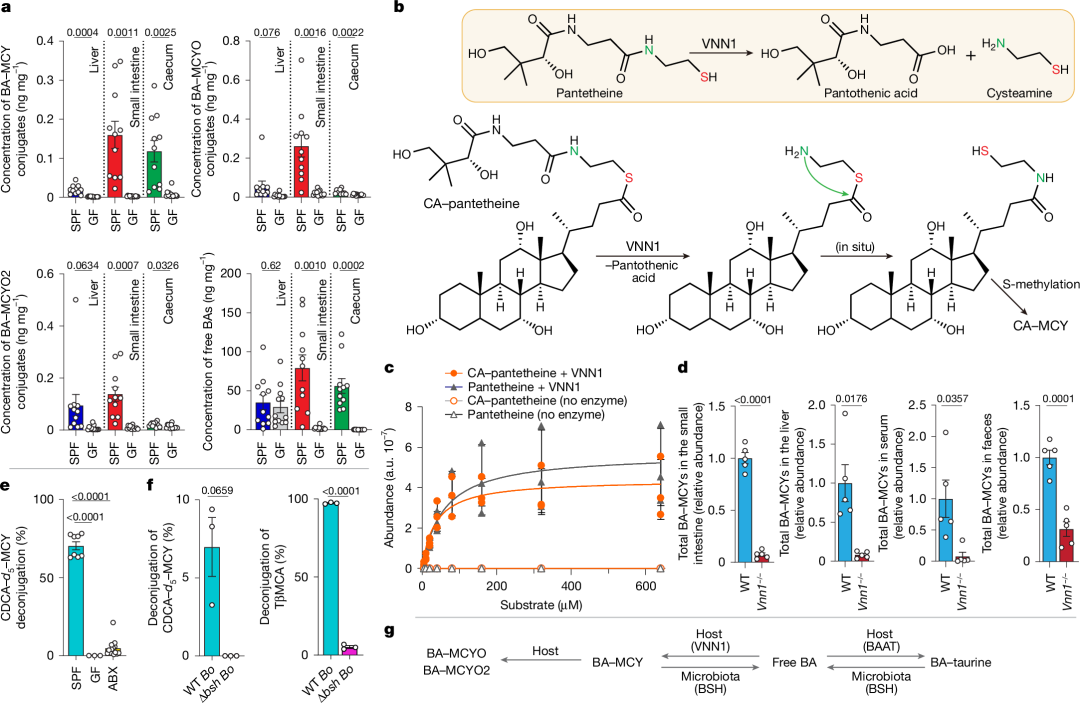
图3 | BA-MCYs的宿主依赖性生成与微生物去结合作用
a,无特定病原体(SPF)小鼠(n=11)以及无菌小鼠(肝脏样本n=12,小肠和盲肠样本n=13)肝脏、小肠和盲肠中胆汁酸-甲基半胱胺(BA-MCY)结合物及相应游离胆汁酸的总量。
b,VNN1在泛酰巯基乙胺水解中的既定功能(方框;上图),以及VNN1在胆酸-泛酰巯基乙胺(CA-pantetheine)水解中的推测作用,随后重排和甲基化形成CA-MCY(下图)。
c,在体外将一系列浓度的CA-泛酰巯基乙胺(CA-pant)和泛酰巯基乙胺与重组VNN1一起孵育,生成泛酸的情况。两种底物的反应均遵循饱和动力学。酶浓度为0.01μM。反应在37°C下孵育15分钟。使用同一批次酶进行的独立测定次数为三次。
d,野生型(WT,n=5)或Vnn1基因敲除(Vnn1−/−,n=5)小鼠小肠、肝脏、血清和粪便中BA-MCY结合物的相对丰度。
e,补充的氘代5-鹅去氧胆酸-甲基半胱胺(CDCA-d5-MCY)在SPF小鼠(n=8)、无菌小鼠(n=3)和使用抗生素处理(ABX,n=15)小鼠粪便中的去结合情况。
f,在分别定殖野生型(n=3)或敲除胆盐水解酶(BSH)基因的卵形拟杆菌(n=3;分别为WT Bo或Δbsh Bo)的无菌小鼠粪便中,胆汁酸的MCY或牛磺酸结合物的去结合情况。数据以平均值±标准误(a、d-f)或平均值±标准差(c)表示,P值采用未配对双侧Student t检验并经Welch校正计算得出。
g,宿主和微生物群在胆汁酸-牛磺酸及胆汁酸-甲基半胱胺结合物的生物合成和代谢中的作用。
BA-MCY的生物合成依赖于宿主的VNN1
由于BA-MCY结合物有可能源自泛酰巯基乙胺的分解,我们试图测试BA-MCY的生物合成是否可能通过在肠道组织中高表达的泛酰巯基乙胺酶VNN1进行,而肠道组织中BA-MCY的浓度是最高的。VNN1的主要功能是将泛酰巯基乙胺水解为半胱胺和泛酸(图3b),这是辅酶A循环利用的一部分,并且最近的研究表明VNN1在代谢调节、炎症及相关疾病中具有重要作用。尽管VNN1可能是BA-MCY生成所需半胱胺部分的一个合理来源,但我们假设,除了水解泛酰巯基乙胺外,VNN1可能也能够水解胆汁酸-泛酰巯基乙胺或胆汁酸-辅酶A结合物。由此产生的硫连接的胆汁酸-半胱胺衍生物会重排为氮连接的异构体,在进行硫甲基化后会生成BA-MCY结合物(图3b)。为了研究这一假设,我们首先测试了重组VNN1是否能够水解合成的胆酸-泛酰巯基乙胺结合物(CA-pant)。