






点击蓝字 关注我们
母体肠道微生物源性大豆苷元可预防孕期强的松暴露所致雌性子代骨质疏松症

iMeta主页:http://www.imeta.science
研究论文
● 原文: iMeta (IF 23.8)
● 原文链接: https://onlinelibrary.wiley.com/doi/10.1002/imt2.70037
● DOI: https://doi.org/10.1002/imt2.70037
● 2025年4月28日,武汉大学汪晖、陈廖斌等在iMeta在线发表了题为“Maternal gut microbiota-derived daidzein prevents osteoporosis in female offspring following prenatal prednisone exposure”的文章。
● 本研究使用16S rRNA测序、代谢组和转录组等多组学联合分析,通过系统的体内和体外验证实验,建立了母体肠道微生物组—表观遗传调控—子代骨发育轴,多维度探讨了母体肠道菌群稳态、表观遗传调控机制与子代成骨分化功能之间的三元关系,进一步提出孕期母体补充肠道菌群代谢物大豆苷元可有效预防子代骨质疏松症发生,并强调了孕期优化母体肠道菌群的重要性。
● 第一作者:马翅、何航元、王鹍鹏
● 通讯作者:陈廖斌(lbchen@whu.edu.cn)、汪晖(wanghui19@whu.edu.cn)
● 合作作者:郭娟娟、刘亮、陈玉婷、李斌、肖浩、李旭峰、路小倩、王婷婷、文印宪
● 主要单位:武汉大学中南医院、武汉大学基础医学院、发育源性疾病湖北省重点实验室、武汉市中医院
亮 点
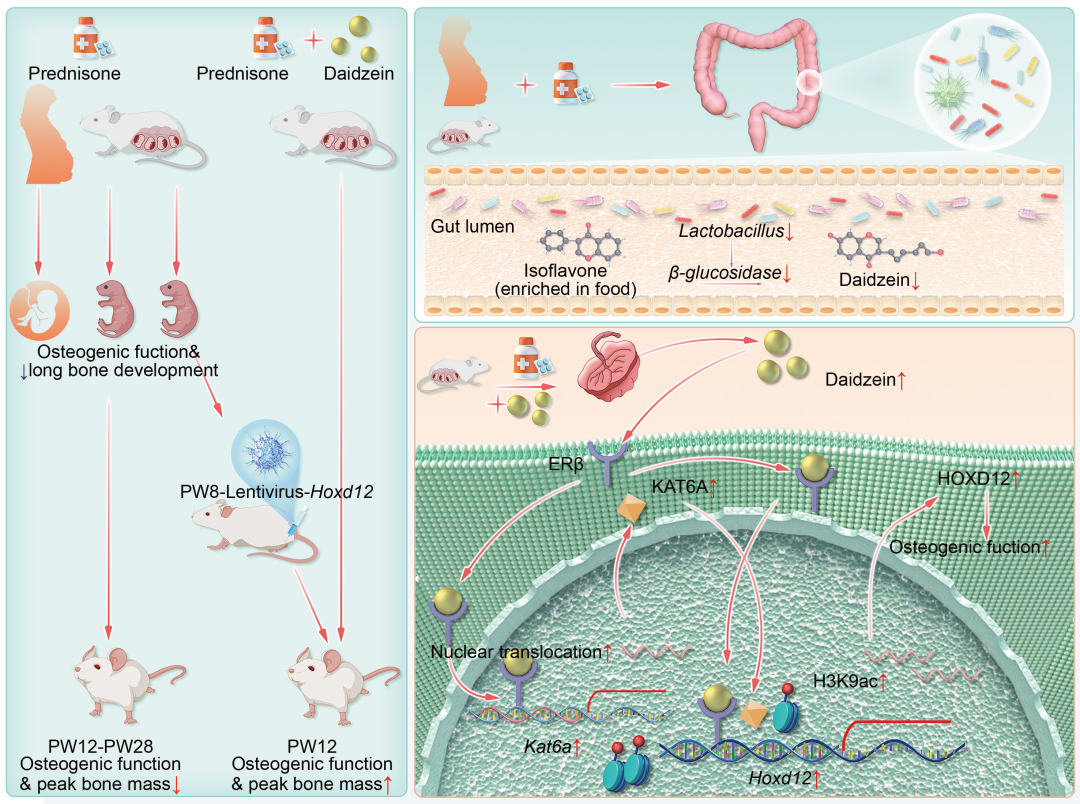
● 孕期强的松治疗(PPT)或孕期强的松暴露(PPE)会导致女性或雌性子代长骨发育不良、峰值骨量降低和成年期骨质疏松症易感;
● 母体肠道菌群代谢物大豆苷元(DAI)介导了PPE所致的雌性子代骨质疏松症易感;
● DAI通过ERβ/KAT6A信号表观遗传调控Hoxd12表达介导了PPE所致的子代成骨功能低下及骨质疏松症易感;
● DAI是PPE所致雌性子代多器官发育不良及相关疾病易感的早期有效预防靶标。
摘 要
孕期糖皮质激素暴露会影响子代远期健康及疾病发生风险,但其母体肠道菌群机制尚未阐明。在本研究中,临床研究与动物实验发现,孕期强的松治疗(PPT)或孕期强的松暴露(PPE)均可性别差异性地影响雌性子代长骨发育,导致其峰值骨量(PBM)降低及骨质疏松症(OP)易感。同时运用多组学技术与菌群移植实验证实,PPE可导致母体肠道菌群组成改变及其代谢物大豆苷元(DAI)显著减少,上述改变可抑制Hoxd12表达及成骨功能,导致雌性子代PBM降低。细胞水平证实PPE雌性子代原代骨髓间充质干细胞(PPE-BMSCs)的Hoxd12表达降低及成骨分化功能抑制,而补充DAI可通过ERβ/KAT6A信号增加Hoxd12启动子区H3K9ac水平及表达,并增强成骨分化功能。整体水平进一步证实,孕期母体补充DAI能明显逆转PPE所致的雌性子代OP易感及其他多器官(如肝脏、海马、卵巢、肾上腺)功能改变。综上,PPE通过改变母体肠道菌群组成及降低其代谢物DAI水平,诱导雌性子代多器官发育不良及相关疾病易感(如成骨不良及OP易感),而孕期母体补充DAI是上述不良影响的潜在有效预防策略。
视频解读
Bilibili:https://www.bilibili.com/video/BV1TyGtzgELZ/
Youtube:https://youtu.be/-qfJGhPg3BQ
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
“健康与疾病的发育起源(Developmental Origins of Health and Disease,DOHaD)”学说的提出,强调了生命早期不良事件暴露对子代远期健康及疾病易感性的长期影响。肠道菌群又称“第二大人类基因组”,在机体营养吸收、免疫系统调节和内环境平衡中具有重要作用。越来越多的研究发现,孕期母体肠道菌群参与子代胃肠道、心血管、免疫和神经等多系统发育。然而,近期研究发现,孕期不良环境(药物暴露、高脂饮食等)可通过改变母体肠道菌群组成,导致子代出生后哮喘、过敏和肥胖等多疾病易感。Kimura等研究也证实,无菌孕鼠的子代成年后患代谢综合征的风险明显增加。综上提示,母体肠道菌群是影响子代发育的重要因素,孕期不良环境可通过改变母体肠道菌群组成,继而增加子代出生后多疾病发生风险。
子代出生后骨量水平与孕期不良环境密切相关,如孕期全氟烷基等物质暴露可影响子代出生后骨量水平。本室前期研究也证实,孕期咖啡因、尼古丁、乙醇和地塞米松等外源物暴露,会导致子代成年期峰值骨量(peak bone mass,PBM)低下和骨质疏松症(osteoporosis,OP)易感。此外,母体肠道菌群与骨量之间的关系也极为密切。研究发现,低骨量无菌孕鼠(C57BL/6)接受高骨量孕鼠(C3H/HeN)的肠道菌群移植后,其子代呈现高骨量表型;反之,高骨量无菌孕鼠接受低骨量孕鼠的肠道菌群移植后,其子代表现为低骨量表型。提示,孕期不良环境所致母体肠道菌群改变可能是影响子代骨发育与PBM累积的关键因素。
强的松(prednisone)是一种常见的人工合成糖皮质激素,常用于治疗妊娠期母体类风湿性关节炎、系统性红斑狼疮等自身免疫性疾病,也常用于改善胚胎着床和预防流产。2016年英国风湿病学会和2020年美国风湿病学会指南推荐,最低有效剂量强的松对于控制母体疾病活动度并为胎儿营造良好的母体环境至关重要,可用于妊娠各时期甚至全程。然而,临床研究发现,孕期强的松治疗(prenatal prednisone therapy,PPT)会导致胎盘功能不全、早产和低出生体、唇和/或腭裂等风险增加。动物实验也表明,孕期强的松暴露(prenatal prednisone exposure,PPE)会导致宫内胎鼠卵巢发育毒性、关节软骨质量低下,甚至会引起成年期骨关节炎易感。综上提示,孕期强的松使用具有“双刃剑”效应:孕期强的松在治疗母体疾病的同时,可影响子代多器官发育,并导致成年疾病易感性增加。
研究发现,强的松可导致成年大鼠肠道菌群紊乱,艾森伯格氏菌和亚伯氏菌相对丰度降低,代谢产物短链脂肪酸降低。那么,孕期使用强的松是否会通过母体肠道菌群影响子代长骨发育及OP易感?其发生机制和防治策略?这些重要问题有待研究阐明。在本研究中,我们拟收集孕期有强的松用药史的子代股骨发育指标和母体粪便等临床数据或标本,并孕全程经口灌胃给予Wistar大鼠强的松0.25 mg/kg.d(相当于临床给药剂量)以构建PPE大鼠模型,证实PPE对母体肠道菌群组成变化、子代长骨发育状况及OP易感的影响。本研究通过检测及分析母体肠道菌群变化、表观遗传调控机制和子代成骨分化功能之间的三元关系,建立了一项全面且新颖的研究。同时,通过16S rRNA测序、代谢组及转录组等多组学分析,通过体内、外实验探究PPE所致OP易感的发生机制及防治靶标。此外,通过回顾性病例−对照研究、动物和细胞实验,多维度探究并验证上述科学问题。
结 果
PPT/PPE可致母体肠道菌群组成改变、子代长骨发育不良及PBM降低并存在明显的性别差异
