






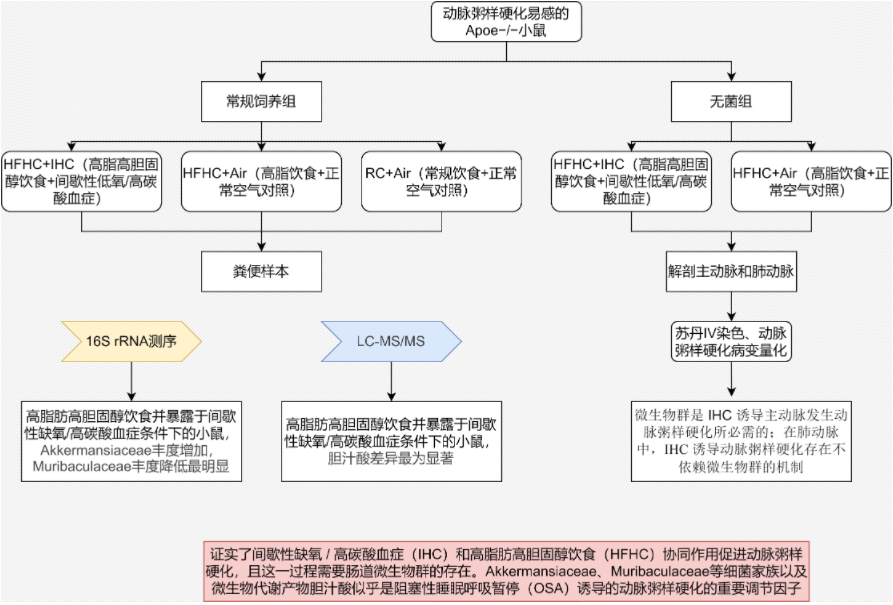
01 研究背景
阻塞性睡眠呼吸暂停(OSA)是一种常见的睡眠障碍,表现为睡眠中反复发生上呼吸道塌陷,导致间歇性低氧(IH)和高碳酸血症(HC)(合称IHC)。全球约10亿人受OSA影响,肥胖人群患病率高达30-70%,并且OSA是动脉粥样硬化的独立风险因素,与心肌梗死、中风等密切相关。肠道菌群及其代谢物在动脉粥硬化中的作用已经被初步探索,但OSA通过菌群促进动脉粥硬化的机制尚不明确。本研究通过多组学方法,探究高脂高胆固醇饮食(HFHC)和IHC如何通过肠道菌群及其代谢物协同促进动脉粥样硬化,同时研究了肠道微生物群与OSA诱导的动脉粥样硬化之间的因果关系。
02 技术路线
03 研究内容
1.IHC加重了HFHC饮食下动脉粥样硬化斑块的形成
为了研究高脂肪高胆固醇(HFHC)饮食和间歇性缺氧 / 高碳酸血症(IHC)对动脉粥样硬化病变发展的影响,研究人员给 SPF(无特定病原体)Apoe⁻/⁻小鼠喂食 HFHC 饮食或普通饲料(RC)10 周。同时,让这些小鼠暴露在室内空气(Air)或间歇性缺氧 / 高碳酸血症(IHC)环境中,以研究阻塞性睡眠呼吸暂停(OSA)潜在的致动脉粥样硬化作用。接受 HFHC 饮食的小鼠比在室内空气中喂食 RC 的小鼠出现了明显更多的动脉粥样硬化病变(图 1)。结果表明间歇性缺氧 / 高碳酸血症(IHC)与 HFHC 协同作用,影响动脉粥样硬化的发展。为了评估饮食和环境暴露对病变发展的影响,研究人员测量了小鼠的体重和食物摄入量。HFHC 饮食导致小鼠体重增加更多;而 IHC 暴露虽然使小鼠体重增加减少,但却加剧了动脉粥样硬化病变的形成。此外,小鼠对不同饮食的摄入量存在差异,且 IHC 应激会抑制小鼠的食物摄入。