我们发现VNN1分解CA-pant的效率与分解泛酰巯基乙胺的效率相当(图3c和扩展数据图7a),并且进一步表明由此产生的CA-半胱胺结合物会重排为相应的胆酸-半胱酰胺(CA-CY),这是我们在Baat−/−小鼠中也检测到的CA-MCY的一种合理前体(扩展数据图7b)。为了评估VNN1是否在体内有助于BA-MCY的生物合成,我们比较了野生型(WT)和Vnn1−/−小鼠在不同组织中的胆汁酸谱(图3d)。我们发现Vnn1−/−突变小鼠的小肠、肝脏和血清中的BA-MCY水平显著降低,在粪便中的降低程度相对较小(图3d)。这些结果表明,体内BA-MCY的生物合成在很大程度上依赖于宿主酶VNN1。此外,我们发现CA-CY的预测前体CA-pant在Vnn1−/−小鼠的小肠和粪便中积累,而在相应的野生型样本中不存在这种化合物,在Vnn1−/−小鼠的肝脏和血清中也检测不到,这表明胆汁酸-泛酰巯基乙胺结合物是肠道中BA-MCY生物合成的直接前体(扩展数据图7c、d)。
BA-MCYs的微生物代谢与宿主代谢
牛磺酸和甘氨酸结合的胆汁酸在肠道中会被微生物胆盐水解酶(BSHs)高效地去结合从而生成游离胆汁酸。相应地,与无特定病原体(SPF)小鼠相比,无菌小鼠粪便中胆汁酸-牛磺酸结合物与游离胆汁酸的比例大幅增加(扩展数据图8a、b)。考虑到肠道微生物群可能也在胆汁酸-甲基半胱胺(BA-MCYs)的去结合过程中发挥作用,我们注意到,尽管无菌小鼠中BA-MCY的水平有所降低(扩展数据图2b-d和8c),但与SPF粪便样本相比,无菌小鼠中BA-MCY结合物与游离胆汁酸的比例大幅增加(扩展数据图8a)。事实上,在无菌小鼠粪便中,BA-MCY的水平与游离胆汁酸水平相近或超过游离胆汁酸水平(扩展数据图8d)。
为了确定BA-MCYs是否确实能被微生物群去结合,我们分析了补充了稳定同位素标记的鹅去氧胆酸-d5-甲基半胱胺(CDCA-d5-MCY)的SPF小鼠和无菌小鼠的粪便和肝脏样本,以检测是否存在游离的标记CDCA以及其他可由CDCA衍生的标记胆汁酸。为了广泛地研究BA-MCYs的代谢情况,我们还使用Metaboseek平台中的Label Finder方法通过非靶向代谢组学对补充了标记物的小鼠和对照小鼠进行了比较。对补充了CDCA-d5-MCY的SPF小鼠粪便样本的靶向分析显示,存在CDCA-d5-MCY以及d4标记和d5标记的游离CDCA(扩展数据图8e-g和补充图5a、b),这表明BA-MCYs可在肠道中被去结合。此外,我们检测到了其他可由CDCA衍生的标记游离胆汁酸(扩展数据图8f和补充图5b),而胆酸(CA)、脱氧胆酸(DCA)和其他由CA衍生的胆汁酸仍未被标记(补充图5b),这与它们各自独立的生物合成途径相符。对补充了CDCA-d5-MCY的SPF小鼠粪便和肝脏样本的进一步分析表明,由补充的CDCA-d5-MCY衍生的标记游离胆汁酸部分会与牛磺酸重新结合(扩展数据图8h-m和补充表4)。