





个体在智力方面的差异,利用认知测试得分进行评估,具有良好重复的、层级的表型协方差结构。这些差异在生命过程中具有显著的稳定性,并且能够预测教育、社会和健康的结果。基于这种坚实的表型基础和其对生活的重要性,人们对智力的环境、社会和遗传病因,以及智力差异在大脑结构和功能中的基础产生了兴趣。在这里,我们总结并评述了过去大约10年的分子遗传学(基于DNA)的智力研究,包括与智力相关的遗传位点的发现、基于DNA的遗传力以及智力与其他性状的遗传相关性。我们总结了新的脑成像与智力的研究发现,包括全脑关联以及灰质和白质的关联。我们总结了区域性脑成像与智力的关联,并根据理论观点进行解释。我们讨论了将遗传学和脑成像结合起来研究智力差异的研究。在所有这些领域中,都有新的(虽然是适度的)关联发现,但缺乏机制性的解释。我们试图找出可能有助于形成更综合的“系统生物学”解释个体间智力差异的增长点。本文发表在Molecular Psychiatry 杂志。
个体智力差异
本文讨论了在理解人类智力个体差异病因方面的一些新贡献。重点在于遗传变异和脑成像衍生的差异,包括这两种来源的重叠之处。一个多世纪以来,研究智力差异的领域一直存在一些争议(见框1)。尽管如此,基于稳健的研究发现,智力研究在许多方面达成了共识。本文的第一部分总结了一些研究发现,基于这些发现,智力差异的还原主义方法(包括脑成像和遗传学)得以发展。
| 框1:智力领域的一些争议与共识 以下是关于人类智力差异的一些争议的简要列举。Galton提出认知能力可能是一般性的、正态分布的,并且具有一定的遗传性(现代数据支持这些观点)。Spearman 发现了认知表现评估之间的正相关矩阵,并提出了智力的双因素理论,包括他称之为 g 的“一般智力”和他称之为“s”的特定能力。Thurstone 和 Gardner 等研究人员持不同意见,认为存在多种独立的智力。Thomson 提出了一个巧妙的理论,解释了如何在没有 g 因素的情况下出现认知测试相关矩阵。是否存在 g(这是一个可重复的统计发现以及它的意义(尚未知道)自那时起就一直被讨论。Henry Herbert Goddard 将 Binet 的智力测试(第一个发明的智力测试)引入美国,并被记录为过度和误用它(Zenderland)。Gould 在他的著名著作《人的误测》中强烈批评了智力中的 g 因素,认为这是应用统计分析方法的必然结果(这是不正确的),并批评了智力与脑容量之间的关联。该书因这两个错误而受到批评(Carroll)。Flynn 发现智力测试得分在 20 世纪中叶的几十年中有所提高。然而,Flynn 也明确表示,这不会改变同一群体内智力测试得分的可靠性、有效性和遗传性。然而,智力测试得分“Flynn效应”的原因仍然是个谜。Herrnstein 和 Murray 撰写了一本名为《钟形曲线》的书。他们分析了 1979 年美国国家青年纵向调查的数据,发现青少年/成年早期的高智力与 30 多岁时的较好生活结果有关。该书因处理种族群体差异而受到强烈和广泛的批评,尤其是因为其分析未通过同行评审发表。 1996 年,美国心理学会(APA)对智力研究进行了平息争议的工作。由于《钟形曲线》引起的争议,APA 组建了一个由认知心理学权威 Neisser主持的特别小组,向非专家说明关于智力测试得分差异的(可靠)已知和(尚未)未知内容。特别小组的 11 人——成为共著文章的合著者——令许多人感到惊讶。他们是观点多样的专家——例如,有些人更倾向于环境,有些人更倾向于遗传,有些人与包括 g 在内的智力差异的分层模型相关联,有些人则持不同的智力模型观点。然而,他们写了一篇关于智力研究的一些坚实基础的文章。讨论的许多主题中,他们承认心理测验方法在智力差异研究中的重要性,并总结了智力测试得分的稳定性、对教育、工作和其他生活结果的预测效度、环境和遗传起源、Flynn 效应以及各种类型的群体差异。APA 特别小组报告仍然是获取关于智力的相对客观和一致总结的必读材料。他们列出的智力未知点仍大多处于这种状态;其中一个是遗传学,我们在此讨论,并且该领域已取得了相当大的进展。我们建议阅读其他更多关于智力已知和未知的最新总结,尽管我们希望读者意识到有些总结来自更倾向于社会/环境的作者群体(Nisbett et al.),有些则来自更倾向于遗传的群体(Gottfredson),以及两篇来自本文的一位作者(Deary)。在寻找智力的社会和环境原因方面的进展可以说不如本文总结的生物学研究成功,尽管 Nisbett 等人的综述讨论了许多增长点。此外,Neisser 等人和 Nisbett 等人的总结还涉及脑成像和遗传学,为本文概述提供了有用的背景资料,Haier 的书《智力的神经科学》也是如此。为了避免读者过度扩展智力的重要性,APA 特别小组强调——正如我们所做的——人类差异中有许多认知和非认知方面不被智力测试和一般智力捕捉到。g 可能很重要,但远非一切重要的东西。 |
描述智力的表型
我们需要向读者澄清,“智力”只是用于描述人类思维技能差异的术语之一;其他有时作为近义词使用的术语包括认知能力、认知表现、认知功能和心智能力。有时也使用 IQ(智力商数),尽管在心理测量学领域内它有特定含义。智力(或前一句中列出的其他术语)作为一种人类表型,是通过认知测试来测量的,这些测试有成千上万种。这给了怀疑论者一种武器,他们可以轻率地否定这个研究领域,因为如 Boring 在 1923 年著名写道,“……智力作为一种可测量的能力必须一开始就被定义为在智力测试中表现良好的能力。智力就是测试所测试的东西。”这句被广泛引用的短句并不是 Boring 的观点;相反,他是在说,如果不了解研究结果,可能会这样认为。他的下一句开始,“这是一个狭隘的定义,但它是严谨讨论测试的唯一出发点”。我们将在这里进行严谨的讨论。在此之前,我们提供了另一个被广泛引用的定义:“智力是一种非常普遍的心智能力,除了其他方面,它还涉及推理、计划、解决问题、抽象思维、理解复杂概念、快速学习和从经验中学习的能力。它不仅仅是书本知识、一种狭隘的学术技能或考试技巧。相反,它反映了一种更广泛和更深层次的理解我们周围环境的能力——‘理解’、‘弄清楚’事情的意义或‘搞清楚’该做什么”。更简洁地说,智力被描述为“快速而准确地解决问题” 。