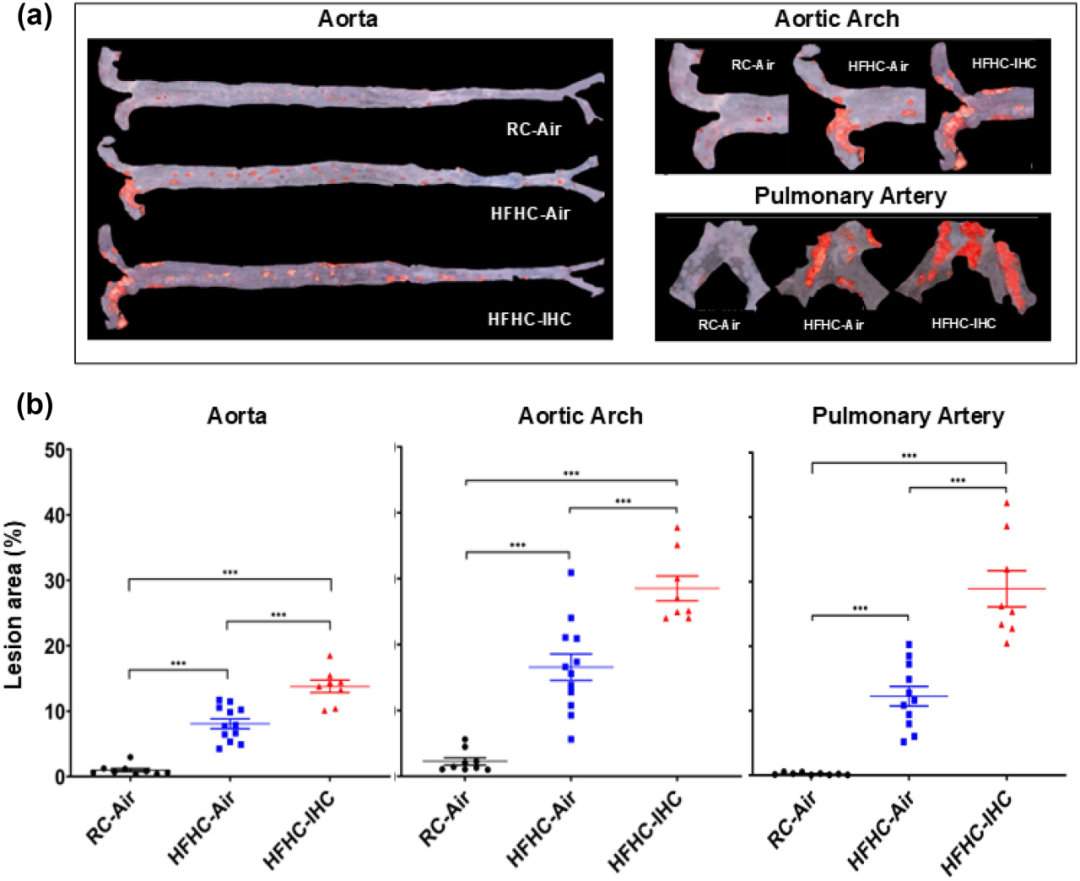
图1 无特定病原体(SPF)的载脂蛋白 E 基因敲除(Apoe⁻/⁻)小鼠在接受为期 10 周的高脂肪高胆固醇饮食且伴有或不伴有间歇性缺氧 / 高碳酸血症情况下的动脉粥样硬化病变情况
2.高脂肪高胆固醇饮食(HFHC)和无间歇性缺氧 / 高碳酸血症(IHC)都会改变肠道微生物群的组成
为了解HFHC和IHC对肠道微生物群组成的变化,在研究期间(10周)每3-4天收集所有小鼠的连续粪便样本,采用16S V4扩增子测序分析。β 多样性(图 2A、B)发现,在初始时间点,当所有动物都食用常规饲料(RC)时,各组之间没有显著差异,但随着时间的推移,差异逐渐累积。此外,对 α 多样性指标的检查也揭示了饮食对微生物组组成的强烈影响,在这些差异中,毛螺菌科(Muribaculaceae)和阿克曼菌科(Akkermansiaceae)是两个关键的重叠细菌家族,它们呈现出相反的富集趋势(图2C、D)。基于之前的研究成果 ,以及利用稳健主成分分析(RPCA)排名(图 2A)、时间张量分解(TEMPTED)排名(图 2B)和 ANCOM - BC 判别特征(图 2C、D)从本数据集获得的差异丰度结果,作者研究了一个相关的自然对数比,用于比较属于毛螺菌科和阿克曼菌科的扩增子序列变异(ASV)的读数(图 2E)。使用线性混合效应模型分析时,该对数比显示随着时间推移,所有三组之间存在显著差异。高脂肪高胆固醇饮食并暴露于 IHC 条件下的小鼠,尤其在实验结束时,是最有可能拥有比毛螺菌科更多阿克曼菌科的一组(图 2F)。
