





 文章探讨了小脑在社会认知中的功能,特别是其在理解他人心理状态、预测社会行为和处理社会互动中的作用。研究表明,小脑不仅涉及运动控制,还在社会理解他人心理和行动序列预测中发挥重要作用。小脑的特定区域,如CrusI和II,与社会心理化过程密切相关。此外,小脑在社会学习和适应性行为中通过预测和错误修正机制发挥作用,这在自闭症和小脑疾病患者中可能会受损。
文章探讨了小脑在社会认知中的功能,特别是其在理解他人心理状态、预测社会行为和处理社会互动中的作用。研究表明,小脑不仅涉及运动控制,还在社会理解他人心理和行动序列预测中发挥重要作用。小脑的特定区域,如CrusI和II,与社会心理化过程密切相关。此外,小脑在社会学习和适应性行为中通过预测和错误修正机制发挥作用,这在自闭症和小脑疾病患者中可能会受损。
传统观点认为小脑主要控制运动行为。虽然近期的研究已经揭示小脑也支持如认知和情感等非运动功能,但直到过去的5年,人们才明确地认识到小脑在社会角色中也扮演着重要的部分。这个角色在社会认知中非常明显,基于通过个体的运动来解释目标导向行为(社会“镜像”),这与小脑在运动学习中的原始角色非常接近,以及在理解其他个体的心理状态中的社会理解,如他们的意图、信念、过去的行为、未来的期望和个性特征(社会“理解他人的心理”)。这种理解他人心理的角色大多数是由后部小脑(例如,Crus I和II)支持的。最主要的假设是小脑协助学习和理解社会行动序列,从而通过支持对即将发生或未来的社会互动和合作的最优预测,促进社会认知。这篇共识论文集结了来自不同领域的专家,讨论了最近在理解小脑在社会认知中的角色,以及理解他人的社会行为和心理状态,其对小脑共济失调和自闭症谱系障碍等临床障碍的影响,以及小脑如何成为非侵入性脑刺激作为治疗干预的潜在目标等方面的努力。我们报告了最新的实证研究结果和理解和操控人类小脑回路的技术。小脑回路现在看起来是阐明社会互动的关键结构。本文发表在the Cerebellum杂志。
引言
关于小脑与社会认知的关系的研究还处于早期阶段,除了偶尔的早期贡献外,这方面的研究在过去的5年中开始浮现。先前关于小脑社会角色的报告通常仅限于情感处理的侧面方面,并且以轶事形式描述小脑病患者有情感缺陷。这些报告侧重于理解他人面部表情中的情感[1],而对他人的高级心理状态的关注并不多。然而,来自社会神经科学领域(Frank Van Overwalle)和小脑领域(Peter Marien和Mario Manto)的研究者之间的新颖合作,发现了小脑的重要社会功能[2,3],这激发了新的研究关于小脑在社会认知中的潜在角色。社会认知过程包括依赖于推断出的其他人未观察到的心理状态的社会“理解他人的心理”(或读心)以及依赖于观察到的他人目标导向的身体运动的社会“镜像”(或读身)。大脑的研究已经证明,这两个过程招募了不同的皮质区域。
社会理解他人心理是一种进化较新的功能,它激活了负责将自己的视角切换到另一个人不可观察的心理状态(例如,意图,欲望和信念)并在更抽象的层面上编码这些信息的关联皮质区域(特别是所谓的默认网络的较大部分),这些信息以性格特征和自传的形式表现出来——这表明某人是什么样的人(例如,[4, 5]的元分析)。关键的皮质区域是颞顶联合区(TPJ),它用于推断他人的意图和信念,而前内侧前额叶皮质则负责对如性格特征和偏好等抽象且稳定的人格进行推断。相反,社会镜像是一种进化较早的功能,它激活了负责检测和理解人体部位(例如,肢体)生物运动的感觉运动区域,比如抓住一个杯子并自动理解其目标——为了喝水(例如,元分析[6, 7])。重要的皮质区域是后上颞沟(pSTS),它检测生物运动,关键的镜像区域包括前内侧顶叶沟(aIPS),它将特定的运动与其典型的上下文联系起来(即,用精细的手势在桌子上抓杯子或用全手抓在洗碗机上),最后是预运动皮质(PMC),它确定其潜在的目标(即,用于喝水还是清洁)。虽然部分区域属于不同的神经回路,但pSTS和TPJ是感觉运动和言语超模态输入的关键整合器,它们部分重叠,其中pSTS位于TPJ的下方,这表明身体阅读和心理阅读经常会相互作用。
许多研究人员在该领域提出的主要假设是,小脑在学习、自动化和微调运动行为序列中的角色是如何扩展到涉及社会行为和交互序列(例如,[8])的社会领域的。为了测试这种序列假设,已经开发出超越传统社会认知及其损害的研究方法,以确定社会行为和行动预测中假定的序列角色。举例来说,小脑研究迅速纳入了社会理解他人心理的关键任务之一:错误信念测试。这个测试涉及的故事有一个角色,他在不在场的情况下不知道一个物体已经被移动或改变了。因此,参与者必须意识到这个角色缺乏关于这种改变的信息,所以他或她对物体的位置或特性持有一个“错误的”信念,这个信念不再是真实的,与现实相冲突[9,10,11]。区分自己看到的现实和他人(错误的)信念,是一个社会能力,只有到四岁时才能完全发展。小脑在错误信念故事中的假设序列角色是相当明显的:一个人离开房间是在另一个人隐藏一个爱玩的玩具之前还是之后,或者说一个秘密等,这都有很大的差异。在小脑的研究中也引入了方法论的进步。这不仅包括神经影像学方法如功能磁共振成像(fMRI)来研究人类小脑中的激活区域,以及这些区域如何使用新的方法如静息态连通性和动态因果建模(DCM)与大脑皮质功能性地交互,还包括非侵入性小脑神经刺激如经颅磁刺激(TMS)和经颅直流电刺激(tDCS)的新用途。