认知能力差异形成了一个变异的层次结构。这源于发现所有认知测试都是正相关的;在一个认知测试中得分高的人往往在所有其他测试中也得分高,无论被评估的认知技能看起来有多么不同。自从 Charles Spearman 在 1904 年发现这一点以来,这一发现一直被不断复制。例如,John Carroll 重新分析了 20 世纪 400 多项研究中多种认知测试的相关矩阵 。这些研究包括了当时许多著名研究人员的研究,包括那些声称没有发现一般智力因子的人。Carroll 发现,在所有研究中,认知测试的分数都是正相关的,并且每项研究都包含一个通用成分,解释了人们表现差异的约 40%,有时更多。在认知领域层面(如记忆、推理和速度)也存在差异;即某些认知测试与某些测试的相关性更高——这些测试的内容与它们相似——而与其他测试的相关性较低。还有个别测试层面的差异。总之,如图 1 所示,人们在任何认知测试中表现良好的原因是:他们普遍智力高;他们擅长那种类型的测试;他们擅长测试中的具体技能;我们不应忘记测量误差,以及由于某种原因今天状态良好。
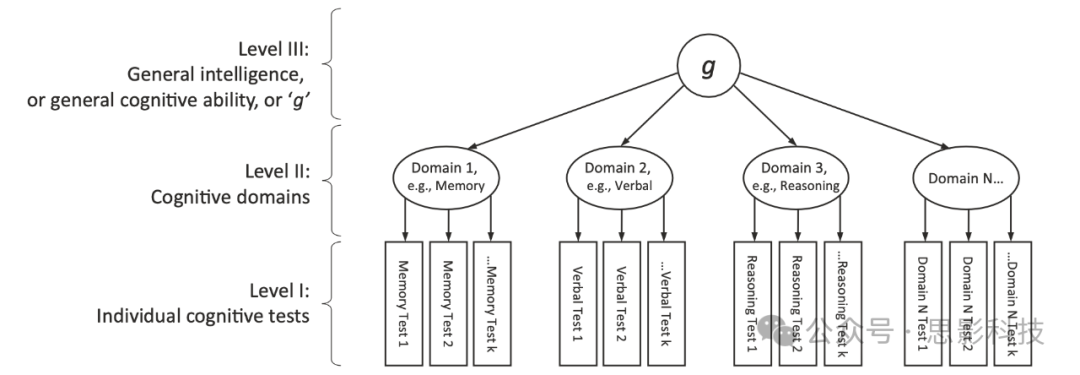
图1:人类智力差异的层次模型
在底层,第 1 层,个体在特定认知测试中的表现存在差异。这里显示使用多项测试(最多达到 k 个)来评估每个认知能力领域。所有认知能力测试的得分都呈正相关。然而,在考察同一认知领域(第 2 层)的测试之间存在特别强的相关性。
第 2 层显示了三个示例性认知领域:记忆、处理速度和语言。根据包含的具体测试类型,还可以有更多(最多达到 ‘N’ 个)。这些领域的名称是基于具体测试的显然共享内容而得出的常识性标签。某些测试可能会同时贡献给多个领域(图中未显示这种可能性)。个体差异在领域层面是可以观察到的。然而,人们在任何一个认知领域的得分与他们在其他领域的得分相关。这意味着有一个第三层描述所有领域的共同变异,因此跨越所有个别认知测试。这在第 3 层表示为一般认知能力、一般智力或简称‘g’。需要注意的是,这种三层结构是从数据中得出的,而不是强加于数据之上的。当对具有广泛认知能力范围的样本进行多样化认知测试时,g 通常占总测试得分变异的约 40%。
一般智力(g)
一般智力(g)作为一种统计现象,是来自不同认知测试电池(编者注:认知测试电池是指一系列用于评估个体认知功能的标准化测试)的普遍发现。然而,导致这种协方差的原因仍然是个谜。如果大样本的人进行两组不同的多样认知测试,这两组的g因子之间高度相关;因此,g不是特异的,其对个体的排名不会因测试电池的不同而显著变化。在可以从一组认知测试分数中提取的个体间差异来源中,g因子往往在与生活结果以及一些可能的起源(如脑影像指标和遗传变异)相关时居于首位。当研究人员测量一般智力时,他们通常使用以下指标之一:理想情况下,从多样认知测试电池中提取的第一个未旋转主成分或因子(编者注:未旋转主成分或因子(unrotated principal component or factor)是指在主成分分析(PCA)或因子分析(FA)过程中获得的最初计算出来的成分或因子,未经旋转处理。这些成分或因子解释了数据中最大的方差);涵盖多样认知技能的异质项目测试的总分;或一个具有较同质项目的测试,这些项目在多个认知测试的第一个未旋转主成分或一般因子上具有较高的载荷。
智力差异和平均分数的稳定性
一般智力的测量具有很高的测试-重测可靠性。以一个极端的例子为例,智力测试分数的稳定系数从11岁到约80岁之间在0.6到0.7之间,即使在纠正了测量误差之前也是如此。
另一种类型的稳定性——与年龄相关的平均值稳定性——显示出一个被很好复制的模式。涉及回忆学习信息的测试(称为晶体智力)——如词汇、常识和一些数字技能——在从年轻成人到老年期间平均水平相对稳定。涉及主动脑力工作的测试(流体智力的各个方面)在从年轻或中年成人到老年期间平均水平下降。这些包括认知领域如处理速度(例如在快速将数字编码成符号的测试中)、记忆(例如延迟回忆故事或单词列表,或工作记忆测试,如倒序数字跨度)、视觉空间能力(如在Wechsler积木设计测试中使用积木复制二维图案)和抽象推理(如Raven渐进矩阵中要求的从抽象图案进行归纳推理)。这些对年龄更敏感的认知领域往往协同衰老,其中一半或更多的个体差异的年龄相关下降可归因于一般流体智力的衰老。智力的老化存在个体差异,随着人口老龄化,这一问题的重要性日益增加;即,了解成功认知老化的决定因素(尤其是可修改的因素)非常重要。
智力的预测有效性:“健康、富有和聪明”
小学结束时的智力测试分数——大约在11岁——与几年后的教育结果高度相关,无论是测量16岁时标准化国家考试的分数(报告的相关系数高达0.8),还是接受的教育年限,或获得的最高学历。可能是智力导致了更长时间和更复杂的教育体验,反过来似乎也有一些小的影响。因此,智力和教育可能有一个动态的双向且可能是因果的关联。
智力是预测工作表现和学习表现最好的(也是最便宜的)指标之一,具有中等相关性。这适用于所有复杂程度的工作,尽管在更复杂的职业中相关性略高。童年时期的高智力与从父母(通常是父亲)职业地位向上流动中度相关。智力是英国社会经济地位差异的许多变量之一。父母较高的社会经济地位和更多的教育是其他独立贡献的变量;前者的影响相对较小,而且尚不确定教育在多大程度上作为先前智力的代理。
童年或青年时期测量的高智力与更长寿和更好健康之间存在稳健且一致的关联 。关于这个话题的研究包括一些非常令人印象深刻的样本,包括一个国家几乎全部的出生年份人口 和包含多达数百万受试者的样本。在早年智力较高的人,数十年后患上健康问题的可能性较小,并且从所有原因以及具体的心脏病、中风、呼吸系统疾病、吸烟相关癌症、消化系统疾病、痴呆、事故和自杀等原因死亡的风险也较低。这一领域的典型效应大小——认知流行病学——是青年时期智力高出一个标准差(15 IQ 分数)与这些疾病和死亡结果在数十年内降低20-25%的风险相关。用相关系数表示,童年认知测试分数与全因死亡率之间的关联通常在0.15到0.2之间。