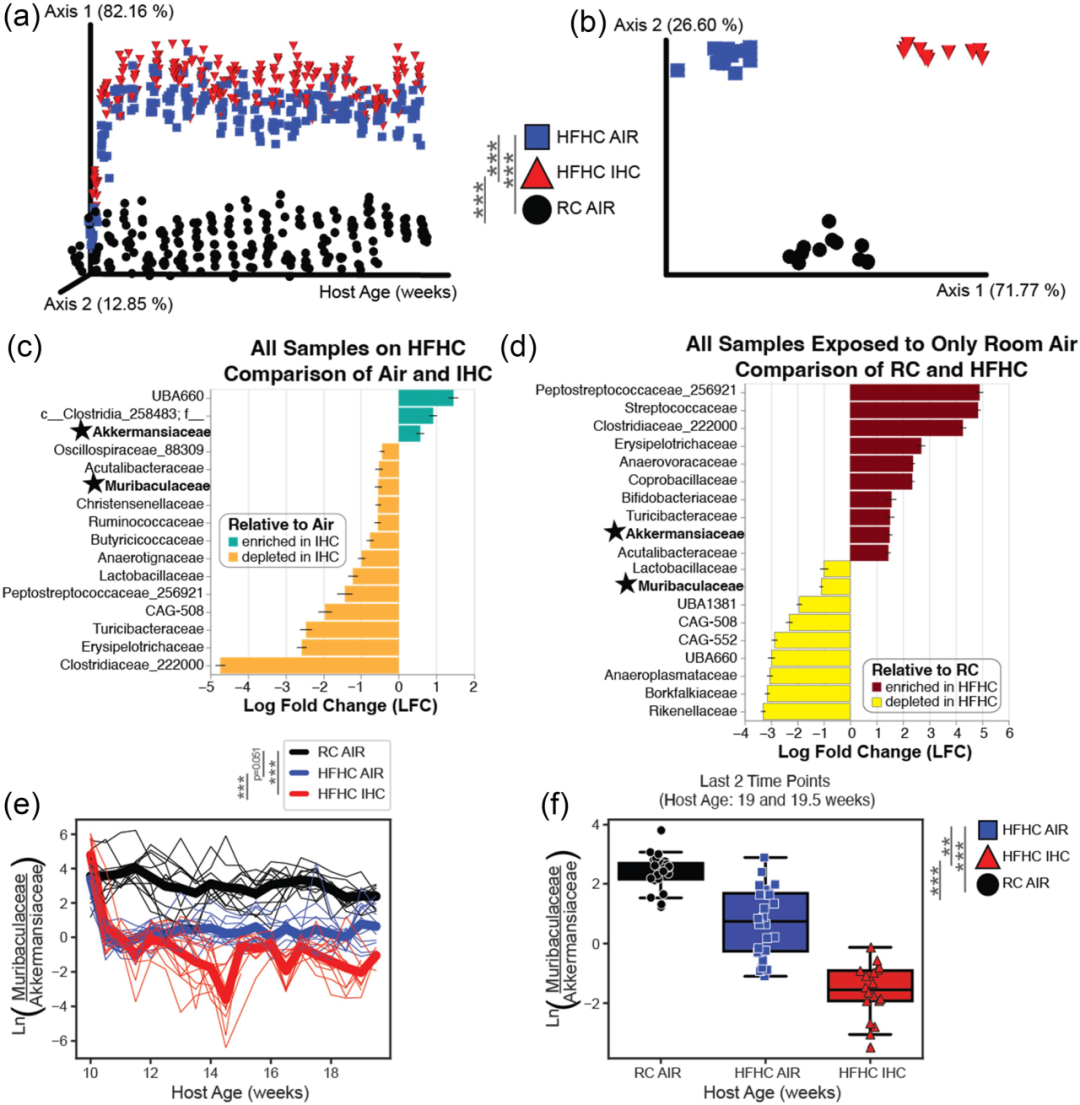
图2 无特定病原体(SPF)的载脂蛋白 E 基因敲除(Apoe −1−)小鼠在接受为期 10 周的高脂肪高胆固醇饮食(HFHC)且有无间歇性缺氧 / 高碳酸血症(IHC)处理情况下的微生物组分析
3.高脂肪高胆固醇饮食(HFHC)和间歇性缺氧 / 高碳酸血症(IHC)深刻影响粪便代谢谱
非靶向代谢组学分析显示,通过主成分分析(PCA)发现,饮食和空气暴露均可使粪便代谢物明显区分开来(图 3A)。差异结果显示,HFHC 饮食使 200 多种已注释或推测注释的胆汁酸增加,包括胆酸(CA)、脱氧胆酸(DCA)、石胆酸、牛磺胆酸等,以及最近描述的微生物胆汁酸 Ile/Leu - CA等。此外,HFHC 饮食还使最近描述的长链 N - 酰基酰胺增加,如 Leu - C16:0、Lys - C16:0、Phe - C16:0等,以及几种磷脂酰胆碱,包括 PC(16:0/0:0)、PC (16:1/0:0)、PC(16:0/20:5)等,还有鞘氨醇(C17 和 C18)。另一方面,HFHC 饮食似乎降低了 50 多种未注释的单羟基、二羟基、三羟基、四羟基和五羟基胆汁酸的丰度,以及几种短链 N - 酰基酰胺,100多种二肽和三肽。在第一个时间点之后,当所有动物都在接受 RC 饮食时,这些提取特征的对数比能够区分接受或未接受 HFHC 饮食的动物(图 3B)。