对小脑在社会认知中的重要角色的共识正在形成,但这个领域还处于早期阶段,正在全面发展中。许多早期的发现和洞见正在涌现,其中一些已经被证明是可复制的,这预示着一个实质性证据体系和对社会小脑更好理解的开始。然而,一些结果还处于初步阶段,需要谨慎对待,而其他一些研究则指出了改进和进一步研究的方向。这篇共识论文提供了一群在新兴的小脑和社会处理领域拥有成熟或初级专业知识的科学家的观点和报告。鉴于对小脑社会功能的了解对于更好地理解和治疗各种社会障碍具有巨大的潜力,本文的时机正好。
作者贡献概述
本共识报告首先由Larry Vandervert讨论了制造石器对社会小脑的潜在进化作用。
随后的部分涉及小脑在心理阅读中的作用。我们首先介绍了Maria Leggio提出的社会小脑的排序假说,这是对小脑传统运动观点的扩展,并影响了许多当前关于小脑在社会理解和预测功能的研究。Xavier Guell、John Gabrieli和Jeremy Schmahmann进一步阐述了小脑在社会认知和其他运动和非运动领域的关系。他们对小脑中的双重任务和过程梯度的深入分析和概述,再次证明了小脑对社会认知的领域特定贡献。在他们的新的元分析中,Qianying Ma和Frank Van Overwalle进一步证实了小脑的Crus II主要涉及社会精神化。最后,Frank Van Overwalle及其同事Elien Heleven,Qianying Ma和Min Pu在新的实证贡献中报道了对Leggio的排序假说的若干测试。
接下来,报告了有关身体阅读和行动理解的研究结果。Marco Michelutti和Arseny Sokolov对非语言身体动作(例如,通过点光源或小标记附着在主要关节上,而身体的其余部分则不可见)和象征性的几何形状动画的研究进行了概述。Chiara Ferrari和Zaira Cattaneo讨论了涉及生物运动感知的小脑区域的因果作用。值得注意的是,他们在不同的时间点应用了TMS,以描绘不同区域的小脑过程的时间顺序。
进一步的章节阐述了与小脑相关的临床方面。这为临床实践中治疗患有神经退行性、精神疾病和神经发育疾病的患者提供了新的视角。Silvia Clausi,Michela Lupo和Maria Leggio对小脑在理解心理状态方面的作用的临床含义提供了一个概述,这可能是小脑病患者以及具有社交障碍(如自闭症)的个体报告的社会认知困难的基础。Giusy Olivito,Libera Siciliano,Frank Van Overwalle和Maria Leggio记录了小脑-大脑理解心理状态网络之间的交互或连接及其临床含义。这些证据揭示了多个小脑-大脑网络的功能活动和连接减少,导致在较低级别的镜像和复杂的高级别的心理化方面的损害。接下来,Laura Rice和Catherine Stoodley关注小脑对特定人群社交行为的贡献:患有自闭症的个体。他们讨论了动物和人类关于小脑结构和功能连接性的有前景的研究,以揭示这些人群中小脑异常的起源和后果。
最后一节关于神经刺激的部分,主要关注通过小脑神经刺激改善社交功能障碍的可能方式。Kim van Dun和Mario Manto讨论了社交小脑作为非侵入性神经刺激在各种社会认知障碍中的有希望的目标,而Elien Heleven和Frank Van Overwalle则从一项关于小脑TMS对社会顺序性表现影响的初步试验中提供了初步证据。
我们通过强调一些稳健的研究结果并指出一些缺乏证据的冲突和问题,以及进一步研究的问题。
参与社会认知的小脑区域
为了让不太了解的读者能充分理解本共识报告的结果,我们可能需要在引言的结尾简要描述一下小脑在社会处理中的功能解剖学。然后,我们也简要介绍了小脑在社会处理中执行的计算。
关于功能解剖学,明显地,与运动相关的镜像移动任务更倾向于激活Buckner及其同事们[12]确定的“体感运动”网络。在小脑中,这些网络主要位于前部小脑区域。对于非运动相关的心理化任务,位于后部小脑的“默认/心理化”网络被激活。这可以在图1中看到,该图显示了在NeuroSynth(一个大规模、自动合成fMRI数据的互联网平台,参见[13])中定位了给定的感觉运动行为观察/镜像与心理状态推理的活动。在这个平台上,我们选择了由关键词“行动”和“镜像”以及“心理化”指定的两个元分析,然后在Buckner及其同事们[12]的七网络小脑结构上定位主要的活动区域。如图所示(图1,右侧面板),“行动/镜像”任务,如观察人类手臂的运动(例如,[14])或者点光体运动显示[15,16,17],在前-45和后-75 y坐标之间(以绿色的MNI坐标表示)显示出活动。预期的一样,活动位于体感感知整合网络,但也在腹侧注意力网络中。另一方面,“心理化”任务,包括以人类方式移动的几何形状的动画[18],以及其他人的更高级别的心理状态、信念和个性特质[2],在后-84 y坐标处(以红色的MNI坐标表示)显示出活动。这种活动主要位于心理化/默认网络[12]中。