Label Finder分析还显示,未被去结合的补充CDCA-d5-MCY的其余部分会转化为其氧化衍生物,即CDCA-d4/5-MCYO,在较小程度上会转化为CDCA-d4/5-MCYO2。事实上,在补充了标记物的小鼠肝脏中只能检测到微量的CDCA-d5-MCY,这表明补充的CDCA-d5-MCY会迅速被氧化为CDCA-d4/5-MCYO(2)(扩展数据图8k、m)。
与SPF小鼠形成对比的是,在补充了鹅去氧胆酸-d5-甲基半胱胺(CDCA-d5-MCY)的无菌小鼠中未检测到去结合产生的标记CDCA或其他标记游离胆汁酸(图3e 和补充图5c、d),这表明CDCA-MCY的去结合依赖于微生物群。同样,与SPF小鼠相比,用抗生素(ABX)处理的SPF小鼠中微生物群受到抑制,导致补充的CDCA-d5-MCY的去结合显著减少(图3e和扩展数据图8f、g、j、k)。在ABX处理的小鼠和无菌小鼠中,补充的CDCA-d5-MCY 主要转化为相应的氧化产物,即CDCA-d4/5-MCYO和CDCA-d4/5-MCYO2(扩展数据图8g、k、m和补充表4)。
接下来,我们证明了BA-MCY结合物可被从SPF小鼠获取的粪便悬液以及已知具有编码胆盐水解酶(BSH)基因的单个肠道细菌去结合(扩展数据图 9a、b)。为了确定BSH是否是BA-MCY去结合所必需的,我们测试了定殖卵形拟杆菌ATCC 8483(野生型Bo)或敲除了编码BSH基因BO_02350的卵形拟杆菌突变株(Δbsh Bo)的悉生小鼠。我们发现,在定殖野生型Bo的悉生小鼠中,CDCA-d5-MCY 部分被去结合,而在定殖突变株Δbsh Bo的小鼠中则未被去结合(图3f),在这些小鼠中,补充的CDCA-d5-MCY像在无菌小鼠中一样仅转化为氧化的CDCA-d4/5-MCYO(2)(扩展数据图9c)。这些结果表明,肠道微生物群的BSH能够使BA-MCY结合物去结合,尽管其效率低于相应的牛磺酸结合物(图3f),并且在缺乏微生物群的情况下,BA-MCY 结合物会被宿主代谢为相应的BA-MCYO和BA-MCYO2衍生物(图3g)。
BA-MCY在体外表现为FXR拮抗剂
为了对胆汁酸-甲基半胱胺(BA-MCY)结合物进行功能评估,我们聚焦于法尼醇X受体(FXR),它是脊椎动物体内胆汁酸(BAs)的主要内源性靶点之一。例如,广泛存在且保守的鹅去氧胆酸(CDCA)、胆酸(CA)和脱氧胆酸(DCA)等游离胆汁酸,以及胆酸的氨基酸结合物,都作为强效的FXR激动剂,对胆汁酸的生成起负调控作用。相比之下,目前尚不清楚是否存在任何保守的内源性FXR拮抗剂来促进胆汁酸的生成。