为评估PPT对妊娠结局、宫内胎儿及骨发育的影响,我们通过回顾性病例−对照研究收集了223例产妇-新生儿病例数据(表S1)并进行组间分析。结果表明,PPT组早产儿有28例而对照组仅有1例;在剔除各组早产儿后,PPT组女、男性新生儿出生体重低于对照组。进一步基于文献方法检测发现,PPT组女胎股骨长度Z值、股骨长度/双顶径比值降低(图1A、1B),而男胎无明显改变。提示,PPT导致早产风险增加、低出生体重和女性胎儿长骨发育抑制。我们还收集了另一批妊娠中、晚期孕妇粪便(表S2),并进行肠道菌群16S rRNA检测。结果表明,与对照组相比,PPT组母体肠道菌群α多样性降低、β多样性发生改变(图1C、1D),其相对丰度在门水平发生改变,线性判别分析(linear discriminant analysis of effect size,LEfSe)显示,肠乳杆菌、艾克曼菌等有益菌相对丰度降低(图S1A、S1B)。典型关联分析(canonical correlation analysis,CCA)显示,在母体肠道菌群诸多影响因素中,强的松最能解释母体肠道菌群的整体变化(32.8%)(图S1C)。综上提示,PPT组母体肠道菌群组成发生改变,伴有女性胎儿长骨发育抑制。
与此同时,我们基于临床PPT方案开展动物实验,于GD0-20经口灌胃给予Wistar孕鼠0.25 mg/kg.d强的松(图1E),以证实上述临床发现并探究其远期效应−成年子代PBM变化。结果表明,与对照组相比,PPE组肠道菌群α多样性无显著变化、β多样性发生改变(图1F、1G),其相对丰度在门水平发生改变,且肠乳杆菌、丁酸梭菌等有益菌相对丰度降低(图S1D、S1E)。同时发现,PPE组雌性胎鼠股骨长度/初级骨化中心(primary ossification center,POC)长度/POC区域骨小梁周长/矿化面积均显著降低、破骨功能无明显变化(图1H−O),成年后(PW12和PW28)的PBM均持续降低(图1P−Y)。进一步发现,PPE组雌性子代出生前、后骨组织中成骨功能标志基因(Bsp、Alp、Runx2、Col1a1与Ocn)的mRNA表达均显著降低,RUNX2蛋白表达亦显著降低(图S2A−G)。然而,PPE组雄性子代出生前后骨发育及PBM均无明显改变(图S2H−V)。综上提示,PPE可致母体大鼠肠道菌群组成改变,同时引起雌性子代大鼠出生前后长骨发育不良及成年PBM降低,但对雄性子代无显著影响。
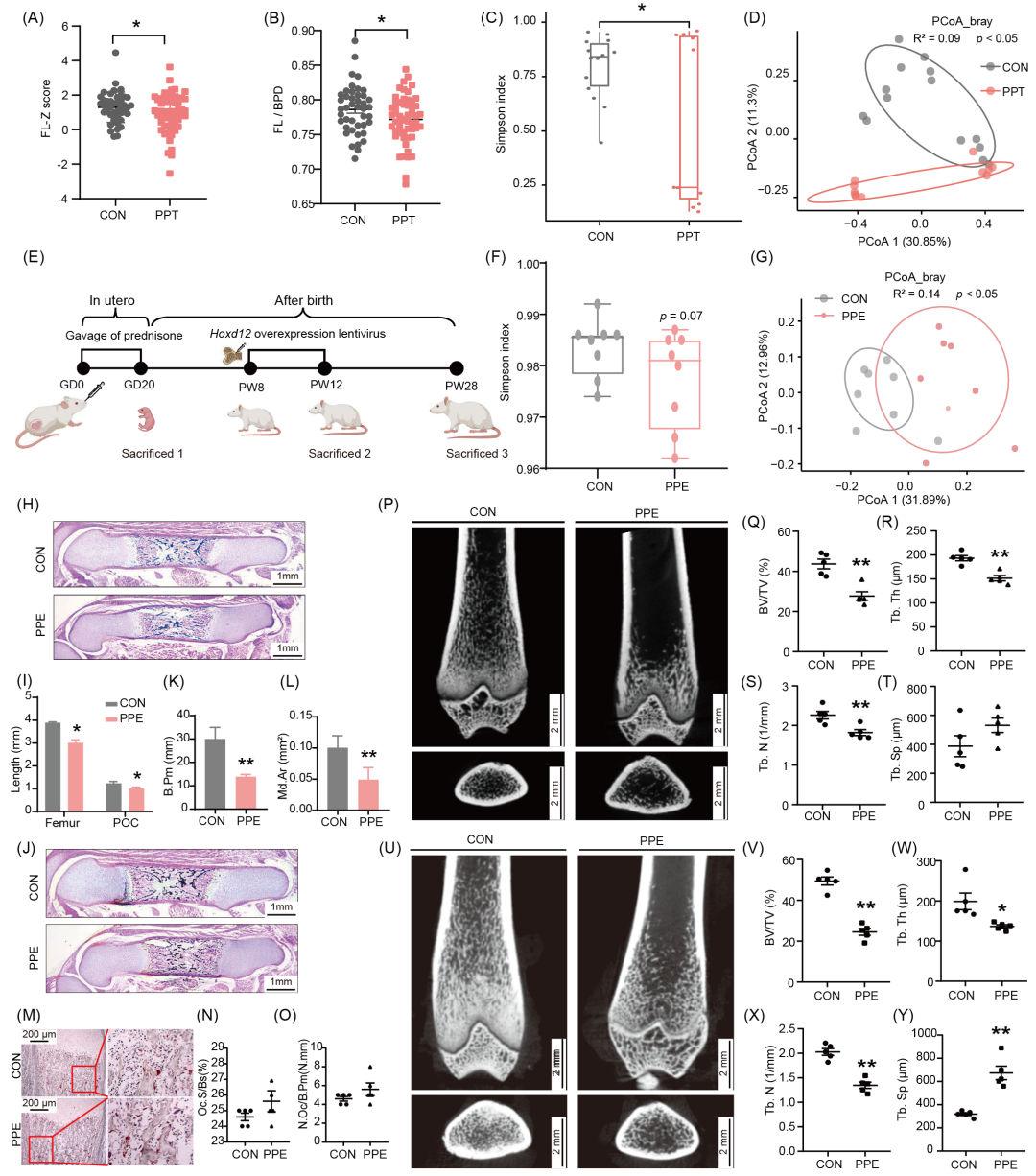
图1. PPT/PPE对雌性子代长骨发育和母体肠道菌群的影响
(A、B)临床研究中女胎FL-Z评分和FL/BPD比值;(C)临床孕妇肠道菌群Simpson指数图;(D)临床孕妇肠道菌群PCoA图;(E)构建PPE大鼠示意图;(F)孕鼠肠道菌群Simpson指数图;(G)孕鼠肠道菌群PCoA图;(H、I)雌性胎鼠股骨H&E染色和股骨总长及POC长度分析;(J−L)雌性胎鼠股骨Vonkossa染色和B.Pm及Md.Ar的定量分析;(M−O)雌性胎鼠股骨TRAP染色和Oc.S/Bs及N.Oc/B.Pm定量分析;(P−T)PW12雌性子代micro-CT及分析;(U−Y)PW28雌性子代micro-CT及分析。
母体肠道菌群代谢物DAI是PPE所致雌性子代大鼠成骨功能及PBM降低的干预靶标
为探究PPE母体肠道菌群组成改变与雌性子代成骨功能及PBM降低之间的关系,我们进行了母体肠道菌群移植实验(fecal microbiota transplant,FMT) (图2A、S3A、S3B)。与FMT-CON组相比,FMT-PPE组骨小梁稀疏、变薄、间隙增大,BV/TV、Tb.Th、Tb.N降低而Tb.Sp升高,且成骨功能标志基因(Alp、Runx2、Col1a1与Ocn)的mRNA与RUNX2蛋白表达均显著降低(图2B、2C、S3C−H)。提示,母体肠道菌群组成改变介导PPE所致雌性子代大鼠出生后成骨功能抑制及PBM降低。
为进一步探究PPE雌性子代大鼠成骨功能抑制的母体肠道菌群发生机制,我们对母鼠和雌性胎鼠血清进行了非靶向代谢谱进行筛选分析(图S4A−F)。结果显示,与对照组相比,PPE组母血和雌性胎血中大豆苷元(daidzein,DAI)水平显著降低(图2D),同时雌性胎鼠长骨组织中DAI水平也降低(图2E)。进一步结果表明,母体肠道中肠乳杆菌相对丰度与胎骨组织DAI水平呈正相关,PICRUSt功能预测显示PPE可导致L-精氨酸及乳酸合成/双歧杆菌代谢通路发生改变,酶学检测结果证实PPE组母鼠肠道菌群内肠乳杆菌中β-葡萄糖苷酶(β-glucosidase,β-GC)活性显著降低而β-半乳糖苷酶(β-galactosidase,β-GAL)活性无明显改变(图2F、2G、S4G−I)。提示,PPE所致母-胎血/组织中DAI水平的下降可能与母体肠道菌群中肠乳杆菌β-GC活性降低有关。最后,我们证实,孕期给予FMT-PPE组或PPE组母鼠补充DAI,均可显著逆转FMT-PPE或PPE所致雌性成年子代长骨组织DAI水平、成骨功能及PBM的降低(图2B、2C、2H−K、S3C−H、S5A−G)。提示,母体肠道菌群代谢物DAI是PPE所致雌性子代大鼠的成骨功能及PBM降低的干预靶标。
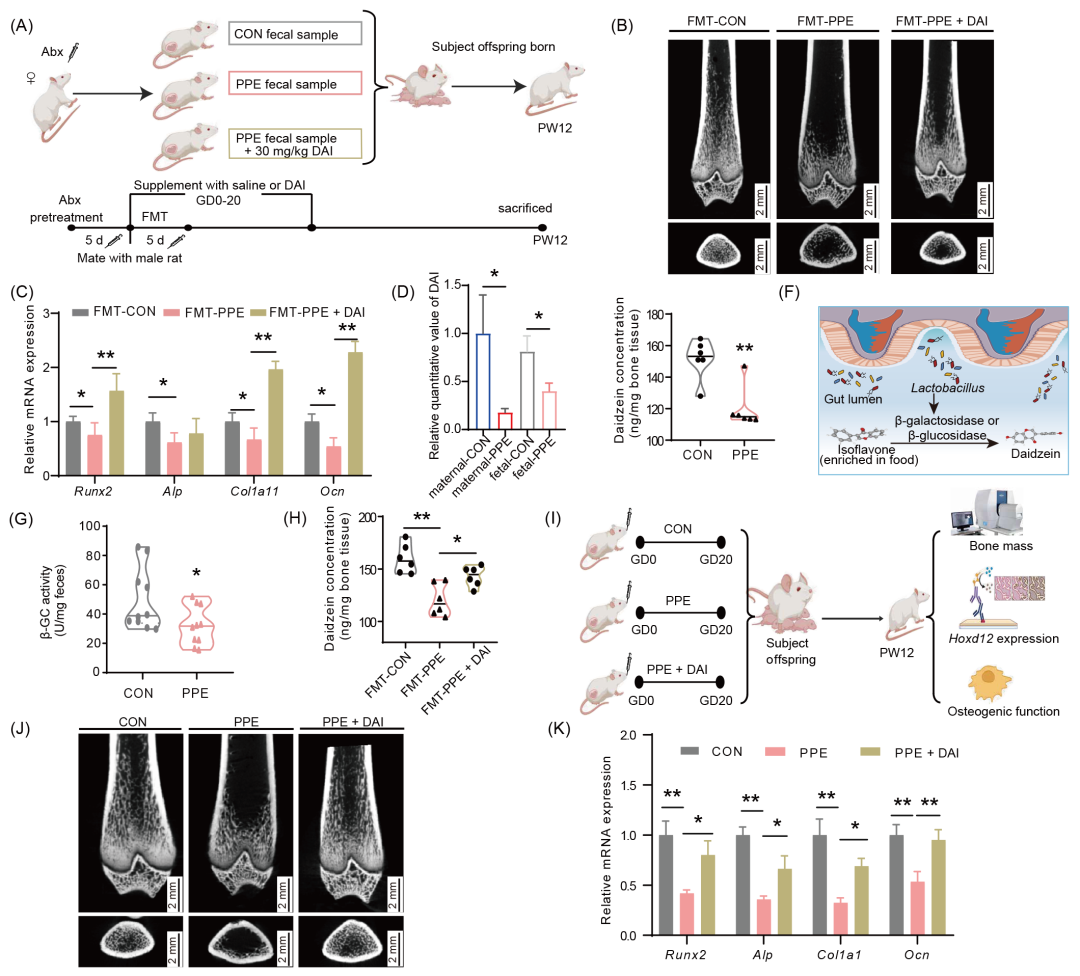
图2. 母体肠道菌群和DAI介导PPE所致雌性子代成骨功能和PBM低下
(A)母鼠FMT实验示意图;(B)PW12子代micro-CT图;(C)PW12子代成骨标志基因表达水平;(D)代谢谱结果中DAI相对定量分析;(E)雌性胎鼠骨组织中DAI水平;(F)富含异黄酮食物经β-GC/β-GAL水解形成DAI示意图;(G)母鼠肠道菌群β-GC活性;(H)胎鼠骨组织中DAI水平;(I)母体补充DAI实验示意图;(J)micro-CT检测PW12雌性子代PBM;(K)成骨标志基因的相对表达量。
Hoxd12是母体菌群代谢物DAI水平改变所致PPE雌性子代大鼠成骨功能抑制的关键靶标分子
孕期不良环境可通过影响子代骨骼组织中关键基因表达编程导致成年期OP等远期危害发生。我们对雌性胎鼠长骨组织进行转录组测序发现,在PPE组下调基因中,参与骨骼系统发育的同源盒转录因子D12(homeobox D12,Hoxd12)基因异常低表达,其差异表达倍数位列第一(图S6A、S6B);RT-qPCR和IHC结果证实,PPE组雌性子代大鼠出生前、后骨组织中Hoxd12表达持续降低(图3A−E)。出生后骨局部注射Hoxd12过表达慢病毒能显著逆转PPE所致雌性子代成骨功能及PBM降低(图3F、3G、S6C−J)。在孕期给予FMT-PPE组或PPE组母鼠补充DAI均可显著逆转长骨组织Hoxd12 mRNA及蛋白表达降低 (图3H−M)。综上提示,Hoxd12介导母体菌群代谢物DAI所致PPE雌性子代大鼠成骨功能及PBM降低。进一步,我们基于血DAI生理浓度,在长骨原代骨髓间充质干细胞(BMSCs)上证实DAI对Hoxd12表达及成骨功能的调控作用。与CON-BMSCs相比,PPE-BMSCs成骨分化功能与Hoxd12表达降低,而补充DAI可显著逆转PPE-BMSCs所致的上述改变,沉默Hoxd12则可显著逆转DAI所致的成骨分化功能增强(图3N−T、S7A−E)。综上提示,Hoxd12介导了DAI所致的雌性子代PPE-BMSCs成骨分化功能增强。