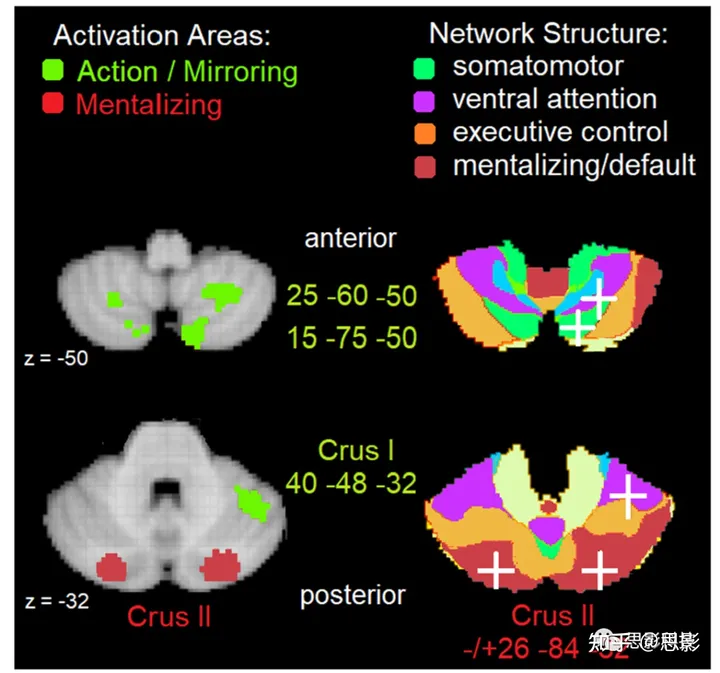
图1 横断面视图展示了位于MNI z坐标-50和-32的小脑下部和上部。
[左]自动元分析的NeuroSynth(50个主题)中小脑最活跃的区域[右]叠加在Buckner等人[12]的7网络结构上,坐标以白色十字表示。三个与NeuroSynth中的“行动”和“镜像”关键词(#19)相关的绿色“镜像”区域是绿色体感运动整合和紫色腹侧注意网络的一部分;两个与NeuroSynth中的“心理化”关键词(#8)相关的红色“心理化”区域是红色心理化网络的一部分。
图1中的NeuroSynth分析与Van Overwalle等人对社会小脑的元分析([2]; 见表1)大致一致,并且他们对Buckner及同事的7网络结构的功能解释也大致一致[12]。镜像任务主要调用了体感运动网络,但表1也显示了强大的左半球簇(在NeuroSynth中相对较小),并再次证明了腹侧注意网络中的一个簇。心理化任务证实了在后坐标-84 y坐标之后的默认/心理化网络的主导活动,但也在前叶IX。表1提供了关于自我心理化的额外信息,这激活了边缘系统和体感网络边界的前叶IV和VI,这可能反映了本体感知和情绪触发的体验。
表1 Van Overwalle等人[2]的元分析及其对Buckner等人[12]的7网络解释。
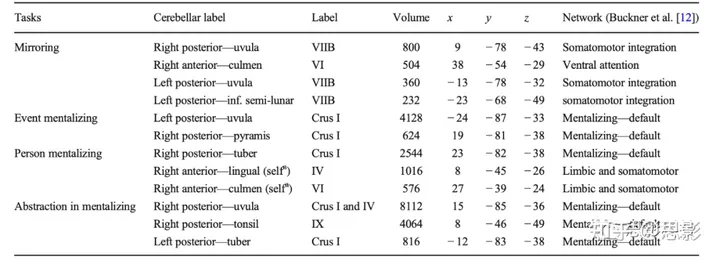
注:根据ALE和Schmahmann等人的图谱[19]给出解剖标签。每个集群的体积在200立方毫米以上;x–y–z坐标转换为MNI a主要参与自我参照的簇。
参与社会认知的小脑计算
大脑最适应的功能之一是预测即将到来的感官、运动和认知状态,并纠正这些预测中的错误,以避免在未来重复这些错误。在这个过程中,小脑被认为具有生成内部模型的核心功能,这些内部模型是对环境、代理和事件的内部表示,包括基于这些表示对未来后果的预测。根据Leggio和Molinari提出的序列假说([8];见[20];以及Maria Leggio关于"社会小脑的序列假说"的贡献),这些内部模型的主要功能是学习和表示时间结构事件的重复模式,或序列。这个功能由前向内部模型[21]捕捉,它们不仅表示事件序列如何随着时间的推移展开,而且还预测它们的后果,如对外部环境的影响以及对自身的本体感觉经验的影响[22]。这些序列预测基于从大脑皮层接收的信息(即,复制的反馈),如从皮质镜像网络中的pSTS(后上颞沟)接收到的关于另一个人运动的社会推断[16],或从皮质理解网络中的TPJ(颞顶联合区)接收到的关于另一个人心理状态的信息[23,24]。当预测不匹配时,前向模型向皮质发送外感和本体感预测错误[22]。这使得皮质能够在重复行动、观察或认知过程时逐渐减小未来的错误。当错误反复出现时,这可能会让小脑适应环境中的系统性变化,或者为不同的环境开发新的模型[25]。这种关于减少预测错误和适应现有模型的观点与预测编码[26,26,28]和监督(即,纠错)的神经功能连接模型[25, 29]相一致,这些模型也已经被应用到社会认知中([30, 31];回顾[32])。在社会认知中,例如,当另一个人执行的一系列行动暗示一个错误的信念时,我们立即意识到这个人可能会犯的潜在错误。如果进一步的行动并未证实这个预测,并暗示这个人毕竟是正确的,错误预测立即告诉我们调整我们的错误信念推断,并预测这个人的未来行动。
通过从皮层到小脑的一系列并行闭环,并返回皮层的方式,完成内部模型的循环通信和调整。小脑结构和生理的均一性,暗示了对来自皮层的入射信号进行类似或“普遍”的计算[33],但也暗示了由这些闭环在小脑中终止的不同区域决定的功能域特异性([21, 34];另请参见Xavier Guell、John Gabrieli和Jeremy Schmahmann关于“小脑社会认知与其他运动和非运动领域之间的关系”的贡献)。然而,也有可能,统一的小脑回路进行一系列多种计算和功能,这些功能或多或少的不同,而不是普遍的,由不同的皮质输入驱动[34],并且这些功能随着小脑和大脑皮层的进化扩张而并行演化。
除了序列本身,事件序列的时间在毫秒到一秒的范围内也是小脑在生产和理解观察事件中的非常重要的功能[25, 35]。在社会领域,时间在行动观察、协调和互动中至关重要,例如在使用虚拟平台进行社会互动时,当经历声音和视觉的小传输延迟时,就会体验到困难。然而,尚不清楚在理解他人思维过程中,毫秒级的时间到底有多重要,因为这个认知过程在相对高的抽象级别上运作,大部分都没有快速的环境反馈。这是一个有趣的研究课题。
内部模型的一个独特类别生成了用于追求自己期望的(社会)目标的自身行动序列的运动命令和预测,被称为内部逆向模型[22]。逆向一词反映了从目标到行动的反向推理,以推断所需的行动步骤。这些逆向行动模型在计划和协调高效社会互动和合作所需的行动中至关重要。例如,它们计算出身体(例如,视线)的预期位置或表达,然后估算需要将当前的位置或表达转变为预期的运动命令;或者,它们计算出自己在一个群体中的预期社会角色(例如,在中心位置担任领导),并估算出从当前角色转变为预期角色所需的行动。