为了测试已鉴定出的BA-MCY结合物潜在的FXR激动剂或拮抗剂活性,我们根据它们在无特定病原体(SPF)小鼠血清样本中的相对丰度,并考虑到先前报道的相应游离胆汁酸的FXR激动剂活性,选择了四种衍生物:CA-MCY、CA-MCYO、βMCA-MCY和CDCA-MCY。我们使用全长人FXR蛋白与类固醇受体共激活剂肽(SRCP)衍生的核融合蛋白之间的蛋白质-蛋白质相互作用测定法,以激动剂和拮抗剂模式对这四种化合物进行了检测。在任何测试浓度下,所测试的结合物均未显示出激动剂活性(图4和扩展数据图10a、b),而CDCA-MCY、CA-MCY和βMCA-MCY则表现出强效的拮抗活性,针对GW4604介导的FXR激活,其半数抑制浓度(IC50)值分别为1.68、19.9和104.5μM(图4a、b和扩展数据图10a)。相比之下,CA-MCYO无活性,这表明硫氧化消除了拮抗活性(图4c)。CDCA-MCY还抑制了由CDCA和更强效的合成胆汁酸奥贝胆酸介导的FXR激活(扩展数据图10c-g)。同时,我们还测试了牛磺-β-鼠胆酸(TβMCA),之前有报道称它是一种作用较弱的、小鼠特异性的FXR拮抗剂。然而,在该测定中,在所测试的浓度范围内,TβMCA无活性(扩展数据图10h)。这些结果表明,BA-MCY结合物作为内源性FXR拮抗剂发挥作用,与游离胆汁酸作为FXR激动剂的作用形成互补。
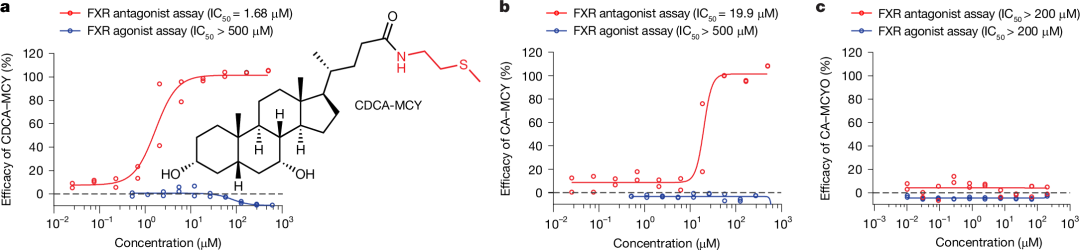
图4 | BA-MCY是FXR拮抗剂
a-c,胆汁酸-甲基半胱胺结合物(BA-MCYs)在体外作为法尼醇X受体(FXR)拮抗剂。采用基于细胞的蛋白质-蛋白质相互作用试验,以激动剂和拮抗剂两种模式对化合物进行测试。鹅去氧胆酸-甲基半胱胺(CDCA-MCY,图a)和胆酸-甲基半胱胺(CA-MCY,图b)对GW4604介导的FXR激活表现出强烈的FXR拮抗作用,而胆酸-甲基半胱胺氧化物(CA-MCYO,图c)则未显示出FXR拮抗作用。在该试验中,所测试的BA-MCY结合物均未表现出FXR激动作用(另见扩展数据图10)。每个浓度的试验均重复进行两次。
BA-MCY在体内调节FXR信号通路
肝脏中胆汁酸(BA)的生物合成受一个复杂的信号网络调控,该网络由肝脏和肠道中的法尼醇X受体(FXR)通过不同途径进行调节(图5a)。在肝脏中,FXR激动剂促进短异源二聚体伴侣(SHP)的表达,而SHP反过来会抑制Cyp7a1的表达,Cyp7a1编码一种细胞色素P450酶,该酶是胆汁酸合成第一步及限速步骤所必需的(图5a)。此外,SHP的表达会抑制Cyp8b1,Cyp8b1催化胆汁酸前体7α-羟基-4-胆甾烯-3-酮转化为7α,12α-二羟基-4-胆甾烯-3-酮,从而控制12α-羟基化胆汁酸(如胆酸(CA))和非12α-羟基化胆汁酸(如鹅去氧胆酸(CDCA))相对含量之间的平衡。