因此,智力,通过认知测试分数操作化,有一个稳健表征的表型,高测试-重测稳定性,并且在教育、工作和健康方面有一定的预测效度;所有这些都为更广泛的构造效度做出了贡献。然而,在理解为什么有些人比其他人更聪明方面还存在不足。我们之前写过这些问题。2000年的一本书探讨了所谓“基本”认知成分、脑参数和遗传变异在智力差异中的可能起源。智力测试分数与一些看似简单的处理速度测量如反应时间和称为检查时间的心理物理程序之间存在稳健的关联。我们在这里不专注于这些,因为我们判断它们作为一些个体间智力差异的可能原因比遗传变异和脑结构和功能更难以捉摸。我们在2010年总结了智力测试分数与遗传学和脑成像的关联。然而,当时还没有智力的全基因组关联研究(GWAS),自那时以来,脑成像研究有了更大的样本、新的脑参数,并与分子遗传学研究联系起来。
智力差异与遗传变异
智力差异的遗传性和遗传结构
双胞胎和家族研究报告表明,遗传差异与智力测试分数的个体差异有关(见框2)。如果将所有年龄段的研究结合起来,遗传差异约占智力变异的50%(标准误差[SE]约为2%)。在成人样本中发现的遗传率估计值较高(可以达到70%或略高),而在儿童样本中,估计值较低(报告的估计值低至20-30%)。智力是可遗传的这一发现已在来自不同国家和时期的多个数据集中得到重复验证。本文重点介绍的是基于DNA的新研究结果,而不是传统的双胞胎和家族研究。
基于DNA的研究表明,遗传水平和表型水平都存在层次方差模式。使用基因组结构方程建模,发现一个遗传一般因子平均解释了58.4%(SE = 4.8%,对个别测试的解释范围为9%到95%)的七个认知测试中的遗传方差。这在一定程度上支持了这样的观点,即智力的表型结构部分是由于作用于智力一般因子和更具体认知水平的遗传效应。
自2011年以来,通过对大量无关个体的DNA直接测试,研究了智力的遗传性。这主要基于对称为单核苷酸多态性(SNPs)(见术语表)的遗传变异的测试。用于估计遗传性的统计遗传方法称为基于基因组的受限最大似然单成分(GREML-SC)(见框2)。这种方法测试了人们在认知测试分数上的相似性与其遗传相似性之间的关联,后者基于成千上万的SNPs。在此类研究中,遗传率估计约为20-30%(最近研究中的SEs < 1%)。使用GREML-SC发现的较低遗传率估计值可能是由于该技术更擅长捕捉与常见SNPs具有连锁不平衡(LD)的遗传变异方差,而不是那些较不常见或LD较低的变异。使用DNA测试和GREML-KIN分析方法在包含家族的大型队列中已经恢复了双胞胎研究和基于SNP的DNA研究之间的遗传率估计差异(见框2)。
| 术语表: 候选基因设计: 这种设计集中于特定基因中的变异,这些基因因为其生物功能和所选择的基因变异而被认为与感兴趣的性状相关。 遗传相关性: 这描述了两个表型之间共享的标准化平均遗传效应。它是每个性状的遗传可遗传元素之间的相关性。 全基因组关联研究(GWAS): 这种研究设计用于识别基因组中与某一性状或疾病状态相关的位点。GWAS可以包括数百万个来自基因组各处的常见单核苷酸多态性(SNP)。这些SNP分别被测试与感兴趣表型的关联性。为控制如此大量测试中的I型统计错误,通常采用严格的p值截断,一般为5 × 10⁻⁸。 遗传力: 这是表型变异中可归因于遗传因素变异的比例。在GWAS的背景下,通常只考虑与所用的常见SNP存在连锁不平衡(见下文)的变异中的加性遗传因素,因此遗传力估计是狭义遗传力。 连锁不平衡(LD): 这是指位于至少两个不同基因座的等位基因之间的非随机关联。 孟德尔随机化(MR): 孟德尔随机化是一种统计技术,使用从受精时起未改变的基因变异来测试暴露和结果之间可能的因果关系。MR通常使用已在全基因组范围内显著的暴露性状(如吸烟)SNP作为该暴露性状的代理变量,即形成其工具变量。这些工具变量由三个关键假设定义。首先,它们与暴露相关。其次,它们通过暴露而与结果相关。第三,它们与结果的混杂因素无关。 多效性: 这描述了多个表型可以与同一基因座的遗传变异相关的发现。首先,这可能是由于该基因座对每个表型有独立的因果效应,这被称为水平或生物多效性。其次,它可以描述一种表型与第二种表型之间的因果关联,意味着与第一种表型相关的任何遗传关联将与第二种表型共享;这被称为垂直或介导多效性。 单核苷酸多态性(SNP): SNP读作“snip”。这是人们之间最常见的遗传变异类型。每个SNP是基因组中特定位置的单个核苷酸替代。在GWAS中,SNP被视为关联的统计单位。 |
非加性遗传变异,包括显性效应和上位性效应,被认为是解释使用双胞胎和家族方法与使用DNA-SNPs得到的遗传率估计差异的部分原因。然而,一项研究发现显性效应与复杂性状的表型方差的关联不到4%。此外,数量遗传学理论预测上位性不太可能与大量表型方差有关。此外,通过使用高质量的插补面板得出的遗传率估计已经在无关个体中得到复制,这表明如果智力存在非加性遗传效应,它们并不是智力差异的主要贡献因素。
尽管GREML-KIN可能恢复一些归因于与常见基因型SNPs具有较差LD的遗传变异的遗传率,但它不能确定遗传率估计中归因于王朝效应的比例(编者注:在遗传学和环境影响研究中,“王朝效应”(dynastic effects)是指父母的基因型通过影响育儿环境来间接影响子女的表现,即使这些基因并没有直接传递给子女。这种效应反映了家庭中基因与环境交互作用的复杂性)。王朝效应包括父母基因型与后代表型之间的关联,包括通过未从父母传给后代的等位基因;这被称为遗传养育。虽然王朝效应的存在并不意味着捕捉间接遗传效应的方法存在偏差,但它确实阻碍了理解遗传差异如何引起表型差异的努力。这是因为所得的遗传率估计可能是直接遗传效应(一个生物体的遗传变异与同一个生物体的表型变异相关)和间接遗传效应(一个生物体的遗传赋予与另一个生物体的表型相关)的组合。
研究表明,教育存在王朝效应。在一项研究中,在移除间接遗传效应之前,其SNP基础的遗传率估计为29.2%(SE = 4.4%),而移除之后为17%(SE = 9.4%)。此外,通过发现预测教育的多基因评分(见下文)在非收养者中解释的表型方差是应用于收养者的多基因风险评分的两倍,建议环境影响对教育的遗传性有影响。多基因评分解释的方差既捕捉了直接遗传效应,也捕捉了通过环境中介的遗传效应。当在收养个体中使用多基因评分预测教育时,由与遗传相关的父母提供的抚养环境与后代的表型之间的联系被打破。由于智力与教育在遗传和表型上高度相关(见下文),因此间接遗传效应似乎也影响智力。