小脑驱动的社会学习内语在石头工具制作和语言的演化中:大脑-小脑系统中先天的手-工具连接(Larry Vandervert)
Vandervert[36, 37]认为,由于石头工具和语言的演化需要(1)重复性和(2)社会学习他人的行动,所以这一演化过程主要由小脑驱动。他提出,这种重复的社会学习是在理论心理(ToM)(人们对他人心理状态推断的模拟能力)[20, 38,38,40]的框架内进行的。Vandervert基于人类学家Dietrich Stout和神经人类学家Erin Hecht[41]对学习他人制作石头工具所必需的严格技能发展的详细分析,提出了这个论点。他们的一些相当详细的发现部分包括以下学习者所需的社会和认知技能发展的关键方面:
因此,敲击的社会复制的关键瓶颈就是所需要的长期练习[斜体强调]以达到感知-运动的能力。这需要掌握关系,例如力的大小和敲击位置与核心的形态、位置和支撑之间的关系[42,42,44],这些对于无知的观察者来说并不能直接感知,并且不能以语义知识的形式直接传达。(p. 7862)
Vandervert [36, 37]指出,在他们对石头工具敲制和大脑演化的整体描述中,Stout和Hecht[41]没有提到(1)小脑在社会化技能发展中的作用[39],以及(2)Marvel和Desmond[45]以及Marvel, Morgan和Kronemer[46]发现的小脑驱动的内在或静默语言在工作记忆中的作用。
心智理论通过内在语言的进化出现
斯托特和赫希特[41]上述的描述似乎主要涉及观察学习,而在石器制作的社会复制中,几乎没有或没有任何与目标相关的口头信息。因此,有些人可能会质疑,就像范德弗特[37]所建议的那样,学习制作石器是否真的涉及到心理化(即理论思维)。在这方面,范·奥弗瓦莱和巴滕斯[7]讨论了大脑中镜像系统和心理化系统的区别,其中心理化部分涉及与目标相关的口头信息。因此,本文的立场遵循范德弗特的[37]关于理论思维的演化方法,他提出石器制作的演化逐渐产生了理论思维能力,源自于预言性的无声自我对话。范德弗特提出,这种预言性的无声自我对话与演化早期的心理化有关,而小脑的预测、误差修正以及这一高度社会化过程的自动化的选择优势,导致了工作记忆中的语音循环和语言。也就是说,语言的演化主要不是来自交流,而是来自与学习者构建与制作石器技能的严格和社会环境相关的理论思维相关的内语。范德弗特认为,这种语音循环到语言的演化是由新的复杂的目标相关注意力模式驱动的,这些模式越来越细致且重复地关注到敲击的因果关系要求,主要是在过去的一百万年中,通过一个新出现的社会认知小脑进行调节的[47,47,49]。因此,虽然公开的语义教学可能对习得/教授石器制作没有帮助,范德弗特提出,无声的自我对话(自我谈话)和早期的理论思维,以及感知运动学习,是习得/教授这些敲击技巧的关键。
Vandervert的[37]上述关于ToM社会起源的关键元素得到了以下与石器制作-内语-社会小脑起源的语言工作记忆相关的研究的具体支持。
1.语音编码导致了语言工作记忆的进化获取[50]。
2.内语随任务需求的增加而增加[51]。
3.年轻学习者的私人内语随任务需求的增加而增加[52]。
4.黑猩猩和人类学习的关键区别在于,人类更倾向于关注自己和他人(社会)行为细节的倾向[53]。
5.当语言信息进入语音存储时,会发生特定的小脑-后顶叶处理[45, 54]。
本文的目的是为小脑驱动的石器制作演化的社会学习解释提供进一步支持。这种额外的支持基于以下发现:(1)在大脑中存在与天生的手-工具重叠[55],(2)在侧后小脑中存在用于实际和想象工具使用的特定工具模块[56, 57]
大脑中的天生手-工具重叠以及侧后小脑的工具模块
至少有两条重要的证据支持Vandervert的[36, 37]论点。首先,Higuchi, Imamizu和Kawato [56]以及Imamizu和Kawato [57]发现,工具的实际和想象使用都在小脑中被模块化(有剪刀、锤子、螺丝刀等特定的模块)。这些工具使用的模块化模型(尤其是工具的想象使用)主要存在于新演化出的侧向小脑半球中,这些小脑半球在过去一百万年里大大扩展。小脑的齿状核向大脑皮层发送实际工具使用和想象工具使用的模型,这样它们就可以在大脑皮层中被有意识地体验到[58]。其次,在研究无手症患者(出生时没有手的人)时,Striem-Amit, Vannuscope和Caramazza [55]描述了在大脑皮层的枕-颞区域演化出的一个天生的手-工具重叠区域,这个区域用于接受将工具握入手中:
手-工具重叠的出现是因为手和工具作为一个共同(或紧密交织的)专门系统的部分,可以进行有效的处理,而这带来了潜在的优势[工具是有利的辅助工具。这个系统,反过来,又与背部的行动处理区域[顶叶皮质]相连接,以便快速有效地塑造手去抓取和使用工具[这需要进化和发育的小脑的精细化]。一旦进化出来,这个先天决定的系统会在发育过程中表现出来,即使在没有任何特定输入的情况下,就像无手症患者一样,这些输入最初是为了提高模式的完全有效性。(p. 4790)