相比之下,肠道中FXR的激活会促进回肠激素成纤维细胞生长因子15(FGF15;在人类中为FGF19)的产生,这是一种信号肽,通过肠肝循环到达肝脏,抑制Cyp7a1的表达。相反,FXR拮抗剂通过抑制SHP和FGF15/FGF19的表达,解除对Cyp7a1和Cyp8b1的抑制,从而促进胆汁酸的合成(图5a)。
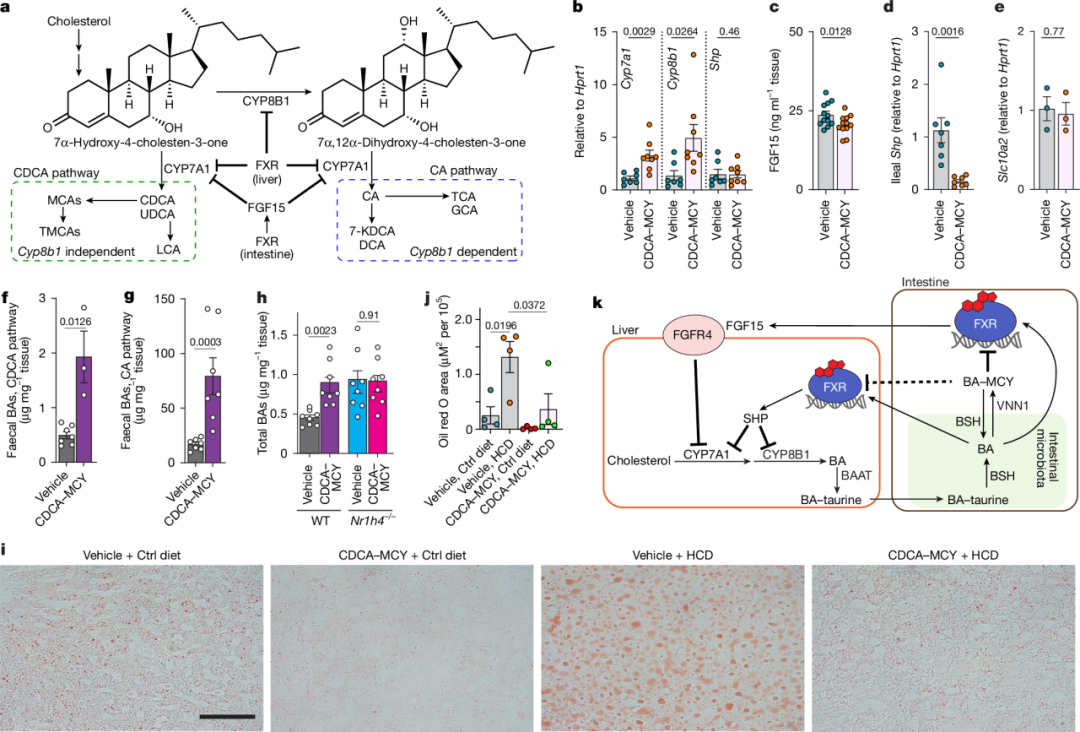
图5 | BA-MCY在体内调节胆汁酸的生物合成以及其在胆汁酸代谢中作用的模型
a,胆汁酸(BA)生物合成示意图,突出显示胆酸(CA)和鹅去氧胆酸(CDCA)的独立合成途径。蓝色和绿色突出显示的化合物(虚线框)分别属于CA和CDCA合成途径。
b,给予鹅去氧胆酸-甲基半胱胺(CDCA-MCY,n=4)或对照组(玉米油,n=3)的小鼠肝脏中,Shp、Cyp8b1和Cyp7a1基因表达量与内参基因Hprt1的比值。
c,给予CDCA-d5-MCY或对照组小鼠的血清成纤维细胞生长因子15(FGF15)水平(n=12)。
d,给予CDCA-d5-MCY或对照组小鼠小肠中,Shp基因表达量与Hprt1的比值(n=7)。
e,给予CDCA-d5-MCY或对照组小鼠小肠中,Slc10a2基因表达量与Hprt1的比值(n=3)。
f,g,连续14天给予CDCA-MCY或CDCA-d5-MCY的小鼠粪便中内源性生成胆汁酸的丰度。图f展示了CDCA衍生胆汁酸的总量(对照组n=7,给予CDCA-d5-MCY的小鼠n=3),图g展示了CA衍生胆汁酸的总量(对照组n = 7,给予CDCA - MCY的小鼠n = 4,给予CDCA - d5 - MCY的小鼠n = 3)。
h,连续14天每天给予野生型(WT)或Nr1h4基因敲除(Nr1h4−/−)小鼠CDCA-d5-MCY后,粪便中内源性生成胆汁酸的丰度。展示了胆汁酸的总量(对照组n=8,给予CDCA-MCY的小鼠n=8)。
i,在所示条件下处理的小鼠肝脏切片油红O染色的代表性显微照片。小鼠喂食对照饮食(Ctrl,载体组n=4,CDCA-MCY组n=4)或高胆固醇饮食(HCD,载体组n=4,CDCA-MCY组n=4)。CDCA-MCY通过灌胃给药,剂量为50mg/kg体重,持续2周。比例尺,50μm。
j,图i中油红O染色平均测量面积。
k,胆汁酸-甲基半胱胺结合物(BA-MCYs)通过法尼醇X受体(FXR)依赖途径调节胆汁酸代谢的模型。图k改编自参考文献60。数据以平均值±标准误表示(b-h,j)。P值采用未配对双侧Student t检验并经Welch校正(b-h)或单因素方差分析并经Tukey校正(j)计算得出。