IHC导致100多种已注释和推测的胆汁酸增加,包括牛磺石胆酸、牛磺脱氧胆酸、牛磺胆酸等物质。IHC 还降低了几种胆汁酸的丰度。空气组相关特征与HFHC 、IHC组相关特征的对数比,显著区分了所关注的三组(图 3C)。分子特征子网络以展示一些导致组间差异的分子,如胆汁酸、磷脂酰胆碱和 N - 酰基酰胺(图 3D)。由于HFHC 促进了动脉粥样硬化的形成,而 IHC 进一步加速了这一过程,因此研究人员识别出了441个具有一致或相反变化模式的特征子集,因为这些分子可能在生物学上与相关的动脉粥样硬化发展高度相关。研究人员还通过单变量分析对具有代表性的感兴趣分子进行了研究,以突出组间差异(图 3E)。
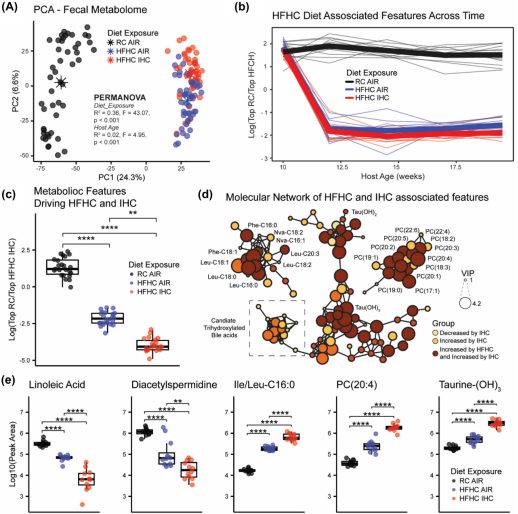
图3 无特定病原体(SPF)的载脂蛋白 E 基因敲除(Apoe−1−)小鼠在接受为期 10 周的高脂肪高胆固醇饮食(HFHC)且有无间歇性缺氧 / 高碳酸血症(IHC)处理情况下的代谢组学分析
4.Akkermansiaceae与Muribaculaceae的比例影响代谢物谱
利用前文所讨论的微生物组和代谢组的对数比,绘制了散点图来观察样本聚类情况(图 4A)。当对数比偏向与常规饮食-空气条件相关的代谢物,且更倾向于Muribaculaceae的扩增子序列变异(ASV)时,多数样本为常规饮食-空气组样本。相反,当对数比偏向与高脂肪高胆固醇饮食(HFHC)-间歇性缺氧 / 高碳酸血症(IHC)条件相关的代谢物,且更倾向于Akkermansiaceae的 ASV 时,多数样本为 HFHC - IHC 组样本。虽然观察到所有三组都有明显的聚类情况,但饮食似乎有着较大影响。此外,还可以看到一种显著且较强的线性关系(图 4B)。总之,这些结果表明,存在一种明显的微生物组和代谢物特征,该特征似乎与对饮食和暴露类型的响应相关。