图3. Hoxd12介导PPE所致雌性子代大鼠成骨功能和PBM低下
(A、B)GD20和PW12子代Hoxd12表达;(C)HOXD12免疫组化图;(D、E)GD20和PW12雌性子代HOXD12表达量;(F)局部注射eGFP标记的HOXD12过表达慢病毒示意图;(G)micro-CT图;(H)FMT实验中雌性子代Hoxd12表达水平;(I)母鼠DAI补充实验中雌性子代Hoxd12表达水平;(J)FMT实验中雌性子代HOXD12免疫组化图;(K)母鼠DAI补充实验中雌性子代HOXD12免疫组化图;(L、M)PW12雌性子代HOXD12表达量分析;(N)CON-BMSCs和PPE-BMSCs中成骨分化标志基因表达水平;(O)CON-BMSCs和PPE-BMSCs的ALP染色图;(P)CON-BMSCs和PPE-BMSCs中Hoxd12表达水平;(Q−T)沉默Hoxd12对成骨分化标志基因表达的影响。
ERβ/KAT6A信号介导DAI诱导Hoxd12表达增加的表观遗传机制
已知孕期不良环境可通过表观遗传修饰异常诱导多器官发育编程改变。为了探究DAI逆转PPE所致Hoxd12低表达编程的表观遗传机制,我们检索表观遗传数据库(Cistrome Data Browser),发现Hoxd12最常见的表观遗传修饰形式为组蛋白修饰(图S8A)。胎长骨组织转录测序筛选结果提示,PPE组中组蛋白乙酰转移酶Kat6a变化最为明显,且RT-qPCR证实其表达降低(图4A、S8B),同时ChIP-PCR筛选证实出生前后Hoxd12启动子区H3K9ac水平均降低(图4B、4C)。在体外细胞水平,DAI能增加Kat6a表达和Hoxd12启动子区H3K9ac水平(图4D、4E),而沉默Kat6a可显著逆转DAI所致的Hoxd12启动子区H3K9ac水平及表达增加(图4F、4G、S8C−E)。综上提示,Kat6a介导DAI所致Hoxd12启动子区H3K9ac水平及表达增加。
已知DAI多通过雌激素受体β(estrogen receptor β,ERβ)发挥作用。为此,我们进一步证实DAI抑制Kat6a表达的ERβ调控机制。分子对接技术提示,DAI可直接结合于雌激素受体β(estrogen receptor β,ERβ)(图4H),Jaspar网站预测(https://jaspar.genereg.net)也提示,Hoxd12启动子区存在ERβ结合位点(图S8F)。我们在细胞水平发现,DAI对ERβ蛋白表达虽无明显影响但能促进其转位入核,且促进ERβ与Hoxd12启动子区结合(图4I−K、S8G),而沉默ERβ可显著逆转DAI所致的Kat6a mRNA表达、Hoxd12 mRNA及蛋白表达增加(图4L、4M、S8H−J)。综上提示,ERβ活化入核通过调控Kat6a表达介导DAI所致的Hoxd12组蛋白乙酰化水平及表达增加。
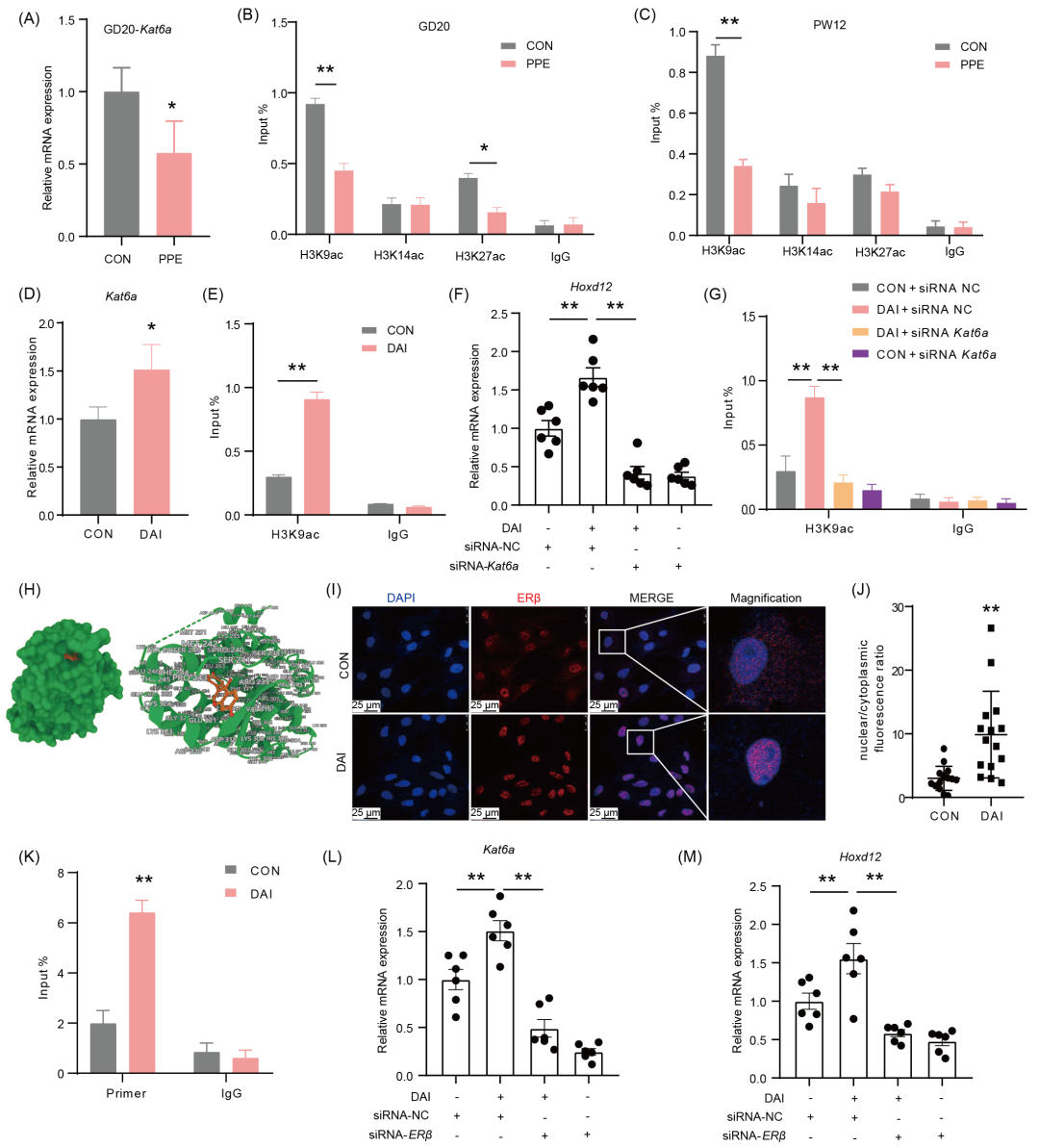
图4. DAI通过ERβ/KAT6A上调PPE-BMSCs中的Hoxd12
(A)雌性胎鼠Kat6a表达水平;(B、C)GD20和PW12子代Hoxd12启动子区H3K9、H3K14、H3K27ac水平;(D)经或未经DAI处理后BMSCs中Kat6a表达水平;(E)经或未经DAI处理后BMSCs中Hoxd12启动子区H3K9ac水平;(F)沉默Kat6a对Hoxd12表达的影响;(G)沉默Kat6a对Hoxd12启动子区H3K9ac水平的影响;(H)DAI(红色)和ERβ(绿色)之间分子对接图;(I、J)ERβ细胞免疫荧光图和核/质比分析;(K)DAI处理后PPE‐BMSCs中Hoxd12启动子区ERβ富集水平;(L)沉默Erβ对Kat6a表达的影响;(M)沉默Erβ对Hoxd12表达的影响。
讨 论
由于胎盘中存在能灭活糖皮质激素类药物活性的2型11β-羟类固醇脱氢酶,强的松通常仅有10%-20%会透过胎盘。因此,强的松被推荐用于治疗妊娠期间合并有类风湿性关节炎等自身免疫疾病孕妇的首选药物。虽然近期研究发现,PPE所致成年雌性子代OP易感与母体强的松龙暴露所致雌性子代骨骼肌线粒体自噬/FNDC5轴改变有关。然而,孕期使用强的松是否可以通过母体肠道菌群影响子代骨发育及OP易感,目前尚未见报道。在本研究中,临床研究发现,PPT组母体肠道菌群组成发生改变,伴有女性胎儿长骨发育抑制;动物实验发现,PPE可致母体大鼠肠道菌群组成改变,同时引起雌性子代大鼠出生前后长骨发育不良及成年PBM降低。提示PPT/PPE所致母体肠道菌群组成改变与子代长骨发育不良成年期OP易感关系密切。本研究进一步证实,PPT/PPE通过改变母体肠道菌群组成及降低其代谢物DAI生成,诱导雌性子代成骨Hoxd12低表达编程,导致长骨发育不良及OP易感,而孕期母体补充DAI是其有效防治策略。
PPE导致雌性子代长骨发育不良、PBM降低及OP易感
已知临床强的松使用通常为孕全程,剂量为1-60 mg/d,且临床指南推荐,孕期应以最低有效剂量使用强的松以控制母体疾病活动。PBM是指骨骼发育成熟时的骨量,是OP发生、发展的决定性因素。流行病学研究表明,PBM增加10%,预计OP的发生时间将推迟13年,重要的是OP相关的骨折风险亦明显降低。大鼠PW12(青春末期)和PW28(成年期)是观察评估PBM的两个常用时间点。研究表明,女性人群尤其是绝经后女性人群OP发病率和严重程度更高。在本研究中,我们基于临床实际和大鼠-人类用药剂量换算系数(6.17),孕全程经口灌胃给予大鼠0.25 mg/kg.d强的松,观察了PPE对雌性子代大鼠出生前后长骨发育、PBM及OP易感性的影响。我们发现,与临床研究一致,PPE组雌性(而非雄性)胎鼠股骨全长和POC长度降低,POC区域成骨矿化程度下降,且成骨功能下降。进一步发现,PPE组雌性(而非雄性)成年子代大鼠PBM下降。提示,PPE可导致雌性子代长骨发育不良、出生后PBM降低及OP易感。
母体菌群代谢物DAI介导PPE所致子代PBM降低及OP易感
母体肠道菌群作为一种可遗传的环境因素,是影响子代骨发育及PBM变化的另一重要因素。Abdul等利用高PBM及低PBM两种孕鼠模型,通过肠道菌群移植实验,发现母体肠道菌群能直接影响子代PBM。本研究发现,PPT/PPE会导致母体肠道菌群组成改变,其中以肠乳杆菌等有益菌丰度显著降低,进一步的肠道菌群移植实验证实,母体肠道菌群组成改变介导了PPE所致雌性子代成骨功能及PBM低下。在本研究中,我们发现PPT/PPE所致的孕妇与孕鼠肠道菌群组成改变并不完全一致。研究报道,妊娠晚期孕妇体内艾克曼菌、双歧杆菌和厚壁菌门等丰度显著增加,而随着孕龄增加,孕鼠则表现为厚壁菌门和疣状菌门减少。提示,孕期母体肠道菌群会随着孕龄增加而发生适应性改变,且妊娠期孕妇和孕鼠肠道菌群在组成上存在差异。此外,人类的生活方式(如饮食、运动、压力等)对肠道菌群的组成和功能有重要影响,然而实验中大鼠的饲养条件相对单一,缺乏人类生活中的复杂性。
研究表明,肠道菌群代谢物是母体肠道菌群与胎儿发育之间的重要“桥梁”。代谢组学与表型改变密切相关,是研究孕期不良环境影响胎儿发育机制的高效技术手段。本研究通过PPE/PPT血标本非靶向和靶向代谢组分析发现,母血与雌性胎血代谢谱发生显著改变,其中DAI水平降低(表S3)。DAI是一种能通过胎盘并影响胎儿发育的游离型异黄酮,广泛存在于母-胎儿组织中(如母体尿液、血液、羊水、母乳以及脐血),且其水平受到肠道菌群的影响。在一项纳入480对母-婴作为研究对象的前瞻性队列研究中发现,“经常”食用豆制品的孕妇尿液中DAI等异黄酮浓度比对照组升高,同时还发现孕妇豆制品食用频率与新生儿出生体重呈正相关,且母体尿液中染料木黄酮、DAI等异黄酮成分的浓度与新生儿出生体重、手臂和腰围以及三头肌和腹部皮褶厚度增加呈正相关。动物实验也发现,孕期低水平摄入异黄酮会增加子代出生后二乙基亚硝胺诱导的肝癌发生风险。研究表明,组织中DAI水平主要取决于肠道菌群(如肠乳杆菌)中β-GC或β-GAL水解食物中结合型异黄酮的能力。在本研究中,我们基于多组学分析发现PPE母体肠道中肠乳杆菌相对丰度与胎骨组织DAI水平呈正相关,基于PICRUSt功能预测提示PPE可致相关代谢通路发生改变,采用酶活性检测证实PPE组母体肠道菌群β-GC活性显著降低。提示,PPE所致母-胎血中DAI水平的下降可能与母体肠道菌群中肠乳杆菌β-GC活性降低有关。进一步,给予FMT-PPE组或PPE组母鼠补充DAI均可显著逆转PPE所致雌性成年子代长骨组织DAI水平、成骨功能及PBM的降低。综上提示,母体菌群代谢物DAI介导了PPE所致子代PBM降低及OP易感,有望成为防治胎源性OP易感的有效干预靶标。
DAI通过ERβ/Kat6a表观遗传调控Hoxd12表达介导了PPE子代成骨功能降低及OP易感