为了确定BA-MCYs是否在体内影响胆汁酸生物合成的法尼醇X受体依赖性调节,我们通过灌胃给无特定病原体(SPF)小鼠补充鹅去氧胆酸-甲基半胱胺(CDCA-MCY),该物质在我们的体外FXR拮抗剂试验中显示出最高的效力(图4a)。基因表达分析表明,肝脏中Cyp7a1和Cyp8b1的表达显著增加(图5b)。此外,我们发现补充CDCA - MCY的小鼠回肠Shp mRNA和血清FGF15水平显著降低(图5c、d),这表明Cyp7a1表达的增加部分归因于对肠道FXR-FGF15通路的拮抗作用。Cyp8b1表达的增加在很大程度上独立于肠道FXR-FGF15通路,这表明肝脏FXR可能也受到CDCA-MCY补充的影响,或者其他通路有助于FXR依赖性的回肠-肝脏信号传导。由于我们发现BA-MCY的产生可能依赖于从回肠重吸收游离胆汁酸,我们还测试了补充CDCA-MCY是否会影响Slc10a2的表达,Slc10a2是介导胆汁酸从肠道重吸收的转运体;然而,Slc10a2的表达没有变化(图5e)。
为了测量CDCA-MCY在体内对胆汁酸生成的影响,我们使用稳定同位素标记的CDCA-d5-MCY进行了额外的补充研究。使用标记的CDCA-d5-MCY避免了因补充的CDCA-MCY去结合和进一步代谢而产生的潜在混淆影响,因为它使我们能够明确区分来自补充的、标记的CDCA-d5-MCY的胆汁酸和新生成的、未标记的胆汁酸(扩展数据图8e)。对补充CDCA-d5-MCY的动物胆汁酸水平的定量分析显示,粪便样本中未标记的CDCA衍生胆汁酸大幅增加(图5f和扩展数据图11a)。同样,补充CDCA-d5-MCY或CDCA-MCY的动物粪便样本中CA衍生胆汁酸的水平也有所增加(图5g和扩展数据图11b)。在使用抗生素处理(ABX)的小鼠中,补充CDCA-d5-MCY也会增加粪便胆汁酸水平(扩展数据图11c)。鉴于补充小鼠肝脏和血清中未标记胆汁酸的水平没有显著变化(扩展数据图11d-g),粪便中CA家族和CDCA家族胆汁酸排泄量的大幅增加表明,补充CDCA-MCY的动物胆汁酸生成强烈上调,这与胆汁酸生物合成基因表达的增加一致(图5b)。
接下来,我们测试了补充CDCA-MCY导致的胆汁酸合成上调是否实际上依赖于FXR。我们发现,补充CDCA-MCY会增加野生型(WT)小鼠粪便中的胆汁酸丰度,但不会增加FXR缺陷型(Nr1h4−/−)小鼠粪便中的胆汁酸丰度(图5h)。补充CDCA对WT和Nr1h4−/−小鼠肝脏和血清中的胆汁酸水平没有显著影响(扩展数据图12a、b),这与我们最初的补充研究结果一致(扩展数据图11d-g)。这些数据表明,补充CDCA-MCY以FXR依赖的方式增加胆汁酸生物合成。