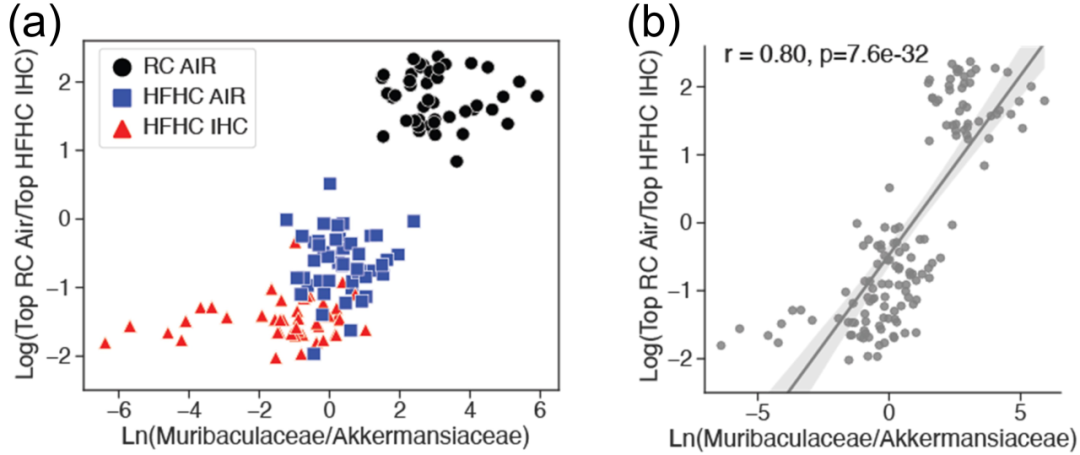
图4 微生物组和代谢组对数比值的比较
5.微生物群缺失可减轻高脂肪高胆固醇饮食(HFHC)和间歇性缺氧 / 高碳酸血症(IHC)在主动脉中诱发的动脉粥样硬化,但对肺动脉中的动脉粥样硬化并无此作用
为了探究肠道微生物群是否在高脂肪高胆固醇饮食(HFHC)和间歇性缺氧 / 高碳酸血症(IHC)诱导的动脉粥样硬化中起作用,对无菌(GF)的小鼠像无特定病原体(SPF)动物一样,在空气环境或间歇性缺氧 / 高碳酸血症(IHC)环境下,喂食相同的高脂肪高胆固醇饮食,持续10周。结果发现在高脂肪高胆固醇饮食(HFHC)诱导下,无菌小鼠主动脉(主要是不包括主动脉弓的主动脉主干部分)中的病变显著少于无特定病原体小鼠(图 5A、B)。当暴露于间歇性缺氧 / 高碳酸血症(IHC)环境时,无菌Apoe−/−小鼠相较于无特定病原体对照组,仅在主动脉主干部分出现统计学意义上更少的病变,在其他被检测的血管区域则未出现这种情况(图 5C、D)。这些数据表明,微生物定殖对不同血管床中动脉粥样硬化的调节作用存在差异。
如图 1 所示,在高脂肪高胆固醇饮食(HFHC)存在的情况下,间歇性缺氧 / 高碳酸血症(IHC)促进了无特定病原体(SPF)的载脂蛋白 E 基因敲除(Apoe −1−)小鼠的主动脉、主动脉弓以及肺动脉中动脉粥样硬化的形成。然而,对于喂食高脂肪高胆固醇饮食的无菌(GF)Apoe −1−小鼠,IHC 对动脉粥样硬化的影响仅在肺动脉中被检测到,在主动脉中则未检测到(图6)。这些数据表明:(1)微生物群是 IHC 诱导主动脉发生动脉粥样硬化所必需的;(2)在肺动脉中,IHC 诱导动脉粥样硬化存在不依赖微生物群的机制。