进一步,我们探究了胎血低DAI水平影响子代长骨发育的表观遗传编程机制。已有研究表明,敲除Hoxd基因会导致Runx2表达及成骨功能抑制;Hoxd12基因敲除小鼠出现骨骼发育障碍、骨量降低。在本研究中,我们通过转录组测序发现胎鼠长骨组织中Hoxd12基因表达降低,同时证实PPE雌性子代出生前、后骨组织Hoxd12表达持续降低。进一步,在PW8骨局部过表达Hoxd12能逆转PPE雌性成年子代成骨功能及PBM下降,而孕期母体补充DAI或给予原代BMSCs补充DAI能逆转PPE所致的子代Hoxd12表达、成骨功能及PBM低下。这些研究结果均提示,出生前、后Hoxd12低表达编程改变介导PPE所致的雌性子代长骨发育不良及PBM低下,Hoxd12是胎源性OP易感的出生后干预靶标。
已知DAI属于植物雌激素,可作为选择性ER激动剂而调控下游基因表达。ER包括ERα和ERβ亚型,其中ERα主要表达于皮质骨中,ERβ主要表达于松质骨中,且在BMSCs成骨分化过程中ERβ作用更为重要,因此本研究聚焦于ERβ。孕期不良环境暴露可通过表观遗传机制增加子代疾病易感性。Cistrome Data Browser提示组蛋白修饰是Hoxd12最常见的表观遗传修饰形式,骨转录组测序并验证发现组蛋白乙酰转移酶Kat6a表达降低,同时ChIP-PCR证实出生前后Hoxd12启动子区H3K9ac水平降低。在细胞水平,DAI能增加Kat6a表达、Hoxd12启动子区H3K9ac水平及表达,而沉默Kat6a可显著逆转DAI所致的上述效应。进一步发现,DAI可活化ERβ并促进ERβ与Hoxd12启动子区结合,而沉默ERβ可显著逆转DAI所致的Kat6a和Hoxd12表达增加。综上提示,DAI一方面通过活化ERβ入核并与Hoxd12启动子区结合,以促进其转录表达;另一方面通过活化Erβ上调Kat6a表达,促进Hoxd12组蛋白乙酰化水平及表达增加(图5)。
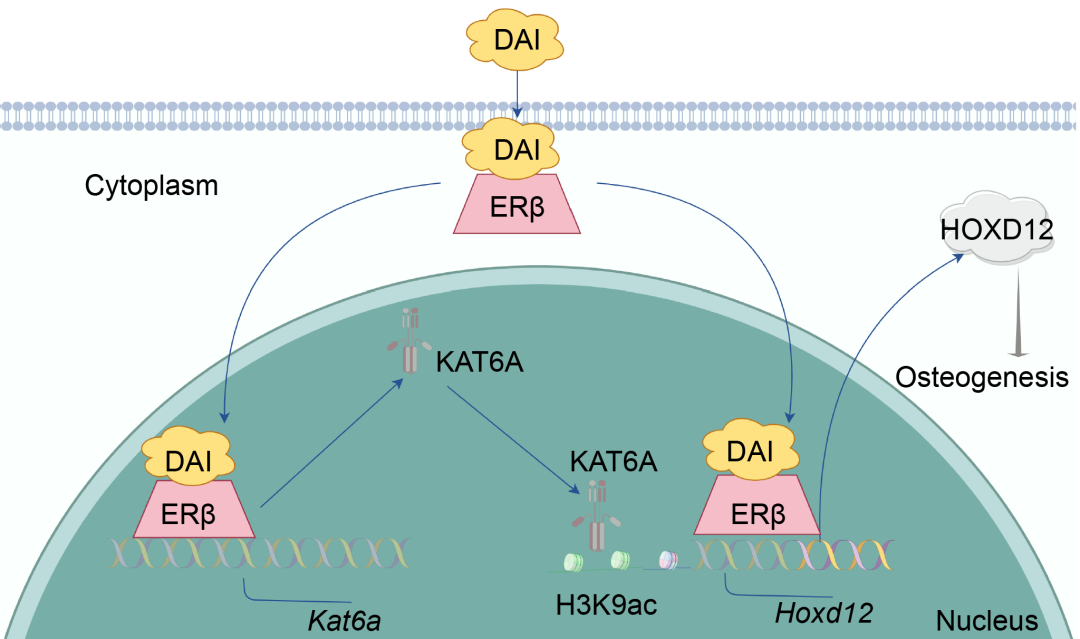
图5. DAI通过ERβ/KAT6A表观遗传调控Hoxd12并促进PPE‐BMSCs成骨分化功能
DAI通过促进ERβ核转位上调Kat6a表达,增加Hoxd12启动子区H3K9ac水平,从而提升其转录活性。同时,DAI通过促进ERβ核转位并直接结合于Hoxd12启动子区,从而促进Hoxd12转录表达,最终增加BMSCs的成骨分化能力。
此外,考虑体内雌激素也是影响宫内胎儿发育的关键因素,并且具有活化ERβ作用。为此,我们进一步检测GD20血雌激素水平发现,PPE对雌性胎鼠血清雌激素水平无显著影响,且强的松或强的松龙对BMSCs中Hoxd12表达均无显著影响(图S9)。提示,PPE所致雌性子代Hoxd12表达及PBM降低与胎血中雌激素、强的松和强的松龙暴露无显著关联。
菌群产物DAI是PPE所致子代多器官功能稳态改变及相关疾病易感的早期共性防治靶标
近期研究提示,孕期外源物(如药物)暴露可通过多途径、多器官及多靶点影响胎儿形态与功能发育。既往研究表明,孕期药物暴露可导致子代发育毒性和多器官功能异常。而且不同器官对同一药物的敏感性也不同。本研究也发现,PPE导致雌性子代出生后成骨功能及峰值骨量下降的同时,肝脏脂质合成增加、海马神经发育受损、肾上腺甾体合成功能增加、卵巢卵泡发育不良等多器官功能改变(图S10A−G)。提示,PPE导致子代多器官(包括长骨、肝脏、海马、肾上腺和卵巢等)功能稳态改变。同时,我们发现女婴血清中DAI含量与其出生体重呈正相关(图S10H)。本研究进一步发现,母体补充DAI能逆转PPE所致长骨PBM下降的同时,肝脏脂质合成增加、海马神经发育受损、肾上腺甾体合成功能增加、卵巢卵泡发育不良等多器官功能改变出现逆转效应。提示,母体补充DAI能有效促进宫内胎儿整体发育,并能预防子代出生后疾病易感。孕期补充DAI是防治PPE相关多种胎源性疾病易感的共性靶标。本研究结果也提示,对于临床合并有自身免疫性疾病的孕妇在使用强的松治疗的同时,可考虑每天摄入富含异黄酮食品(大豆制品或普通豆腐),以预防子代出生后多器官功能稳态改变。
本研究存在的局限性或不足之处
本研究中存在一定的局限性或不足之处。第一,由于胎源性疾病具有发病时间长,本研究仅通过回顾性病例-对照研究评估PPT对宫内胎儿长骨发育的影响,未探究PPT对子代出生后骨量累积的影响,未来需通过妇产科、营养科、骨科等多学科协作建立多中心临床队列研究进一步确证PPT与子代出生后骨量累积的关系。第二,本研究基于体内、外实验提出母体补充DAI是防治子代多器官稳态改变的有效策略,仍有待大样本量的临床研究进一步确证其安全性及有效性。
结 论
本研究首先通过临床和整体动物实验证实,PPT/PPE通过改变母体肠道菌群(如乳杆菌)组成并抑制β-GC活性,降低母血菌群代谢物DAI水平;而胎血低水平DAI通过诱导Hoxd12低功能编程,导致雌性子代成骨细胞分化抑制、长骨发育不良及成年PBM低下。进一步,在细胞水平证实,DAI是通过活化ERβ上调Kat6a,继而增加Hoxd12启动子区H3K9ac水平、表达及BMSC成骨分化。最后,在整体动物水平证实,孕期母体补充DAI能逆转PPE子代成骨功能、PBM低下及OP易感,且能逆转肝脏、海马、肾上腺和卵巢等其他器官发育损伤性改变。本研究提出肠道菌群−表观遗传−骨发育轴新观点,为OP防治提供了新的靶点。本研究创新性提出DAI是非常具有转化潜力的胎源性OP防治靶点,且孕期补充菌群代谢物可能是防治孕期糖皮质激素暴露所致胎儿发育编程改变的有效策略,提示优化孕期母体环境的重要性。
方 法
化学品和试剂
强的松(LA21334,486-66-8,纯度 ≥ 98%,Xianju Pharmaceutical Co.)。DAI(486-66-8, 纯度 ≥ 98%,RHAWN®)。异氟烷(R510–22–10,Baxter Healthcare Co.)。Taq Pro Universal SYBR qPCR Master Mix(Q712–02)以及HiScript III RT SuperMix(R323–01)购自Vazyme Biotech Co.。Lipofectamine®3000试剂(L3000015)、TRIzol TM试剂(15596026)、α-MEM培养基(12571063)、磷酸盐缓冲液(10010023)和胎牛血清(A5670402)购自Thermo Fisher Scientific Co.公司。Omni-ECL™Femto Light化学发光试剂盒(SQ101L)购自Epizyme Biotech公司。Erβ siRNA、Kat6a siRNA和Hoxd12 siRNA由Tsingke Biotech Co.合成。其他化学试剂均为分析级。HOXD12抗体(Ab230697)购自Abcam Co.公司。RUNX2(A11753)、COL1A1(A24112)、ERβ(A26153)、H3K9ac抗体(A7255)、H3K14ac(A7254)、H3K27ac(A7253)、IgG以及GAPDH(AC002)抗体均购自武汉Abclonal公司。
临床研究
本研究获得武汉大学中南医院医学伦理委员会批准(伦理号:临研伦[2023141K])。在病例-对照研究中,我们选取2020至2023年就诊于武汉大学中南医院的孕产妇病历,获取既往用药史、新生儿性别及出生体重、产前B超等数据。纳入标准:20-35岁育龄期产妇;单胎妊娠;定期于中南医院门诊完成产检且能获取到孕晚期产检B超结果。排除标准:多胎妊娠;妊娠期合并糖尿病、高血压等疾病;产前诊断已确诊合并胎儿畸形或其他疾病;孕期有地塞米松、倍他米松、氢化可的松等其他糖皮质类激素用药史。本研究共获取到223对产妇-新生儿病例数据(表S1),根据病历记录中既往有无孕期强的松用药分为:CON组与PPT组,PPT组孕妇均有口服2-10 mg/d强的松用药史。考虑到胎龄是影响宫内胎儿生长发育指标的主要影响因素,依据参考文献计算生长指数Z值和FL/BPD比值以匹配胎龄这一混杂因素。
在孕妇母体粪便收集中,在武汉大学中南医院和武汉市中医院产科门诊招募25例孕妇(表S2),分组、纳入与排出标准与病例-对照研究基本相同,但要求收集粪便标本二周前无抗生素使用史。以签署纸质知情同意书的方式取得患者或家属知情同意,收集母体粪便于含保护液的无菌采样管(MGI Tech Co.,Ltd,A0212),收集后24 h内转存于−80 ℃冰箱,用于后续16s rRNA测序分析。
在临床血标本收集中,在武汉大学中南医院产科招募41对母体-女婴(表S3),分组、纳入与排出标准与病例-对照研究基本相同,以签署纸质知情同意书的方式取得家属知情同意,收集母血和女婴脐带血,样本在–80 ℃下保存,用于后续定量检测DAI水平。分组、纳入与排出标准与病例-对照研究基本相同,以签署纸质知情同意书的方式取得患者或家属知情同意,收集母体血液及女婴脐带血,并保存于−80 ℃冰箱,用于后续DAI水平定量检测。
动物与处理