尽管Striem-Amit, Vannuscope和 Caramazza并未特别提到石器制作的演变导致了手-工具的重叠,但这里提出的是这种先天的手-工具重叠在大脑中的演化至少发生在过去的一百万年[47, 49, 59],这段时间里,石器制作和石器使用的技术不断提升,社会-认知小脑也在扩展。
将天生的手-工具重叠与侧向小脑中的工具模块化结合起来
根据Van Overwalle, Van de Steen, 和 Mariën的研究[23],我们建议在大脑皮层的颞顶联合区(Striem-Amit, Vannuscope, 和 Caramazza[55]的手-工具重叠区)和侧向后部小脑([56, 57]工具模块)之间建立一个闭环的社会心理化连接,这将共同优化工具制作的社会认知,在心智理论构建中[36, 37]。此外,我们进一步建议,在这种工具制作的社会心理化过程中,小脑将主导优化手的形状以及抓握的动力学[60]。
结论
随着小脑驱动的内语介导的理论心智的生产的精细化的演进,工具与手一同嵌入到天生手-工具重叠的区域特化[55]以及小脑的实际和想象的工具模块化表示中[56, 57]。此外,由于小脑显然对抓握动力学的精细化至关重要[60],并且根据Stout和Hecht在本文开头的分析[41],这种精细化是由社会驱动的,工具的演变大部分是由演化中的社会小脑产生的,正如Van Overwalle, Manto, Leggio, 和 Delgado-Garcia所描述的[20]。Vandervert[36]提出,这个关于社会小脑的故事在很大程度上是智人崛起的故事。
小脑与心理阅读
这一部分首先介绍了Maria Leggio提出的社会小脑的排序假设,这是对小脑传统的运动观点的扩展。Xavier Guell、John Gabrieli和Jeremy Schmahmann详细阐述了小脑中社会认知与其他运动和非运动领域之间的关系,并为小脑对社会认知的领域特定贡献提供了进一步的证据。Qianying Ma和Frank Van Overwalle提出了一项新的元分析,该分析记录了小脑十字架II主要参与社会心理化。最后,Frank Van Overwalle及其同事Elien Heleven、Qianying Ma和Min Pu在新的实证贡献中报告了对Leggio的排序假设的几项测试。
社会小脑的排序假设 (Maria Leggio) 社会认知的一个基本组成部分是估计他人的心理状态的能力[61, 62]。理解他人的心理状态需要创建一个对个体的心理模型,并能预测他们的心理状态可能如何影响他们的行为[21]。这个过程也使我们能够认识到社会互动的结果何时偏离了我们的期望,并使用这个信息来校准未来的社会预测[21]。
在复杂的心理化过程中,预测是通过基于人类行为的内部模型实现的,这些模型是基于预期行动将是有效的,并且与个人信仰、性格特征和社会规范一致的[62]。有人建议小脑在预测处理中起作用,作为一种前向控制器[20,21,63],并且序列检测可能是其操作模式[8,64,65]。实际上,根据"序列检测理论",小脑检测并模拟时间或空间结构事件的重复模式,无论它们是否构成运动计划中自身行动的感官后果,预期的感官刺激在感知预测,或者更高级别的过程(例如,认知过程)的推断[8,64,65]。这种模拟使得可以创建内部模型[66],并且这些内部模型可以用来对包含任何类型组件的未来事件做出预测,例如身体,其他人,或者环境。
在更复杂的概念层面,有人提出小脑参与在社会互动过程中构建心理过程的内部模型,其中预测序列事件起到了核心作用[20,38]。事实上,社会心理化,社会认知的更反思和意识的组成部分,有能力将心理状态归因于他人,并采取他人的观点来预测即将发生或未来的社会行为[67,68]。因此,与感官运动领域的信息处理类似,小脑可能通过检测社会上可预测的序列(例如,社会行动的内部模型)并推动完成这些功能所必需的优化前馈控制,来调节更高级别的皮质活动[23,69,70],这样可以以流畅和自动化的方式进行[20,21,38]。通过这种方式,可以实现社会互动的两个主要要求:理解和预见自己和他人的行动,并理解对自己的后果,以及识别社会互动预期结果中的偏差,以修改未来的社会期望[20,38]。
在一项针对健康受试者的最近的fMRI研究中,Heleven及其同事们[71]显示,构建需要理解主角心理状态的社会行为序列(例如,涉及错误或真实的信念)强烈激活了后小脑,主要是Crus I–II,它们涉及到社会认知的更复杂和抽象的方面[21,38]。这些数据与证据一致,这些证据显示当社会预测被违反时(例如,违反社会规范)小脑被激活[72]。
有趣的是,Clausi及其同事们[38]发现,患有退行性小脑萎缩的病人不仅在估计他人心理状态的低级别和自动化过程中(例如,身体阅读)受损,而且在更复杂的概念层面的心理化中也受损,这一点通过在高级ToM任务[73]和社会“失态”故事[74]中的受影响的表现得到证实。在这些任务中,序列事件是出乎意料的和模糊的(例如,当需要准确识别一个角色的言论背后的潜在意图,而这并非字面上的真实,或者理解一个说话者说了一些东西,而没有考虑到听者可能不想听或者可能会受到所说的东西的伤害),需要不断比较事件和社会期望,以及高水平的预测[38]。另一方面,当故事中的模式需要较低级别的预测和错误监控时,例如在“无失态”故事和情绪归因测试[75]中,小脑病患者表现出良好的表现[38]。