鉴于肠道FXR拮抗剂已被证明可减轻肥胖小鼠模型中的肝脂肪变性,我们探究补充CDCA-MCY是否能改善喂食高胆固醇饮食(HCD)小鼠肝脏中的脂质积累。肝脏组织学和油红O染色显示,与未处理的HCD喂食小鼠相比,补充CDCA-MCY的HCD喂食小鼠肝脏脂质积累显著减少(图5i、j),这与之前关于作为肠道FXR拮抗剂的合成化合物的研究结果一致。当以低十倍的剂量供应CDCA-MCY时,我们观察到了类似的效果(扩展数据图12c、d)。
总之,我们的结果支持这样一个模型:宿主衍生的BA-MCY结合物作为肠道FXR拮抗剂,平衡微生物群衍生的游离胆汁酸的FXR激动活性,这是在肝胆系统内微调胆汁酸信号传导的调节回路的一部分(图5k)。
作者简介
美国康奈尔大学Tae Hyung Won、Mohammad Arifuzzaman、Christopher N. Parkhurst为本文的第一作者,教授David Artis和Frank C. Schroeder为本文通讯作者。
Tae Hyung Won(第一作者)

Tae Hyung Won,2024年9月至今,嘉泉大学药学院,助理教授;2015年9月-2024年8月康奈尔大学Boyce Thompson研究所,博士后研究员;2015年3月-2015年6月首尔大学天然物科学研究所,资深研究员。
Tae Hyung Won的研究室运用基于HPLC-HRMS的非靶向代谢组学以及二维核磁共振(2D NMR)分析技术,致力于从动物、植物、微生物等多种模式生物中探索生理活性物质。目前主要开展的研究有肠-脑轴的肠道微生物代谢组研究,以及真菌次生代谢产物相关研究,旨在阐明生理活性物质的化学生物学作用机制。研究室官网:https://sites.google.com/view/wonlab。
Mohammad Arifuzzaman(第一作者)

Mohammad Arifuzzaman,威尔康奈尔医学院医学免疫学助理教授、医学博士后助理,Mohammad Arifuzzaman博士的研究重点是探究包括饮食和微生物群信号在内的环境因素如何调节免疫系统。他的研究项目融合了免疫学、微生物学和化学生物学,采用前沿的基于代谢组学的方法,将非靶向代谢组学、细菌遗传学和转录组学研究相结合。通过这种方法,阿里夫扎曼博士的实验室研究了营养、微生物代谢与宿主免疫系统之间的复杂相互作用,这对感染性疾病、炎症性疾病、代谢性疾病以及癌症的研究都具有重要意义。
Christopher N. Parkhurst(第一作者)

Christopher N. Parkhurst,2021年至今,威尔康奈尔医学院医学科讲师;2020-2021年,威尔康奈尔医学院医学科研究员,2016-2017年,威尔康奈尔医学院医学科临床助理。