SPF级雌性Wistar大鼠(240±10 g)和雄性Wistar大鼠(280±20 g)(No. 110324210100788048)购买于北京SpePharm公司。本研究中的设计方案与动物处理流程均符合相关伦理要求,且所有动物实验方案均由武汉大学动物实验中心实验动物管理与使用委员会(IACUC)批准(批准号:NO. WP20210060),并按照美国国立卫生研究院(NIH)《实验动物护理和用法指南》和《体内实验动物研究报告指南》(ARRIVE)进行实验。所有Wistar大鼠均在标准条件下饲养: 12 h光照/黑暗周期,温度控制在23±2 ℃,相对湿度为40%−60%。经适应性喂养1周后,将大鼠按2雌+1雄进行合笼交配,次日于镜下在阴道涂片中发现精子确定为受孕大鼠,记为GD0。PPE组母鼠于GD0−20每日给予灌胃0.25 mg/kg强的松,而对照组母鼠于GD0−20给予等剂量溶媒(0.5%羧甲基纤维素钠)灌胃。
在GD20时,随机选取部分对照组和PPE组孕鼠,给予吸入异氟烷麻醉后处死收集胎鼠。本研究仅纳入每窝胎鼠数在8−14只的孕鼠。收集每窝胎鼠股骨组织标本进行分析。其余对照组和PPE组孕鼠自然分娩,为保证仔鼠哺乳期每窝营养均衡,于出生后1 d至PW4将每窝仔鼠调整为12只(雌鼠及雄鼠数量相等)。断奶后,每窝随机选择两只雌性和两只雄鼠,分别在PW12和PW28取材。此外,每窝选出一只雌鼠于PW8和PW9进行股骨远端和胫骨近端Hoxd12过表达慢病毒或阴性对照慢病毒注射(10 μL,2.91×108 TU/mL),慢病毒注射4周后收集股骨标本检测分析。实验示意图1E所示。
肠道菌群移植(FMT)
FMT实验中使用的伪无菌鼠前期文献报道中的方法构建:给予6−8周龄SPF级雌性大鼠连续5 d灌胃广谱抗生素混合溶液(万古霉素:100 mg/kg;硫酸新霉素、甲硝唑和氨苄青霉素均为:200 mg/kg)。考虑到强的松低脂溶性和低肠道滞留特性且其半衰期约为3−4 h,本研究为确保肠道内容物中无强的松残留,GD20母体粪便均在给予母鼠强的松后12 h后收集。收集抗生素应用前后大鼠粪便标本,分别在厌氧和有氧条件下进行琼脂平板培养以观察伪无菌鼠构建是否成功。将构建的伪无菌Wistar大鼠与雄鼠进行交配以获得伪无菌孕鼠,并参照前期文献报道方法于GD0−5进行FMT。即,用无菌粪便采集器分别收集对照组和PPE组GD20母鼠新鲜粪便样本;在洁净工作台中,将新鲜粪便样品重新悬浮在无菌 PBS(7.5 mL/g 粪便)中,并剧烈涡旋5 min,然后600 g离心3 min;收集上清,连续5 d给予伪无菌孕鼠灌胃上述制备的上清。
骨组织形态学分析
将收集的股骨标本固定于4%多聚甲醛中。对于成年大鼠,股骨于10% EDTA液中进行脱钙12 d。将胎鼠股骨标本及脱钙后的成年大鼠股骨标本进行石蜡包埋,制作厚度约4 μm矢状面切片。H&E、Von Kossa和TRAP染色方法及骨组织形态学分析如前面文献报道所述。使用Olympus AH‐2光学显微镜进行拍照,并使用Image-Pro Plus软件(6.0版)对每个样品的五个随机区域进行结果分析。
免疫组织化学
骨组织样本依次进行脱水、石蜡包埋并制备4 μm切片。脱蜡至水后,于pH 6.0柠檬酸抗原修复液进行抗原修复。PBS洗3次后,将切片置于3%双氧水溶液中孵育20 min,接着用3% BSA封闭1h。随后切片滴加一抗并于4 ℃湿盒中孵育24 h,然后加入HRP标记的二抗孵育1 h。PBS洗涤三次后,加入新配制的DAB溶液(Servicebio)显色,阳性表现为棕黄色。用苏木精复染后,用进行拍照,并使用Image-Pro Plus软件(6.0版)对每个样品的五个随机区域进行结果分析。
免疫荧光染色
组织切片使用pH 6.0柠檬酸抗原修复液进行抗原修复。细胞样本在室温下用4%多聚甲醛固定15 min。骨组织切片和细胞样本均使用0.1% Triton X-100进行通透,并于室温下使用3% BSA封闭60 min。用PBS洗三次后,将样品在4 ℃分别与一抗(HOXD12(1:100)、ERβ(1:100)和RUNX2(1:100))孵育过夜。次日,PBS洗3次后,将样品与CY3标记的荧光二抗(1:200)在室温下孵育1 h。使用Leica TCS SP8 X激光共聚焦显微镜或尼康H550S照片成像系统(日本)拍照并对每个样品的五个随机区域使用Image-Pro Plus软件(6.0版)进行结果分析。
微型计算机断层扫描(micro-CT)
使用高分辨率微型CT(Skyscan1276,Bruker Biospin)扫描股骨样本,参数如下:source voltage 50 kV,source current 200 μA,image pixel size 20 μm以及 energy filter of 1.0。ROI区域为股骨远端骨骺生长板开始的200张CT影像图。使用CTAn软件(版本1.18.4)进行骨量分析,参数包括BV/TV、Tb.N、 Tb.Th和Tb.Sp。
16s rRNA肠道菌群测序
使用CTAB或SDS方法提取样品中基因组DNA,并用1%琼脂糖凝胶检测基因组DNA的纯度和浓度。扩增16S rRNA基因的V3-4区域,引物序列如下:F:5’-CCTAYGGGRBGCASCAG-3’,R:5’-GGACTACNNGGGTATCTAAT-3’。定量后,使用NEBNext® Ultra™ IIDNA文库制备试剂盒(Cat No. E7645,MA,USA)生成测序文库。文库质检合格后,在Illumina NovaSeq 6000平台完成测序。
非靶向代谢组学
取100 μL血清标本于EP管中,加入80%甲醇400 μL。涡旋振荡混匀后,置于冰上孵育5 min。在4 ℃以15000 g离心20 min。保留上清,并转移到新的EP管中。加入质谱级水,使甲醇浓度稀释至53%。再次在4 ℃下以15000 g离心20 min,收集上清液并注入LC-MS进行分析。LC-MS/MS分析由Vanquish超高效液相色谱系统(德国赛默飞世尔公司)和Orbitrap Q ExactiveTM HF质谱仪(Thermo Fisher)共同完成。
在实验过程中,通过混合等体积的实验样品制备七个质量控制(QC)样品,用于平衡色谱-质谱系统、监测仪器状态以及评估整个实验过程中的系统稳定性,同时还设置了一个空白样品以去除背景离子。血清样品被注入Hypesil Gold色谱柱(C18)。正极性模式的洗脱液为洗脱液A(0.1% FA 水溶液)和洗脱液B(甲醇)。负极性模式的洗脱液为洗脱液A(5 mM乙酸铵,pH 9.0)和洗脱液B(甲醇)。对原始数据使用Compound Discoverer 3.1(CD3.1,Thermo Fisher)进行处理,以获得代谢物的定性和定量结果,并对数据进行质量控制,以确保数据结果的准确性和可靠性。对代谢物进行统计分析,包括主成分分析和OPLS-DA模型中第一主成分的变量投影重要度(variable importance in the projection,VIP)以评估各组之间的代谢差异。VIP>1且倍数变化>2的代谢物被认为发生了显著变化。
液相色谱/串联质谱(LC-MS/MS)
参照文献报道所述进行DAI水平定量检测。简而言之,使用LC-MS/MS系统(TSQ Quantis,Thermo Scientific),参数设置如下:色谱柱:C18 柱(4.6 mm,150 mm,5 μm);柱温:30 ℃;流动相:乙腈和 1%磷酸(30:70),流速:1 mL/ min。
转录组测序和分析
对照组和PPE组胎鼠骨组织转录组测序在Novogene Biology Information Technology Co.,Ltd使用Illumina NovaSeq平台完成。使用DESeq2软件(v1.20.0)进行差异表达分析,差异表达基因定义为校正p值<0.05。通过上一步获得差异表达基因后,使用ClusterProfiler软件(v3.4.4)进行GO和KEGG富集分析。
原代BMSCs的提取、培养和处理
原代BMSCs的提取、培养及成骨分化依据前期报道方法进行。简而言之,显露股骨和胫骨后去除骨组织周围肌肉等软组织,并用PBS冲洗3次。去除干骺端两端骨骺,暴露骨髓腔后用α-MEM培养基冲洗骨髓腔。收集冲洗液,在室温下以1000 r离心3 min,,弃上清。再次用PBS重悬、离心2次后,用完全培养基(α-MEM培养基+10%胎牛血清+1%青霉素/链霉素双抗)进行重悬,置入37 ℃、5% CO2 培养箱中培养。对于成骨分化,将BMSCs接种于12孔板并培养贴壁至60%−80%时,更换为成骨分化诱导培养基(α-MEM培养基+10%胎牛血清+1%青霉素/链霉素双抗+10 nM 地塞米松+ 50 μg/l VitC+10 mM β-甘油磷酸钠)继续培养2周。成骨诱导分化过程中,分别使用DAI、强的松和强的松龙等进行处理。
siRNA沉默Kat6a、Hoxd12和Erβ表达