为了进一步支持小脑在生成适当的社会行动序列中的特异性,Van Overwalle及其同事们[3]描述了被小脑退行性疾病影响的患者在执行错误信念任务的序列版本[76]时的能力受损。考虑到这些证据,我们可以概念化地认为,小脑在不同领域都是一个独特的预测结构。就像在感觉运动控制中一样,在社会认知中,小脑可能通过匹配外部信息(社会输入)与内部模型(与先前经验相关的特定社会事件)来行动,从而有助于形成对他人心理状态的判断。因此,当小脑受损时,外部刺激和内部模型之间所需的快速连续的信息交换可能会受到影响,从而干扰小脑识别社会互动结果中的偏差/错误的能力,以及它使用这些信息来调整和调节未来社会期望的能力[38]。
与这个理论相一致,已经在受小脑损伤影响的患者中描述了参与不同方面的社会互动的小脑皮质网络中的结构和功能改变[21, 38, 77,77,79]。更多细节在Olivito等人的“小脑-大脑心理化网络内部的连接性和临床意义”一文中报告。
被认为受损的序列化和预测机制会影响到许多以小脑-大脑功能障碍为特征的神经精神病和神经发育病理的社会能力[20, 21, 38, 77, 79]。在这个框架内,举几个例子,例如在精神分裂症中,前向建模的改变被认为是幻觉的原因,因为无法区分内部状态和外部事件[80, 81]。在自闭症谱系障碍中,主要的行为标志是识别和归因他人的心理状态以解释和预测他们的行为的能力受损[82]。Clausi等人的“小脑在心理化中的作用的临床意义”一文中报告了小脑病患者和自闭症患者的心理化能力的比较。
总的来说,小脑在感觉运动领域的自适应控制和预测编码的典型角色需要扩展到社会认知领域,因为预期、适应和学习似乎对成功的社会互动和自适应的社会行为是必不可少的。
通过人类功能性磁共振成像探索小脑社会认知与其他运动和非运动领域的关系:来自Xavier Guell, John D.E. Gabrieli, Jeremy D. Schmahmann的见解。
像人类脑连接组(HCP)[83](n = 1003)这样的大型fMRI数据库使我们有可能以前所未有的力量分析人类小脑的组织。我们对小脑任务和静息态HCP数据的分析已经在小脑中识别出非运动任务激活的三重表示[84],描述了小脑皮质中的功能梯度[85],并在迄今为止分析的最大数据集中描述了小脑基于任务的功能地形图[84]。在HCP中的社会任务对比了一个理论化(理论思维)的条件,参与者观看社交互动的移动几何物体,减去了一个随机条件,显示随机移动的几何物体[86, 87]。HCP参与者还完成了静息状态、运动、工作记忆、情感和语言fMRI任务。在这里,我们通过HCP索引的小脑皮质中的其他运动和非运动领域的社会相关过程进行分析,特别是通过任务激活地形图,多重表示的原则和功能梯度的视角。
任务激活分析
显示社会认知与其他功能领域的领土重叠的任务激活图与现有观点一致[88],即小脑对社会认知有特定领域的贡献。社会任务主要涉及十字形叶I和II以及叶IX(图2a;箭头“1”指向IX激活)。中等效应大小的阈值显示,社会任务与运动、工作记忆或情感任务没有重叠(图2a)[84]。社会激活与语言激活重叠(图2a,见箭头“2”),但这种重叠可能是由于两项任务之间的心理共性。具体来说,HCP的语言任务对比了一个故事条件,参与者在这个条件下听故事,减去了一个数学条件,参与者在这个条件下回答算术问题。因此,HCP对语言处理的评估中存在一个理论化的成分。此外,通过使用数学作为对照条件,减去了注意力任务聚焦的需求。结果,小脑语言激活在很大程度上类似于任务非聚焦、默认模式处理的地图(见图2a中的语言与默认模式)。小脑皮质中的社会和默认模式处理的重叠已经被描述过[88],这种重叠与大量证据支持的默认网络在社会认知相关过程中的作用,特别是社会思维[89, 90]是一致的。由于在小脑中,社会和非社会任务激活没有重叠,因此推断出小脑对社会认知有特定领域的贡献[84, 88]。
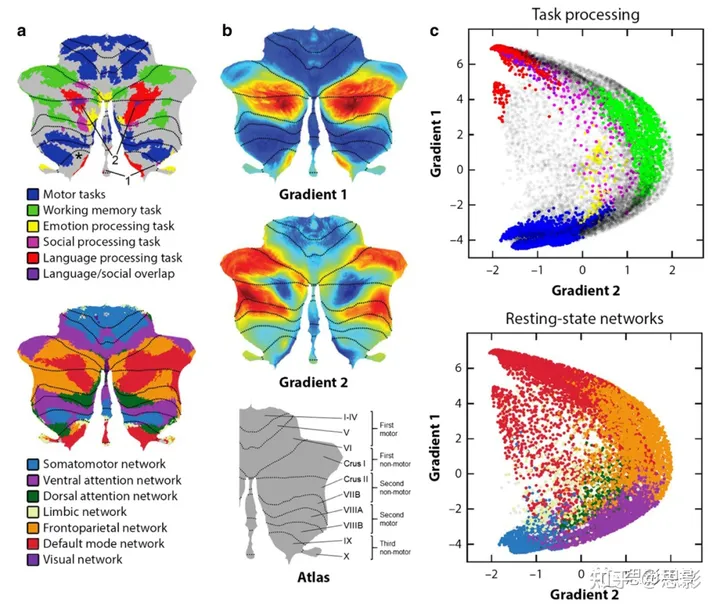
图2 a 小脑任务激活地图[84](上)和静息态网络[12](下)。
1 = 指示IX叶情感处理激活(为了清晰)。2 = 指示语言/社会重叠区域(为了清晰)。星号(左IX叶)= 如果使用较低效应大小阈值,如[84]的补充材料中所示,指示工作记忆任务激活的区域。b 小脑功能梯度[85]。地图表明每个运动和非运动表示的位置[84]。c 功能梯度1和2与任务激活地图(上)和静息态网络(下)的关系。每个点对应一个小脑体素;每个点的垂直/水平位置对应该体素的梯度1/梯度2值;每个点的颜色表示每个体素是否属于特定的任务激活(上)或静息态网络(下)地图[85]。
通用组织原则