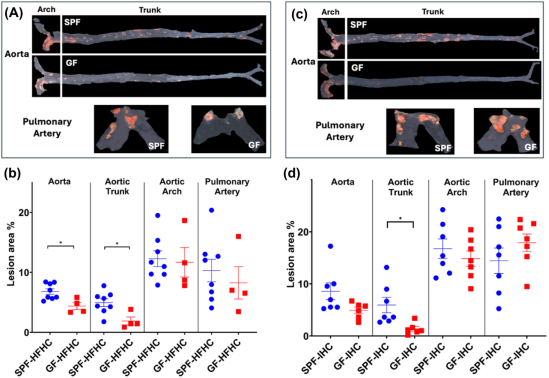
图5 无菌(GF)和无特定病原体(SPF)的载脂蛋白 E 基因敲除(Apoe−1−)小鼠在经过 10 周高脂肪高胆固醇饮食(HFHC)或间歇性缺氧 / 高碳酸血症(IHC)暴露后的动脉粥样硬化病变情况
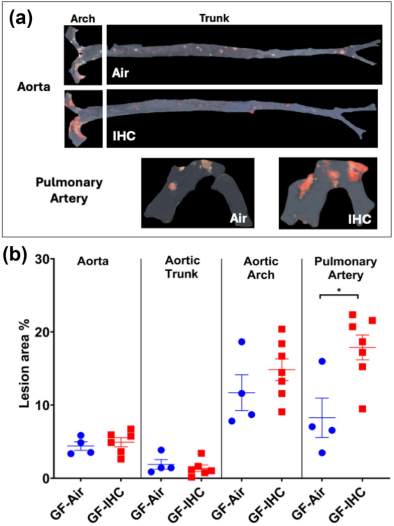
图6 无菌(GF)载脂蛋白 E 基因敲除(Apoe−1−)小鼠经过 10 周间歇性缺氧 / 高碳酸血症(IHC)及空气处理后的动脉粥样硬化病变情况
04 研究结论
综上所述,该研究证明了间歇性缺氧 / 高碳酸血症(IHC)和高脂肪高胆固醇饮食(HFHC)协同促进动脉粥样硬化,且这一过程需要肠道微生物群的存在。阿克曼菌科和毛螺菌科细菌家族,以及微生物代谢产物胆汁酸,似乎是阻塞性睡眠呼吸暂停(OSA)诱导的动脉粥样硬化的重要调节因子。这些研究成果有助于我们更好地理解饮食、微生物群与 IHC/OSA 诱导的动脉粥样硬化之间的机制联系,并为未来预防和治疗 OSA 诱导的动脉粥样硬化的治疗方法提供了依据。例如,可通过使用益生元、益生菌或合生元纠正肠道菌群失调,或使用法尼醇 X 受体(FXR)/ 武田 G 蛋白偶联受体 5(TGR5)激动剂或拮抗剂调节胆汁酸信号传导,并结合饮食调整来进行干预。

















 1843
1843

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