为沉默Hoxd12、Erβ以及Kat6a基因表达,我们设计了siRNA转染实验。将BMSCs接种于6孔板并培养贴壁至60%−80%时,用Lipofectamine 3000进行转染6 h,采用RT-qPCR验证沉默效率。由北京擎科生物技术有限公司设计合成Kat6a、Hoxd12和Erβ等基因siRNA,为保证沉默效率各基因均设计合成了三条siRNA序列,靶向序列如下:Erβ-1:F:5’-CACGAATCAGTGTACCATA-3’;Erβ-2:F:5’-GTCCTGCTGTGATGAACTA-3;Erβ-3:F:5’-CACCTTGAGTCCAGAGCAA-3′;Kat6a-1:F:5’-GCGCTATGCTAATCCAATA-3’;Kat6a-2:F:5’-CTTCCACACGAGAAAGATA-3’;Kat6a-3:F:5’-GGCGAATAGCACTTCCTAA-3’;Hoxd12-1:F:5’-GGTTCACTCGGCTCTCAAA-3’;Hoxd12-2:F:5’-GACCAGGTCAAGTTCTATA-3’;Hoxd12-3:F:5’-CTTCAAGGAAGACACCAAA-3’。
茜素红S(ARS)染色
为检测BMSCs成骨分化能力,BMSCs成骨诱导分化2-3周后进行ARS染色。简而言之,用PBS洗涤细胞并用固定液在室温下固定细胞样品20 min。再次用PBS洗涤后,用ARS溶液在室温染色30 min。用蒸馏水轻柔飘洗,采用尼康显微镜(ECLIPSE TE2000‐E,Japan)进行拍照,阳性表现为红色矿化结节。
碱性磷酸酶(ALP)染色
诱发BMSCs成骨分化10 d后,使用BCIP/NBT ALP 显色试剂盒(MA0197,Meilunbo)进行ALP染色。用PBS洗涤细胞后,用4%多聚甲醇固定细胞样品5至10 min,再次用PBS洗涤3次。去除PBS进行细胞固定细胞后,每孔样品与400 μL ALP染色液于避光处孵育30 min。用PBS洗后,采用尼康显微镜(ECLIPSE TE2000‐E,Japan)进行拍照,阳性表现为深蓝色至蓝紫色。
RNA提取和RT-qPCR
根据TRIzol试剂盒(Invitrogen)说明书提取骨组织及BMSCs中RNA。简要步骤如下:往样品中加入1 mL TRIzol试剂和200 μL氯仿,剧烈震荡摇匀后,于冰上静置10 min。以12000 r离心15 min,往收集的上清液中加入400 μL异丙醇沉淀RNA。放于室温,静置10 min后,以12000 r再次离心10 min。弃上清后加入1 mL预冷的75%乙醇洗涤RNA两次。使用紫外分光光度计(NanoDrop One C)进行RNA浓度与纯度测量。取1 μg RNA使用HiScriptⅡ 1st Strand cDNA Synthesis Kit (R212‐01; VAZYME)进行逆转录合成以DNA。在QuantStudio 6 Flex实时PCR系统(Thermo Scientific)上使用Taq Pro Universal SYBR qPCR Master Mix(Q712-02,VAZYME)进行RT-PCR。PCR条件如下:95 ℃ 预变性30 s,40个循环反应(95 ℃ 10 s、60 ℃ 30 s)。以Gapdh为内参基因,根据Y=2^(-ΔΔct)公式进行计算相对表达水平。
Western blot实验
用预冷PBS洗涤细胞样品三次后,使用含有1 mM PMSF的RIPA裂解液(Beyotime Biotechnology,中国上海)进行裂解。于冰上静置20 min,在4 ℃下以12000 r离心10 min,保留上清即为提取的总蛋白。使用BCA试剂盒(ZJ101,Epizyme Biotech)检测蛋白质浓度。往10% SDS-PAGE胶样品孔中加入30 μg蛋白样品,电泳后进行转膜。用无蛋白快速封闭液中(PS108P,Epizyme Biotech)封闭5 min后,加入一抗(ERβ(1:1000)和GAPDH(1:2000))并在4 ℃孵育过液。用TBST洗膜后,加入HRP标记的二抗(1:8000)室温孵育1 h,使用Omni‐ECL™Pico Light Chemiluminescence Kit(SQ202,Epizyme Biotech)进行显影,条带结果采用Image Pro Plus软件进行统计分析。
分子对接分析
为了评估大豆苷元与ERβ之间的结合能和相互作用模式,我们使用了计算机蛋白-配体对接软件AutodockVina 1.2.2。首先,从PubChem化合物数据库(https://pubchem.ncbi.nlm.nih.gov)中获得大豆苷元的分子结构(PubChem CID 5281708),从PDB(http://www.rcsb.org/)下载ERβ蛋白(PDB编号,1QKN,Resolution:2.25 Å)的3D坐标。接着,将所有蛋白质和分子文件都转换为PDBQT格式,去除了所有水分子,并添加了极性氢原子。网格框居中以覆盖每个蛋白质的结构域并适应自由分子运动。对接口袋设置为一个30Å×30Å×30Å的正方形口袋,格点距离为0.05 nm。分子对接研究由Autodock Vina 1.2.2(http://autodock.scripps.edu/)用于模型可视化。
染色质免疫沉淀-qPCR(ChIP-PCR)
ChIP-PCR方法参照前期文献报道所述。简要步骤如下:将匀浆后骨组织和BMSCs样品与1%甲醛交联10 min,接着在室温下加入0.125 M甘氨酸终止交联5 min。以3000 r离心2 min后,弃上清,置入超声破碎仪(功率30%,持续运行2 s后暂停1 s,共5 min)以获得约200 bp的DNA片段。在4 ℃下,以12000 r离心10 min,保留上清,并将10 μL上清液保留一份为“Input”,保存于4 ℃备用。剩余上清在4 ℃下与Protein G Beads和一抗(IgG、H3K4ac、H3K9ac、H3K27ac)于旋转摇床上(20-30 r)孵育过夜。收集Beads,用Elution buffer(200 μg/mL蛋白酶K)进行洗脱及重悬。继续将获得的标本转入65 ℃恒温水浴中进行解交联过夜,按照PCR Clean Kit(Beyotime Biotechnology,中国上海)操作步骤进行DNA纯化,针对纯化后的DNA启动子区进行RT-qPCR。Hoxd12启动子区域核苷酸序列引物如下:F:5’-CTGCCAGACCTTCTCCAGTC-3’;R:5’-TTGGGAGTTTTCCTGGTCTG-3’。用于检测ERβ和Hoxd12启动子区结合的引物序列如下:F:5’-TGGTCGCCATAACTCACACA-3’,R:5’-AGAGACTGTGTGATGGGAGC-3’。
数据统计分析
使用SPSS 20和GraphPad Prism 8.0软件进行数据统计分析。除非另有说明,数据均以平均值±标准误,当数据满足正态分布且方差齐时,两组比较使用Student’s t检验,多组间采用方差分析并用Dunnett或Tukey进行组间比较。当数据不满足正态分布或方差不齐时,采用Nemenyi及Kruskal-Wallis H检验。p值小于0.05被认为存在显著差异。
代码和数据可用性:
本研究结果所使用的原始测序数据已经上传至NCBI序列读取档案(SRA)(https://www.ncbi.nlm.nih.gov/bioproject)中:编号PRJNA1241687(https://www.ncbi.nlm.nih.gov/sra/PRJNA1241687);PRJNA1243860(https://www.ncbi.nlm.nih.gov/sra/PRJNA1243860);PRJNA1241537(https://www.ncbi.nlm.nih.gov/sra/PRJNA1241537)。在Novomagic(https://magic.novogene.com)及metware(https://cloud.metware.cn)云平台进行RNA测序、16s rRNA肠道菌群测序和非靶向代谢组学数据分析。主要数据和代码已上传至Github网站,网址为:https://github.com/machihn/ChiMa-iMeta.git。补充材料(文本、图、表、中文翻译版本或视频)也可从线上DOI或iMeta Science(http://www.imeta.science/)上获取。
引文格式:
Chi Ma, Hangyuan He, Kunpeng Wang, Juanjuan Guo, Liang Liu, Yuting Chen, Bin Li, et al. 2025. Maternal Gut Microbiota‐Derived Daidzein Prevents Osteoporosis in Female Offspring Following Prenatal Prednisone Exposure. iMeta 4: e70037. https://doi.org/10.1002/imt2.70037.
作者简介