一种不同的探究方法检查了社会和其他运动和非运动领域共享的一般组织原则。在已经被广泛确认的I-VI和VIII叶中有双重运动表示的描述之后,我们的分析表明,小脑皮质中也存在多重非运动任务处理的表示。具体来说,小脑皮质中的所有非运动过程可能同时涉及一些VI/Crus I叶(第一非运动表示)、Crus II/VIIB叶(第二非运动表示)、以及IX/X叶(第三非运动表示)的方面[12, 84](参见图2中的地图,指示每个表示的位置)。值得注意的是,第一和第二非运动表示可以是连续的(如在语言任务或默认网络中所见,见图2a)或是分开的(如在工作记忆任务或前额顶叶网络中所见,见图2a)。HCP中的社会处理在Crus I/II叶表现出第一和连续的第二表示,并在IX叶表现出第三表示(参见图2a中由"1"箭头指示,指向IX激活)[84]。因此,小脑社会神经解剖学在更大的三重表示原则中得到了情境化,这个原则在小脑皮质的许多(可能是所有)非运动领域中都是共同的。
功能梯度在小脑中
通过将社会任务激活与小脑功能梯度[85]相映射,可以获得关于小脑社会认知的解剖和心理结构的额外见解(见图2b)。这些梯度定义了小脑皮质中功能领域的位置和关系。梯度1解释了功能连接模式中的最大变异量,并从运动处理区域扩展到默认模式处理区域(见图2c)。梯度2则突出了专注于任务的注意力/执行过程。HCP社会任务沿着梯度1的范围广泛分布,有些偏向于高梯度1(默认模式)领域(见图2c顶部的粉红色和紫色)。这种广泛的定位与其他HCP任务形成鲜明对比,如语言任务(听故事减去数学;图2c顶部的红色)主要定位在梯度1的高值处,运动任务主要定位在梯度1的低值处(图2c顶部的蓝色),工作记忆任务定位在梯度2的高值处(图2c顶部的绿色)。社会任务在功能梯度空间中的广泛分布与小脑中的多模态社会处理理念相呼应,沿着小脑功能神经解剖的主要维度参与多层次的信息处理,而没有在其任何极点(默认模式,注意力/执行或运动)上独占定位。以这种方式,这些数据说明小脑的社会认知可能涉及包括感觉运动,注意,非注意,外向和内向思考在内的多种脑功能模态。情绪处理在功能梯度中也观察到类似的中心位置(图2c顶部的黄色);情绪处理并未完全符合默认模式(梯度1的高值),运动(梯度1的低值)和注意力/执行(梯度2的高值)的划分。
揭示小脑中社会处理的多重表征的意义
通过将社会处理与小脑功能神经解剖学中的其他运动和非运动领域进行比较,可以阐明小脑中社会处理的多重表征(Crus I/II中的第一和连续的第二表征,IX中的第三表征)的意义。每个运动和非运动表征领域沿着小脑功能梯度1和2的位置表明,不仅在两个运动表征之间,而且在三个非运动表征之间可能存在功能差异(参见[85]的第三图)。此外,由于第二运动表征和第三非运动表征都位于功能梯度1和2的更中心位置,与第一运动表征和第一/第二非运动表征领域相比,可能第二运动表征与第三非运动表征有功能相似性(如在[85]中讨论的)。因此,对小脑第八叶在运动处理中的角色(与小脑I-VI叶的运动处理相比)的理解可能会提供对小脑第九叶在社会处理中的角色(与Crus I/II的社会处理相比)的理解。
社会认知作为思维失调理论的典型例证
这里提出的神经影像学发现,以及它们对社会认知与其他运动和非运动领域关系的意义,与普遍的小脑变换(UCT)和思维失调理论有强烈的联系[1, 33, 92,92,93,95]。普遍的小脑变换假说认为,所有小脑对行为的贡献都是由一个单一的神经计算支持的。这个理论基于两个相对而言的互补现实:小脑皮层的结晶重复细胞结构[96],与连接小脑半球和核区与不同大脑运动、认知和情感区域的地理精确的解剖连接图[33, 92, 97,97,98,99,100,101,103]形成鲜明对比。小脑皮层的均匀结构使得一个独特的计算,即普遍的小脑变换,能够调节大脑半球中的多个信息处理流,包括社会认知。思维失调理论是普遍的小脑变换假说的推论。因为所有小脑对行为的贡献都源于一个均匀的神经计算,所以小脑损伤引起的神经症状和体征反映了一种常见的神经功能障碍,即失调。在运动领域,小脑病变导致运动失调,并降低了运动控制的协调性、精确性和流畅性。在认知和情绪领域,小脑功能失调导致思维失调,损害了思维和情绪的协调性、精确性和流畅性,包括社会处理[33, 92, 93, 103]。
功能梯度的分析及其与任务激活图的关系源于对行为是分布式神经回路中多个独特节点的共同产物的理解,这些节点在神经系统中地理分布。这对于支持运动控制[104]和注意力[105]的回路同样适用,也适用于复杂的社会和道德推理Fox [106],这是社会处理所必需的,对于小脑和大脑皮层区域、丘脑核团和基底节部分同样适用[107]。例如,虽然小脑第V和第VIII叶物理上距离较远,但它们都位于小脑功能组织主要梯度的运动极点附近。社会认知中也可以观察到小脑任务激活图的空间位置与其在小脑功能梯度上的分布之间的独立关系。社会任务激活图在功能梯度值广谱上的分布表明,小脑的社会处理涉及到多种大脑功能模式,例如外向和内向的思维。这个观察与小脑皮层中存在特定参与社会认知相关过程的不同区域的事实是独立的,也是相容的。支持社会认知的神经回路招募了不止一个大脑皮层区域,以及与那些大脑区域相互连接的不止一个小脑区域。因此,分布式神经回路存在于小脑自身中,这既是大脑-小脑连接的结果,也是小脑中认知的第一、第二和第三表示的结果。这些考虑与指导普遍的小脑变换理论的解剖原理一致,也与思维失调理论一致,后者是提出的小脑在社会认知中的角色的理论基础[108, 109]。