框2 智力的遗传性:为何不同方法会给出不同结果遗传性描述了在被测试的人群样本中,由遗传变异解释的表型变异的比例(通常以百分比表示)。需要注意的是,遗传性估计适用于特定时间的样本;在其他群体中,或在同一群体的其他时间,估计值可能会有所不同。通常,在人类研究中,只考虑加性遗传因素。不同的方法用于估计智力(及其他表型)的遗传性,它们给出的估计值不同。这些并不矛盾;相反,它们反映了不同方法可以获取的遗传变异来源,如下所述。 双胞胎和家庭方法双胞胎和家庭方法的遗传性估计使用参与者之间预期共享的等位基因比例作为样本中遗传变异的估计值。这些方法可以包括单卵双胞胎和双卵双胞胎之间的比较,以及包含父母、兄弟姐妹和其他关系的家庭研究。在每种情况下,如果遗传相似度更高的个体在智力测试分数上也更相似,则推断出遗传效应。对于智力,在双胞胎研究和家庭研究中发现了显著的(50%或更多,除非研究的是幼儿)遗传性估计。我们注意到,双胞胎研究假设双卵双胞胎与单卵双胞胎拥有同样相似的共享环境,这一潜在限制不会影响基于DNA的研究。 基于基因组的受限最大似然单成分(GREML-SC)GREML-SC,有时称为“GCTA方法”,是第一个用于推导智力遗传性估计的基于DNA的遗传方法。与双胞胎和家庭方法一样,遗传相似度与表型相似度进行比较。然而,遗传相似度是通过构建自基因分型的常见SNP的基因组关系矩阵来测量的,而不是推断的。重要的是,密切相关的个体被排除在分析之外(通常那些相似度超过0.025的人,即比二等表亲更亲近的个体)。这是为了确保家庭成员之间环境相似度不会被基因组关系矩阵捕获,从而导致遗传性估计的膨胀。使用GREML-SC的智力遗传性估计通常在20%到30%之间。GREML-SC的一个主要假设是遗传相似性与环境相似性不相关。虽然在某些情况下这一假设被证明不成立,但由此导致的遗传性估计的膨胀被认为是微不足道的。此外,GREML-SC假设一种无限小或多基因模型,即所研究的性状与大量变异相关,每个变异对表型方差的贡献极小。GREML-SC假设SNP效应呈正态分布,并且与LD(连锁不平衡)(见术语表)无关,并与次等位基因频率成反比。 基于基因组的受限最大似然亲缘(GREML-KIN)GREML-KIN引入了捕捉GREML-SC无法捕捉的稀有和不常见遗传变异效应的重要性。虽然它使用与GREML-SC相同的遗传数据,但GREML-KIN使用具有密集和已知谱系的样本,推导出额外的矩阵,以捕捉来自与常见基因分型SNP(单核苷酸多态性)具有较差LD(连锁不平衡)的遗传变异的额外方差来源,并控制环境影响的效应。当GREML-KIN应用于智力研究时,智力测试变异的54%得到了解释;因此,基于DNA的智力遗传性约与使用双胞胎方法得到的估计相同。GREML-KIN的假设与GREML-SC相同。此外,GREML-KIN使用密切相关的个体,且应用的数据必须包含足够密集的谱系,以防止由于这些密切相关个体之间的共享环境影响而导致的遗传性估计膨胀。 |
全基因组关联研究(GWAS)智力研究:寻找基因位点
遗传性分析表明某个性状受到遗传影响,但它们并不指出哪些遗传变异与性状变异有关。最初,候选基因设计(见术语表)被用来测试遗传变异与智力测试分数之间的关联。然而,这些设计缺乏统计功效,未能产生可重复的结果。唯一的例外是携带载脂蛋白E(APOE)基因的e4等位基因,这与老年时期略低的认知功能可靠地相关,占据了大约1%的变异。在解释智力差异方面,没有其他基于SNP的遗传变异接近这一效应大小。这种关联可能是因为APOE参与神经元修复,而在老年时期需要更多修复,并且神经退行性变的个体差异(变异)也更大。这个领域转向了GWAS(见术语表)的开展,这种方法对哪些(如果有的话)基因位点与感兴趣的性状有关持不知情态度。这是由覆盖整个基因组的数十万SNPs的经济实惠的芯片的可用性、大样本量的收集和多样本联盟的形成所推动的。
2011年,首次样本量超过3000的智力GWAS未检测到任何显著相关的基因位点。然而,它包括了第一个基于DNA(GREML-SC派生的)智力遗传性估计,并显示基因分型的SNPs确实在总体上解释了其变异。接下来的6年里,关于智力测试分数的GWAS大多未能识别出相关的基因位点。2018年,三项研究使用了显著重叠的样本,达到了超过200,000参与者的样本量,并发现了数百个与智力显著相关的基因位点。
这三项研究中的第一项样本量为248,428;它发现了187个(首次在线时有172个是新的)与智力相关的独立基因区域。这项研究的主要贡献来自于大型英国生物银行样本的短期言语和数字推理测试(VNR;被英国生物银行称为“流体”测试,这是一个用词不当)。该研究使用了元分析方法(MTAG)将使用不同认知能力指标的数据集结合起来,并包括教育成就以增加统计功效。然而,尽管以前使用替代表型方法识别了与教育和智力共同关联的SNPs,但MTAG中使用的元分析方法有所不同,因为它旨在检测与目标性状智力相关的遗传关联,而不是那些特定于教育成就的关联。
第二项研究样本量为300,046名参与者,发现了148个(首次在线时有53个是新的)与智力相关的基因位点。这项研究也使用了英国生物银行的VNR测试和其他几个样本,这些样本形成了三种或更多大多是流体智力类型测试的通用智力成分。第三项研究通过269,867名参与者确定了205个(首次在线时有84个是新的)基因位点。这项研究基于社会经济地位进行调整,结合了认知能力测试和学术能力测试的分数。
因此,很明显,智力差异的遗传贡献是高度多基因的,即有大量独立的遗传变异,每个变异解释的智力变异比例微乎其微。
上述三项研究使用多基因评分来基于DNA-SNP数据对智力进行样本外预测。多基因评分是基于SNP的效应等位基因总和的个体级预测因子,权重由描述每个SNP与性状(在本例中为智力)关联水平的回归系数决定。多基因评分在独立样本中预测了4-7%的智力变异;另一项研究预测了10.6%。因此,在这些样本中,出生时的血样预测智力的效应大小与父母的社会经济地位相当,即它们的预测效果不佳;对于预测个体智力而言没有实际用途。多基因评分解释的变异比例随着样本量的增加而增加,因此随着样本量的增加,其预测能力可能会提高。这引发了伦理问题——更多详细讨论见其他地方——这些问题应由相关兴趣群体和专业领域的知情专业人士和普通人共同解决。我们强调,上述结果适用于测试的样本,这些样本均为欧洲血统,并且相对较少来自非常贫困的情况。因此,这里报告的结果不应被假设适用于其他人群,或同一人群在其他时间的情况。
在智力的全基因组关联研究(GWAS)之后:对机制的线索?