马翅(第一作者)
● 博士,2024年6月毕业于武汉大学中南医院。
● 研究方向为骨与关节疾病的胎儿发育起源,以第一作者在iMeta、Toxicology Letter等期刊发表SCI论文5篇。

何航元(第一作者)
● 武汉大学第二临床学院在读博士研究生。
● 研究方向为骨与关节疾病的胎儿发育起源。以共同第一作者在iMeta、Faseb Journal、Life Sciences等期刊发表SCI论文8篇。

王鹍鹏(第一作者)
● 硕士,2012年6月毕业于湖北中医药大学。
● 研究方向卵巢储备下降,自身免疫与生殖相关疾病。以第一作者或共同第一作者在iMeta、《中国病理生理杂志》《中医药导报》等期刊发表论文。参与国家十三五科技部重大专项“针灸对卵巢功能影响的疗效评价研究”。

汪晖(通讯作者)
● 武汉大学基础医学院药理学二级教授、珞珈杰出学者、博士生导师。兼发育源性疾病湖北省重点实验室主任、武汉大学中南医院妇儿医院特聘教授。
● 研究方向为外源物发育毒理、胎源性疾病防治策略。主持国家重点研发计划重点专项、国家自然科学基金重点/重点国合/联合重点项目5项、教育部科学研究重大项目等。在iMeta、Cell Reports Medicine、Advanced Science、Acta Pharmaceutica Sinica B、Journal of Advanced Research、Metabolism、BMC Medicine、Cell Death & Disease等国际权威杂志上发表发育毒理学相关系列论文和综述300余篇,授权专利17项,出版主编《FETAL ORIGIN OF DISEASE》《胎源性疾病》等中英文学术专著4部。科研成果获全国妇幼健康自然科学一等奖、湖北省科技进步一等奖、中国出生缺陷干预救助基金会科学技术二等奖等。