小脑后部Crus II区在社会理解中的领域特异性作用" (Qianying Ma, Frank Van Overwalle)
日益增多的证据表明,小脑支持社会认知[2, 88]。可以说,最先进的人类社会认知功能涉及到解读他人的心智,被称为心理化[5, 7, 110]。它需要洞察他人或自己的精神状态,从理解具体的此时此刻的意图、原因、情绪和信念,到抽象和远离的社会推理,如人格特质以及过去的、未来的或假设的事件[111,110,111,112,115]。社会心理化由小脑后部支持[2, 88],这一部分在进化上较年轻[116],尤其是由小脑的心理化/默认网络支持[12]。
然而,问题仍然是,小脑后部在多大程度上偏好于心理化,如果是这样,小脑后部的哪些区域更偏好?过去的小脑功能性元分析并未报告社会过程,而是报告了经典的感知运动和非运动认知功能,如语义、语言和执行控制[117, 118],或者报告了非常有限的社会任务,如几何形状的生物运动感知,这并不能很好地代表人类的社会心理化[84]。即使是到目前为止最详尽的元分析研究,Van Overwalle等人[2]的研究也没有权衡小脑中社会功能与其他非社会过程的重要性。
为了研究小脑后部哪些区域专门负责社会心理化,Van Overwalle、Ma和Heleven[119]确定了在社会心理化过程中频繁参与的一些感兴趣区域(ROI)。从最近的两个研究中得出了两个“序列化”ROI,这两个研究研究了小脑心理化的一个关键方面:生成需要理解他人信念的社会事件的正确序列(见图3,左侧面板)。ROI 1 来自一个研究社会行为序列生成的功能磁共振成像(fMRI)研究([71]; n = 73),也在早期的研究中被识别出来[24, 39]。ROI 2 是其左侧的镜像位置。这些 ROI 非常接近 Guell 等人的元分析报告中的两个峰值([84]; 约 6-8 毫米远)。另外两个 ROI(图3,右侧面板)从 NeuroSynth 中的自动化“心理化”元分析中提取出来(即,从 2018 年 7 月 NeuroSynth 数据库中的摘要主题 50 个主题中提取;参见 [13])。所有四个 ROI 都位于楔状叶 II,在Buckner等人[12]划定的心理化网络中。选择了 NeuroSynth 中所有位于这四个 ROI 坐标周围半径 6 毫米内的功能性 MRI 研究,条件是它们符合一系列相关性/有效性标准,如坐标以 MNI 模板表示,涉及未用药的健康参与者,提供了一个适当的对照条件,等等。
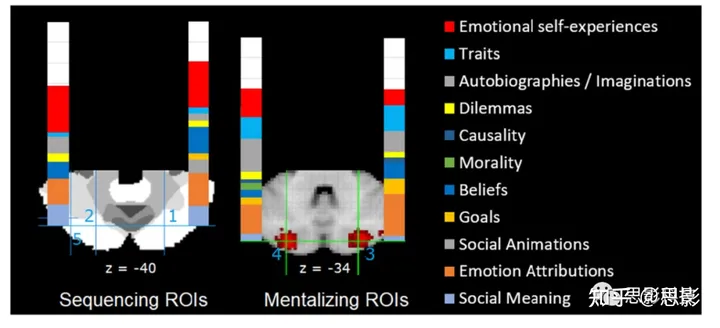
图3 元分析结果以所有识别到的研究的100%比例显示不同的心理化任务子类别。
所有其他非心理化的功能都用白色表示。ROI 1和2(MNI坐标± 25, − 75, − 40)被叠加在Bruckner等人[12]的7网络划分上,其中白色区域反映了心理化/默认网络;ROI 3和4(MNI坐标± 26, − 84, − 34)来自NeuroSynth中的心理化元分析。 每一项选定的 fMRI 研究都根据主要和对照条件的刺激材料,被归类为不同的心理化和非心理化子类别,使用的标准与早期的元分析相似[2, 5, 7]。图 3 的结果令人信服地展示出,“社会心理化”是涉及约 75% 研究的主要功能类别(当也包括情绪自体验时)。在心理化任务中,最高的百分比出现在其他人的情绪归因(27%)、情绪自我体验(17%)、自传或想象的情况(16%)、他人的信念(14%)、他人和自我特质(12%)以及人类环境中的自发社会意义(12%)的子类别中。为了提供每个任务类别的比较基准率,查询了 NeuroSynth 数据库(2018年7月提取;[13]),结果显示心理化任务类别在所有的 NeuroSynth fMRI 研究中的比例低于 20%。这表明,在Buckner等人[12]的心理化网络中选定的 ROI 中心理化研究的高发生率并不是由于一般的更高基准率,而是特定于这些区域。
总的来说,Van Overwalle、Ma 和 Heleven[119]的元分析证明,与社会心理化和自我相关的情绪认知相关的领域特定的社会认知得到了小脑楔状叶 II 中选定区域的支持,平均发生率约为 75%。这指向了楔状叶 II 中对社会心理化过程有高度专业化的区域。这些社会心理化功能涉及对其他人或自我的心理状态的明确推断,或在人的社会环境中隐含的推断。ROI 的起源表明,稍微前置的“序列化”区域更多地捕捉了社会认知的序列化性质,而更后置的“心理化”区域稍微更好地捕捉了不一定依赖于行动序列顺序的社会理解。此外,最近的动态因果建模(DCM)分析证明,“序列化”ROI 通过双向循环有效地连接到双侧 TPJ,这些部分是皮质中负责角度切换的心理化网络的关键部分[23, 24]。

















 2万+
2万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