找到与智力差异相关的基因位点变异只有在我们理解这些基因变异的机制后果时,才能帮助理解这些差异。GWAS数据集的智力研究结果发现了SNP变异与大脑皮层多个区域的组织特异性基因表达之间的关联(图2)。与智力相关的SNP变异已被链接到特定神经元类别的组织特异性基因表达,包括躯体感觉皮层的锥体神经元、海马CA1区、中脑胚胎期GABA能神经元和中型棘状神经元。这些关联表明,基因变异与智力变异之间的关联可能部分是通过皮层上基因表达的个体差异介导的,而不是特定的某个区域。
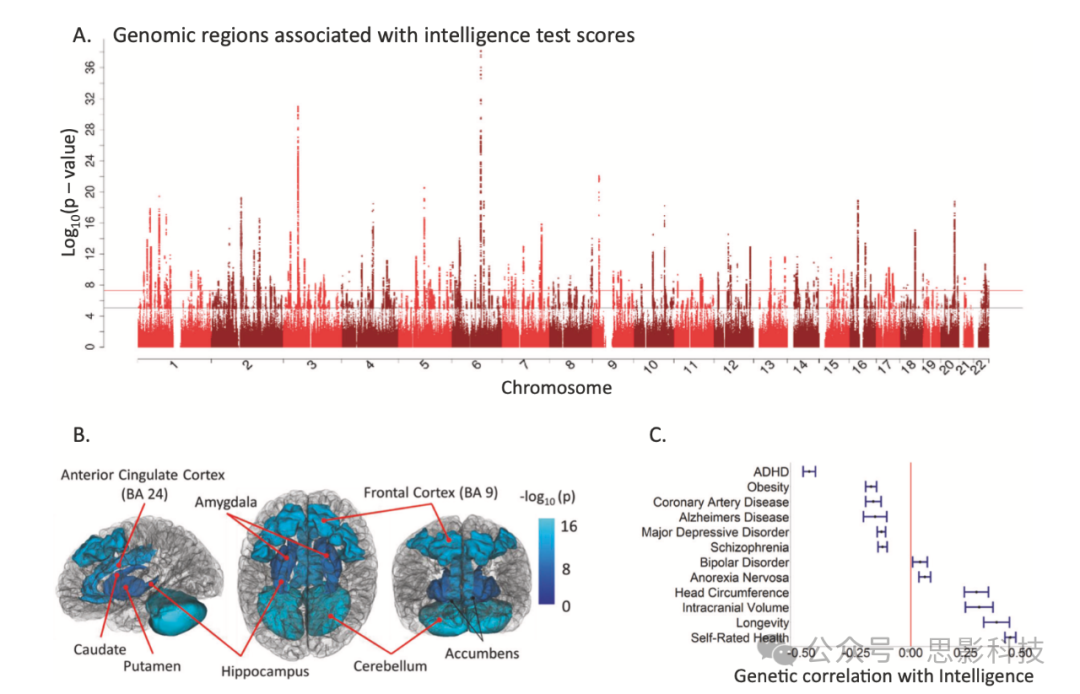
图2:显示了与智力测试分数相关的遗传位点,智力与大脑转录差异的重叠,以及智力与身体和心理健康的一些遗传关联。
A 显示了与智力测试分数相关的187个基因组区域的曼哈顿图。染色体在x轴上,-log10 P值在y轴上。每个点代表一个单核苷酸多态性。水平红线表示全基因组显著性阈值P=5×10⁻⁸,水平黑线表示全基因组建议变异阈值P=1×10⁻⁵。
B 大脑皮层中特定基因表达谱与智力基因关联之间的关系。这里展示的是在大脑中和跨大脑的组织特异性表达与智力之间的关联;完整的关联列表在原始报告中。
C 一些智力测试分数与健康、心理健康和大脑大小相关表型之间的遗传相关性。正遗传相关性表明,与较高智力测试分数相关的基因变异与特征值更高或发展障碍的可能性更大相关。长寿、颅内体积、自评健康和头围均显示出与智力的正遗传相关性。负遗传相关性描述了与较高智力相关的基因变异也与较低特征值或较低发展障碍的可能性相关的实例。注意缺陷多动障碍(ADHD)、肥胖、冠状动脉疾病、重度抑郁症和阿尔茨海默病等特征显示出与智力的负遗传相关性。
与智力测试分数相关的遗传变异往往集中在与神经发生、突触、神经元分化和少突胶质细胞分化相关的基因群中。这些结果与之前发现智力与大脑表达基因之间存在关联的研究一致,也与在突触后密度及其相关的NMDA受体信号传导复合体和Arc复合体中表达的基因有关。这些研究共同强调了突触,可能还有突触后密度及其相关成分作为生物系统的角色,当这些系统因常见的遗传变异而受到扰动时,与智力的某些表型差异有关。然而,在这些生物学上合理的组织中发现的与智力相关的SNPs解释的变异可能非常小。尽管如此,试图在神经元类型和基因系统等水平上理解DNA与智力表型的关联是驯服大量与认知相关的SNPs的方法,每个SNP的效应大小微乎其微。
多效性关联与智力
多效性(见术语表)描述了基因组区域的变异与多种表型相关的情况。通过导出两个表型之间的遗传相关性(见术语表)可以检测到两种表型之间的广泛多效性。遗传相关性描述了两个性状之间共享的平均遗传效应以及它是正的还是负的;即,当与一种性状增加相关的遗传效应也与第二种性状的增加相关时,发生正的遗传相关性,而当与一种性状增加相关的遗传效应也与第二种性状的减少相关时,发生负的遗传相关性。遗传相关系数是使用来自GWAS的所有SNPs导出的,无论这些SNPs与性状的关联水平如何。遗传相关性可以使用两个独立的样本导出;这有一个优势,即发现智力与疾病之间的任何遗传相关性不会由于个体患有疾病的症状而产生(即,如果智力GWAS是在健康个体中进行的)。多基因评分也可以通过导出智力的多基因评分并用其预测健康或脑成像性状来跨性状进行预测。然而,这些共享的遗传关联——通过遗传相关性或多基因评分检测到的——可能由于垂直多效性、水平多效性或虚假的多效性而产生,我们在下文解释。
使用GWAS数据集导出的遗传相关性表明,与较高智力测试分数相关的遗传变异平均也与更长寿命、更好的身体健康和更优越的社会经济地位相关。与较高智力相关的遗传变异更可能与较低水平的心理健康问题相关的性状相关(图2)。另一方面,与较高智力测试分数相关的遗传变异通常与自闭症谱系障碍和厌食症略有正相关。
孟德尔随机化(MR;见术语表)研究超越了智力与健康变量之间的关联,寻找一种表型可能与另一种表型存在因果关系的证据。MR结果表明,智力和教育可能有双向因果关系,并且智力可能与例如阿尔茨海默病存在某种因果关联,而这与教育的任何保护效应无关。然而,由于已知会影响教育的家族效应,智力与教育高度遗传相关,这种MR结果应谨慎解释。家族效应的存在违反了MR的独立性假设,因为它们在孩子成长的环境与其遗传继承之间引入了相关性。
选择性婚配,即根据与自己相似的遗传性状选择配偶的倾向,可能会偏倚MR的结果。教育和智力是有选择性婚配证据的性状;有报告称,配偶之间智力的相关性为r=0.40,教育的相关性为r=0.60。这些与个性测量的相关性形成对比,个性测量的相关性为r=0.10。由选择性婚配引起的偏倚可能是由跨性状选择性婚配引起的,例如,更高学历的女性可能会选择身高较高的伴侣,导致一个明显的“发现”,即身高与教育因果相关。可以通过在家庭内进行MR来控制由家族效应和选择性婚配引起的偏倚。
教育成就的GWAS显示与智力高度遗传相关(rg=0.70-0.80)并已确定了1271个独立的全基因组显著SNPs。这些数据的生物信息学分析表明,与大脑和其他皮质组织中表达的基因以及表达水平在产前和产后都升高的基因有关。此外,许多被识别的基因编码参与突触功能如突触可塑性和神经递质分泌的蛋白质,这与智力研究中确定的一致。然而,在遗传研究中使用教育作为智力的替代表型时应谨慎解释。例如,尽管与精神分裂症的遗传相关性表明,与较高智力测试分数相关的遗传变异也与较低的精神分裂症风险相关,但与获得更长和更高水平教育相关的遗传变异与更高的精神分裂症风险相关。在研究这一现象的研究中,与较低智力测试分数、较少教育和较高精神分裂症风险相关的SNPs也与早期发育过程相关。这与那些与较低教育和较低精神分裂症风险相关的SNPs的生物过程(成熟突触)形成对比。
对智力和教育与精神分裂症之间遗传相关性差异的另一种解释集中在“非认知技能”上。该研究使用基因组结构方程模型通过减法进行GWAS,结果在去除与智力差异有关的方差后发现了与教育特定的关联。所谓“非认知技能”的GWAS发现其遗传性为6.6%(SE=0.2%),与精神分裂症的遗传相关性为rg=0.26(SE=0.02)。然而,尽管捕捉教育的认知性状与精神分裂症有负遗传相关性,但“非认知性状”在遗传上与更大的精神分裂症风险相关。使用了一个单一的认知表现测试(英国生物银行的VNR)。这个简短的测试不太可能捕捉到与智力相关的所有方差,因此,这些未捕捉到的认知能力方差也会包含在“非认知技能”中(因此这个名称可能是误称),这表现在与其他智力测量的遗传相关性为rg=0.31(SE=0.06)。