陈廖斌(通讯作者)
● 武汉大学中南医院骨科教研室主任、关节与运动医学科主任、武汉大学关节疾病研究中心主任,二级教授,博士生导师。
● 研究方向为骨与关节疾病的胎儿起源;关节疾病的临床诊治。国务院政府特殊津贴专家。主持国家自然科学基金重点项目1项、面上项目5项;主持省部级重大、重点等其它项目10余项。以第一及通讯作者在iMeta、Advanced Science、Journal of Advanced Research、Chemical Engineering Journal、Carbohydrate Polymers、Bone and Joint Journal、Arthroscopy、American Journal of Sports Medicine等高影响力期刊发表SCI论文159篇;主编胎源性骨与关节疾病专著2部,关节疾病临床专著2部。
更多推荐
(▼ 点击跳转)
iMeta | 引用16000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 兰大张东组:使用PhyloSuite进行分子系统发育及系统发育树的统计分析
iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVI
iMeta封面

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

3卷2期

3卷3期

3卷4期

3卷5期

3卷6期

4卷1期

4卷2期
iMetaOmics封面

1卷1期

1卷2期

2卷1期
期刊简介
“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述,重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊!相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录!2024年6月获得首个影响因子23.8,中科院分区生物学1区Top,位列全球SCI期刊前千分之五(107/21848),微生物学科2/161,仅低于Nature Reviews,学科研究类期刊全球第一,中国大陆11/514!
“iMetaOmics” 是“iMeta” 子刊,主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任,是定位IF>10的高水平综合期刊,欢迎投稿!
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science

















 1523
1523

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