David Artis(通讯作者)

David Artis教授,吉尔·罗伯茨炎症性肠病研究所所长、Michael Kors免疫学及医学教授、微生物学与免疫学教授。David Artis实验室的目标是弄清楚控制身体屏障表面免疫细胞稳态的调控机制。通过运用微生物定植、病原体感染和慢性炎症等多种模型,David Artis实验室正在研究哺乳动物宿主基因以及源自共生微生物群落的信号,如何影响皮肤、肺部和肠道中的先天性免疫反应和适应性免疫反应。2014年,David Artis博士转至威尔康奈尔医学院,担任Michael Kors免疫学教授,同时出任位于纽约市的威尔康奈尔医学院吉尔·罗伯茨炎症性肠病研究所所长。阿蒂斯博士开展了一项研究项目,专注于剖析在健康与疾病状态下,调节屏障表面固有免疫和适应性免疫细胞功能的信号通路。他的研究项目还致力于将临床前模型的研究成果转化为针对免疫介导疾病的患者研究。David Artis博士的研究得到美国国立卫生研究院(NIH)、克罗恩病与结肠炎基金会(CCFA)和伯勒斯·韦尔科姆基金会(BWF)的资助,他还荣获了美国免疫学家协会(AAI)、克罗恩病与结肠炎基金会(CCFA)和国际黏膜免疫学会(ICIS)颁发的青年研究员奖、科尔顿奖(Colyton Prize)、斯坦利·科恩奖(Stanley Cohen Prize)以及美国免疫学家协会-BD(AAI - BD Biosciences)研究员奖。
Christopher N. Parkhurst(通讯作者)

Christopher N. Parkhurst教授,哈佛大学医学院博士后、康奈尔大学博士后、汉堡大学博士&硕士。其研究兴趣在于鉴定次生代谢产物(天然“小分子”)的结构,并阐明其生物学功能。目前其实验室开展了三个主要项目,采用三种不同方法,将小分子的分子结构与生物学功能或活性联系起来。
翻译:曾美尹,中国农科院基因组所,硕士在读
审核:朱志豪,广东医科大学,基因组所联合博士后
终审:刘永鑫,中国农科院基因组所,研究员/博导
排版:荀佳妮,中国农科院基因组所,生物信息学硕士在读
宏基因组推荐
本公众号现全面开放投稿,希望文章作者讲出自己的科研故事,分享论文的精华与亮点。投稿请联系小编(微信号:yongxinliu 或 meta-genomics)
猜你喜欢
iMeta高引文章 fastp 复杂热图 ggtree 绘图imageGP 网络iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文

















 1万+
1万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