智力与大脑
智力与大脑体积
大脑大小与智力测试分数之间存在一个重复验证的适度正相关关系。大脑大小通常通过磁共振成像(MRI)扫描评估为总体积。一项对超过8000人的148项研究数据进行的荟萃分析估计相关性为r=0.24。对这些数据重新分析,仅包括健康成年人后估计相关性为r=0.31;当仅包括被认为使用了质量更高的智力测试的研究时,这一值上升到r=0.39。在英国生物银行中对18426名中老年参与者(年龄范围44-81岁)的单一样本中,智力与总脑体积之间的相关性估计为r=0.276(95% CI=0.252, 0.300)。这大约在之前两个估计值之间,并且消除了影响荟萃分析结果的跨队列异质性。
除了总体脑体积外,还有许多其他方法可以探讨大脑差异,单靠总体脑体积并不能提供有关大脑复杂性的有用信息。例如,尽管大脑大小存在显著的性别差异,但智力的平均性别差异非常小或不存在,这可能是由于大脑结构、功能和连接的多方面补偿了任何明显的脑体积差异。值得注意的是,脑-智力相关性没有性别差异。有证据表明,与智力的关联程度因脑组织类型和位置而异。一项包括脑皮层特征(体积、面积和厚度)、皮层下结构总体积以及白质宏观和微观结构测量的研究发现,它们共同解释了73岁老人中多达18%的普通智力变异。在英国生物银行的更大年龄范围内,多个结构测量在老年参与者中的智力变异解释了更多(13.6%)的方差,而在中年参与者中则为5.4%,仅在前者组中超过了单一变量的总体脑体积。
顶叶-额叶整合理论(P-FIT)试图总结结构、功能和扩散成像研究中与智力相关的大脑区域(见图3)。它指出,外侧和内侧额叶、顶叶、外侧颞叶和外侧枕叶皮层的结构和功能变化以及底层白质连接(如弓状束)的变化与个体智力差异有关。当时可用的神经成像研究(37项研究)受到样本量小、智力测量方法各异以及当时扩散MRI(dMRI)研究数量有限的困扰,这可能导致了结果的相对较弱的趋同;即使是最强相关的区域也仅在≤60%的研究中得到支持。P-FIT理论得到了某些新的实证发现的支持(见图3)。我们将结合它考虑更多最近的研究。
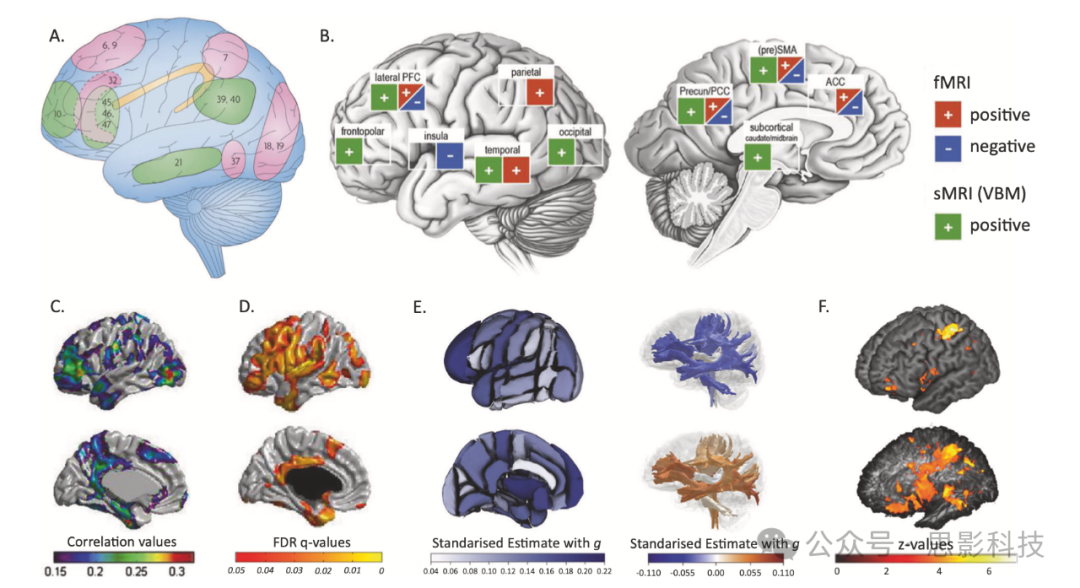
图3:人类智力差异可能的脑部位置
A 顶叶-额叶整合理论(P-FIT)解释的智力差异,由Jung和Haier提出。
B 一项智力功能和结构(仅体素基础形态学)研究的荟萃分析。
C 儿童(年龄范围6-18岁;N=216)皮层厚度与智力的关联。
D 老年人(73岁;N=588)皮层厚度与智力的关联;。
E 中年和老年人(年龄范围44-81岁;N范围=7201-18426)智力测试分数与区域皮层体积(左)、白质束分数各向异性(右上,蓝色)和平均弥散度(右下,橙色)的关联;。
F 皮层(上)和皮层下(下)病变位置与智力的关联(年龄M=49, SD=16年;N=241)。
框3 从大脑到智力理论的路径P-FIT(顶叶-额叶整合理论) 及其扩展我们在主文中提到了顶叶-额叶整合理论(P-FIT),作为将大脑测量与智力测试分数联系起来的多样研究结果的框架。在这里,我们描述其他不一定专门针对智力差异在大脑中位置的理论(如P-FIT),但它们对智力差异的神经科学研究仍具有重要意义。例如,对功能性大脑成像文献中几种不同传统的叙述综合确定了一些在多种执行功能任务中一致显示趋同激活的区域。命名法有所不同(如多需求网络【MDN】,前顶叶控制网络,上级认知控制网络,和外部模式网络),但这些模式通常包括背内侧和外侧前额皮层、岛叶和顶叶皮层。更近期的扩展MDN还包括壳核、丘脑和前额皮层的更多背外侧部分。尽管这些结果是基于多个任务中的趋同功能激活(而不是由多个任务指示的一般智力因素的激活模式),这些功能研究中涉及的区域与P-FIT有着惊人的相似性。整合结构和功能数据(以及其他模态)将是检验关于智力本质的更广泛理论的核心。这将为有关智力在大脑中是单一构造还是由多个重叠的脑网络支持的问题提供重要的生物学约束,这些网络支持从智力衍生的多种心理过程。它还将提供对皮层连接性由白质促进的假设的见解,这种连接性是处理速度个体差异的生物基质,处理速度在层级上从属于流体智力。 纵向和病变研究的贡献如果智力由大量分布的脑区及其连接支持,那么可能会有许多潜在的脑途径导致智力差异和智力变化。这是神经影像学可以提供重要见解的问题。病变研究和衰老过程的数据可以被视为有助于三角测量某些脑区是否对更高的智力是必要的,而不是由于混杂因素(例如衰老中的终生脑容量差异)相关。尽管如此,这两种方法仍然有限。 纵向研究比横断面研究提供了更严格的因果假设检验。例如,如果白质健康对更高的智力很重要,我们会预期两者的下降是相关的。然而,即使发现智力测试分数的变化与给定脑区或路径的变化相关,仍可能受到混杂因素的影响。一个不直接与智力功能相关的区域可能仍会表现出相关变化,因为其结构性下降仅仅与支持智力的区域相关。改进对纵向脑衰老的表征将阐明这种情况(区域脑变化相关的程度)对智力定位的影响。 病变研究为个体提供了宝贵的数据,这些个体在大脑中经历了突然且特定的局部损伤,可以与智力差异联系起来。那些与较低智力更强相关的大脑位置可以帮助绘制特定区域的重要性相对图。然而,病变研究可能受限于所需的数量,以消除大脑中统计力量变异性;即,样本中病变位置的区域覆盖可能是异质的。病变通常涉及灰质和白质是一个进一步的问题。在特定位置解析其对行为结果的相对重要性(并识别每种情况下受影响的底层连接路径)需要许多参与者具有选择性的(灰质或白质)组织损伤。 |
大脑灰质和智力
在成年人中,较高的智力测试分数与更大的大脑皮层体积和厚度相关,病变研究的数据也显示了P-FIT(顶叶-额叶整合理论)(见框3)中提到的区域有较强的关联,尽管效果大小始终较小。智力与皮层厚度的关系在生命周期中发生变化,在10岁儿童中报告了负相关。在这些最近的成人研究中(N = 18,426,年龄范围44-81岁),智力与岛叶、后扣带/楔前叶体积之间的一些最强关联也被发现(r < 0.20)。这些区域在最初的P-FIT中没有被提及,但在使用功能和结构MRI数据的元分析更新中被确定。然而,仅考虑结构体素为基础的形态学分析在容易进行元分析的常见空间中意味着排除了其他类型的结构研究。基于UK Biobank的大型研究发现,智力与丘脑体积(r = 0.25)的关联比任何其他皮层下结构都要大1.5倍(r = 0.06–0.17)。以前也在较小样本中发现过与智力差异相关的尾状核在UK Biobank分析中也与智力相关(r ≈ 0.13),但在包括所有皮层下结构的多变量模型中,只有海马(r = 0.05)和丘脑体积(r = 0.19)显示出与智力的独特关联。这些发现与岛叶和丘脑的广泛皮层连接模式一致。
大脑白质和智力
过去20年,由于扩散磁共振成像(dMRI)的出现,白质大脑成像研究激增。通过利用各种白质属性(如轴突髓鞘化和直径等)对水分子扩散运动的影响,dMRI能够推断出白质的微观结构。常用的测量指标包括各向异性分数(FA,扩散方向一致性的指标)和平均扩散率(MD,扩散的平均强度)。两者都显示出一般和局部的衰老效应:平均而言,FA随年龄增长而下降,MD随年龄增长而上升。
当作为白质多个部分的汇总测量时,更方向一致的水分子扩散(更高的FA)和更低的总体扩散强度(更低的MD)与智力测试分数较高相关,这些研究涵盖了8至81岁的年龄段,通常效果大小较小。在老年人中,FA和流体认知能力在三年内的显著耦合下降(r = 0.31【94】)。尽管我们承认白质束命名和识别上的不一致,但有证据表明,长程皮层-皮层(关联通路)、胼胝体膝(比胼胝体压部更明显)和皮层下-皮层(主要是丘脑)通路显示出与智力关联较大的数字(尽管所有r < 0.11)。这些通路的识别与上述灰质发现一致,因为这些通路大致是促进额叶、顶叶、外侧颞叶(和皮层下)互连的连接。白质损伤的年龄相关累积(通过白质高信号【WMHs】测量)与较低的认知功能(r = -0.106)和纵向耦合下降(r ≤ -0.334)相关。WMHs可能干扰选择性通路,因为它们倾向于主要积累在侧脑室上方。
多种白质微观结构测量的综合可以找到更大的脑智力关联效果大小。一项研究使用三个基于扩散的指标(FA、纵向弛豫时间和磁化转移率)在12条白质束中得出白质完整性的一般因子。这三个一般因子彼此之间的相关性很小,表明它们可能捕捉到白质微观结构的非重叠方面。总体上,它们解释了73岁老年人智力差异的约10%。这种白质完整性与智力之间的联系完全由处理速度的认知域介导。
大脑区域和它们的底层连接与我们最复杂的认知能力的个体差异相关,它们也可能:(i)随着大脑体积的增加而显示出更大的面积扩展;(ii)是那些最晚发育的区域;(iii)是那些对脑老化及其潜在决定因素(如血管风险)最敏感的区域。重要的是,即使使用大脑区域测量,效果大小仍然相对较低,在样本充足的研究中上限约为~r = 0.30;然而,这些估计在大样本中是稳健和可复制的。这些发现说明了多种大脑结构特征与智力差异的小但显著关联。这在老年人中尤为明显,因为这些测量中的较大部分变异可能是由年龄相关的神经退行性过程差异驱动的。考虑到大脑变量的宏观规模以及上面讨论的测量只是可能测量的大脑特性中的一小部分,适中的关联并不令人惊讶。
新兴的大脑与智力关联方法
新的方法开始对不同灰质区域和白质路径对智力的多变量跨组织贡献进行建模。例如,少突胶质纤维束小纤维和前极体积的FA介导了18.2%的年龄与流体智力的关联。这些方法有潜力更直接地测试基于大脑网络的智力理论的特异性,例如P-FIT。利用大脑的结构连接组提供了比通过测量更少、更大的路径更高保真度地评估此类基于网络的分析的机会(尽管这带来了不同的一组限制)。全局测量如连接组效率,或形态学相似网络中节点的“度”变异,已显示出预测智力差异的潜力(在一项对296名年轻成年人的研究中高达40%)。静息态fMRI连接矩阵预测了年轻成年人中20%的智力差异(N = 884)。这些结果需要复制。因此,继续开发结构和功能大脑连接组的清晰且可解释的整合是很有意义的,以增强我们对智力差异与大脑的理解。
下一阶段的研究应尝试解释大脑指标(有时相当粗略)与智力之间的关联,并寻找和测试新的大脑变量。拥有更大体积的大脑(及其中的组织)似乎与拥有更多的神经元有相对强的关联。皮层厚度差异与神经元密度、柱状排列、树突树冠和胶质细胞特性相关。另一个有前景的指标可能是通过神经纤维方向分散和密度成像(NODDI)评估的灰质扩散特征,这些是树突密度和分支的假定标志,并展示了一些类似P-FIT的区域与智力的关联(|r | <0.25)。然而,在我们理解这些具体的微观特征(其中一些是通过神经影像估计的)是否以及如何具体导致智力差异之前,还需要做大量的工作。虽然新方法、更大的样本量和样本外预测设计有可能为智力变异提供更多见解和补充贡献,但我们认为重要的是:(i)测试较新测量方法在传统指标之外的增量效度;(ii)批判性地评估这些较新测量方法是否提供了更多生物学上可处理的变量,以理解大脑差异;如果是这样,(iii)将这些新指标与其他解释层次整合起来,跨越从宏观结构到细胞的范围。
向更好地整合基因、大脑和智力迈进
我们将这篇综述分为关于智力的表型问题、智力测试分数的基因关联及其大脑相关性的部分。这在一定程度上反映了作者之间的分工,但也反映了当前这些研究方向的普遍分离。我们希望能有更多融合的大脑-认知-遗传研究,并希望能有更多综合性的部分、示例和观点可供回顾。然而,我们认为这对我们和整个领域来说是一个教训,关于可以进行的研究类型。这些划分部分也是因为智力的统计表型性质的研究已经进行了一个多世纪。遗传学和大脑成像相对来说还很年轻,而且方法学上发展迅速。这些领域中涌现了许多新方法,关于这些方法的优劣讨论和探讨将继续快速进行。举几个例子,关于扩散成像和功能磁共振成像工作在跨实验室稳定性方面的讨论,以及关于智力与其他表型之间观察到的基因相关性的因果性质,以及非传递性遗传效应的讨论。智力的表型性质也存在争议:神经科学和认知文献中的一些领域侧重于评估所谓执行功能方面的实验认知任务,而这些任务与智力分数所基于的心理测验形成对比。因此,仅关注例如执行功能或工作记忆的综述可能无法完全体现其与心理测验智力的强表型,基因型和神经结构(见第3框)重叠的情况。开发一个理论上的超结构来统一这些认知学科将是有趣的;遗憾的是,认知神经心理学家和心理测量学家在实际强相关的情况下没有更多地将他们的测试和构念结合在一起。在这一部分中,我们总结了一些最近结合基因和大脑以尝试理解智力差异的研究,然后讨论其影响和未来可能的方向,以期为智力差异的一些生物学基础提供更综合和细致的解释。
综合考察基因、大脑和智力
基因相关性发现,与智力相关的遗传变异部分与大脑结构的体积测量相关,例如颅内体积(rg = 0.27)、总脑体积(rg = 0.23)、灰质体积(rg = 0.08)、白质体积(rg = 0.08),以及左后扣带回皮层的体积(rg = 0.23)。更健康的大脑白质微结构与更高的智力之间存在正向的基因相关性。反应时间——较高的数值表示较差的反应——与大脑白质健康/完整性的基因相关性平均为−0.18,基于FA,以及平均为0.17的轴向扩散、MD、各向异性模式和径向扩散的广泛正向基因相关性。
一项研究发现,智力的多基因评分在新样本中预测了3-5%的智力差异,并报告了这种关联部分通过前扣带皮层、前额皮层、岛叶和内侧颞叶皮层的脑皮层厚度和表面积小范围地中介了。结果一致地表明,与智力测试分数相关的遗传变异部分可能通过其在已与智力差异表型上联系起来的多个区域中大脑结构的变异来解释。
智力和大脑结构测量的高度多基因性可能表明大量生物系统与两者的个体差异有关。这可能成为揭示遗传变异如何与大脑和智力差异相关的机制性解释的障碍,因为与两者相关的任何生物系统可能仅能解释大脑和智力测量中表型差异的一小部分。然而,尽管每种机制分别可能与小效应量相关,但未来的工作应致力于产生更机制化的智力差异解释,应该检验任何已识别生物系统的相对重要性,并检验其与大脑成像测量的关联。
智力和皮层测量之间确定的多效性也需要进一步研究。具体来说,垂直多效性和水平多效性在生成认知能力和大脑变量之间的基因相关性中的相对重要性目前尚不清楚。通过理解导致这些基因相关性的多效性形式,可以制定出更多机制化的智力差异解释。例如,如果垂直多效性驱动智力和大脑结构之间的基因相关性,那么这可能表明大脑结构在智力差异中是因果性的;然而也可能存在相反的情况,即智力部分驱动大脑差异。如果这些基因相关性是水平多效性的结果,这将表明相同的遗传位点与大脑结构和智力相关,而两者之间没有因果关系。
解释智力与大脑形态学之间基因相关性的相关困难来自于非传递性遗传效应。在这里,智力与皮层测量之间的基因相关性部分可能是由于父母的基因型与培养实践相关,这些实践支持健康的大脑发育和智力增长。家族内部的GWAS将为未来识别这些效应提供机会,并在没有父母基因型对这些变量的影响的情况下评估智力与大脑结构之间剩余的基因关系的大小。
基因g因子的发现也对与智力相关的遗传位点的功能注释有影响。具体来说,与认知能力相关的位点可能与一般因素的变异相关,也可能是与所研究领域特定的或具体测试相关的关联。当研究以前与Trails B认知测试相关的基因位点时,可以清楚地看到这一点;研究表明,Trails B的分数与基因位点的关联与其他认知能力测试共享,因此比最初假设的更为普遍。这对于研究智力差异的更多机制化解释来说是一个潜在问题,因为所有认知测试中的共同变异和特定测试领域或单个测试的变异可能被包含在一起。通过使用基因组SEM等技术将这些关联提炼为跨认知领域的一般关联,以及特定于每个领域或测试的关联,可能有助于识别与每个领域相关的可行生物机制。
超越基因组学
迄今为止,直接整合基因变异、大脑成像指标和智力测试分数的研究还很少。我们甚至还没有对如何将被确定为与智力测试分数和大脑指标相关的大量基因变异直接联系到机制(如蛋白质表达)以及这些机制如何促进神经生物学机制的属性的理解。这种解释差距是巨大的,可能会引发一系列反应,从布尔克崇高观念中的愉悦和恐惧(尽管任务艰巨,但有待完成),到更不理想的过早和简单的还原论或绝望情绪。
有一些领域可能有助于弥合这一差距,并有助于理解智力差异,例如表观遗传学、转录组学、蛋白质组学、虚拟组织学等。与例如GWAS方法相比,这些领域还较新,需要时间来填补基因、大脑和智力三者之间的空白。然而,已经开始出现一些关于智力和大脑结构的全基因组甲基化研究结果。一项研究发现染色体10上的cg12507869在INPP5A基因中与认知能力(使用简短精神状态检查量表测量)以及语音流畅性有关。INPP5A基因中的甲基化探针也显示与海马体积有关,尽管在对多个大脑区域进行校正后这一关联未能经受住检验。尽管如此,我们认为多重表观遗传学方法(例如EWAS及其产生的表观遗传分数在样本外创建)和更大的样本量,特别是在表观遗传学-神经成像研究中,将进一步推动进展。例如,吸烟、死亡率和炎症的表观遗传特征在老年人中表现出与P-FIT(顶叶-额叶整合理论)区域重叠的认知关联和区域皮层相关性。
“虚拟组织学”方法——利用区域基因表达的组织学数据——也有潜力理解基因和大脑测量在认知差异背景下如何相互关联。统一特定空间层次(例如皮层分区)的基因表达模式和大脑结构信息,使研究人员能够询问大脑结构-表达关联的区域差异是否与大脑结构的其他相关性有关,使用相关性的相关性方法。例如,那些显示出较强的三酰基甘油表达与皮层厚度关联的大脑皮层区域,也是皮层厚度与一般认知能力关联更强的区域。多变量和系统特定的方法(考虑多种蛋白质)将进一步阐明特定蛋白质的独特贡献及其可能与认知差异的神经生物学基础相关的潜在机制。
最后,我们想明确关于使用基因或大脑成像变量“预测”智力的问题。我们倡导在样本间进行重复测试;也就是说,如果在发现样本中一组基因变异或一个或多个大脑成像变量与心理测量智力相关,那么在重复样本中测试关联是否成立是明智的。这是一种验证和理解从起始样本到智力的基因和大脑相关性的普遍性的方法。因此,通过样本外预测到另一个样本的重复是我们可以增强对人们认知能力差异原因理解的工具。另一方面,从基因或神经成像变量预测个体智力并不是一个实际的或我们认为理想的目标。
结论
在过去的10年中,大脑成像和基因与智力测试分数差异的关联取得了进展,基于新方法和大样本获得了一系列结果。成像和基因变量解释了智力变异的一小部分。我们在这两个领域的结论是:应寻找其他变异来源;我们与哪怕是部分机制解释为何人们在智力上存在差异之间仍有很大的解释差距;这些关联不应被视为对智力有不可改变的贡献。当我们理解这些及未来的关联时,可能会有一些关于什么倾向于最佳认知发展和健康认知衰老的提示。我们认可并鼓励对智力其他重要变异来源的研究,包括社会和生物因素。

















 896
896

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









