







本文是昆士兰大学化学与分子生物科学学院的Yosephin Gumulya等人发表在Nature Catalysis上的名为“Engineering highly functional thermostable proteins using ancestral sequence reconstruction ”的文章,本文通过祖先序列重建技术成功开发了耐热性和活性显著提升的酶,这些酶为生物催化、合成生物学及工业应用提供了更加稳健的起点。
摘要
在21世纪新兴的生物经济中,酶作为生物催化剂在可持续的绿色生产领域发挥着关键作用,涵盖精细化学品和生物燃料的生产。然而,与传统化学催化剂相比,酶的应用受到稳定性差和生产成本高的限制。同时,耐热蛋白在合成生物学中的应用非常受欢迎,因为它们更易于结晶化以及进行其他生物物理特征化分析。
本文通过重建祖先酶,展示了其在温度提升约30°C和延长≥100倍的孵育时间下依然具有高度稳定性。这一研究以动物细胞色素P450酶和细菌酮酸还原异构酶(KARI)为模型系统,分别应用于合成高价值化学品和丁醇基生物燃料的生产。研究发现,重建的脊椎动物CYP3 P450祖先酶在溶剂耐受性方面优于人类代谢酶CYP3A4,同时表现出相似的广谱底物活性。重建的祖先KARI酶在25°C时比对应的大肠杆菌形式具有高达8倍的比活性,并在50°C时进一步提高3.5倍。因此,仅利用序列数据即可设计出具有显著热稳定性的蛋白,即便是近期的祖先形式。
引言
在21世纪新兴的生物经济中,酶因其作为生物催化剂的作用而在可持续性和绿色生产中占据重要地位,包括用于生产精细化学品和生物燃料。然而,当前酶的应用受到其稳定性较差和与传统化学催化剂相比较高的生产成本的限制。此外,耐热蛋白在合成生物学中的应用尤其重要,因为它们不仅更易于结晶化,还更适合用于其他生物物理特性分析过程。
尽管已经尝试对酶进行工程化以提高其耐热性和溶剂耐受性,但目前尚未建立一种通用的策略来设计具有显著稳定性提高的蛋白。对大多数蛋白来说,关于序列、结构和功能之间关系的信息不足,这限制了高效的理性设计,尤其是当需要结合个体稳定性突变时,很难预测可能发生的上位效应(epistatic effects)。尽管计算方法有所应用,但这些方法依赖于结构数据,且通常需要进行耗时和资源密集型的研究来提供训练数据。祖先序列重建研究表明,与现存形式相比,前寒武纪时代的酶可能具有更高的热稳定性。常见的改进方法是结合定向进化和靶向理性设计、共识序列或祖先突变,这些方法被用来选择具有更高热稳定性、溶剂耐受性和其他期望特性的酶,例如底物特异性和催化效率。然而,这种方法通常需要多轮突变和筛选大量突变体,来逐步实现性能的改进。
以CYP102A1为例,这是一种细胞色素P450(P450)家族的代表性酶,能够对未活化的C–H键进行区域和立体选择性功能化,这在高价值化学品的生产中是一项极具挑战性且高度需求的反应。CYP102A1已经成为开发多种定向进化方法的模型系统。为了开发一种耐热的CYP102A1形式,研究需要通过多轮点突变来使蛋白的变性温度较原始形式提高约6°C。通过重组定向进化,又将温度进一步提高了3.4°C。随后,利用高斯过程景观建模结合重组CYP102文库的热稳定性数据筛选,进一步实现了约5.3°C的提高。
重组进化已被证明能够生成具有多样且常常是新颖特性的突变体文库,探索了比随机突变或位点饱和突变方法更广泛的序列空间。然而,当自然进化过程中在不同进化分支中出现的结构上不兼容的残基在体外重组时,所得突变体的适应性(包括稳定性和活性)可能会受到影响。我们假设,即使是相对较新的嗜中温祖先,其蛋白骨架与后代的不同变异更兼容,因此比现存形式更适合作为重组定向进化的起点,用于开发更稳定的酶。
本文以P450酶和酮酸还原异构酶(KARI)两种与生物催化相关的不相关酶类作为模型系统,验证上述假设。
结果
1. 嗜外源性代谢酶的耐热祖先形式
我们聚焦于现存P450家族中底物特异性最宽的一个家族,并推测及重建了CYP3家族的祖先(图1a),称为CYP3_N1。该祖先酶被预测为存在于约4.5亿年前的第一批脊椎动物中。表达结果显示,CYP3_N1能够在大肠杆菌中以血红素蛋白的形式表达,并具有典型的Fe(II)·CO与Fe(II)差异光谱,表明其正确折叠。表达水平超过500 nmol l⁻¹(约25 mg l⁻¹)。出乎意料的是,在未经过任何定向进化之前,CYP3_N1已表现出较现存CYP3A(人类CYP3A4、CYP3A5,以及鱼类CYP3A27和鸟类CYP3A37)更高的内在耐热性。其60分钟折叠保留率(60T₅₀,即60分钟内仍保持50%折叠状态的温度)为66±2°C,比现存CYP3家族成员高出约30°C(图1b)。此外,CYP3_N1的最佳反应温度也较CYP3A4提高了约20°C(图1c)。
生物催化工艺的经济可行性依赖于最大化酶反应的体积-时间产量;稳定性提升至更高温度有助于延长在温和工艺条件下的半衰期。在37°C下,通过光谱检测未观察到CYP3_N1的任何活性损失,持续数日。然而,在50°C和60°C条件下,与CYP3A4和其他现存CYP3形式相比,CYP3_N1的半衰期从1-5分钟延长至约10小时(图1d)。从45°C到80°C,祖先和现存CYP3酶均表现出一级失活动力学,Arrhenius图显示CYP3_N1的失活能量为251 kJ mol⁻¹,是现存P450(91-151 kJ mol⁻¹)的1.7至3倍。
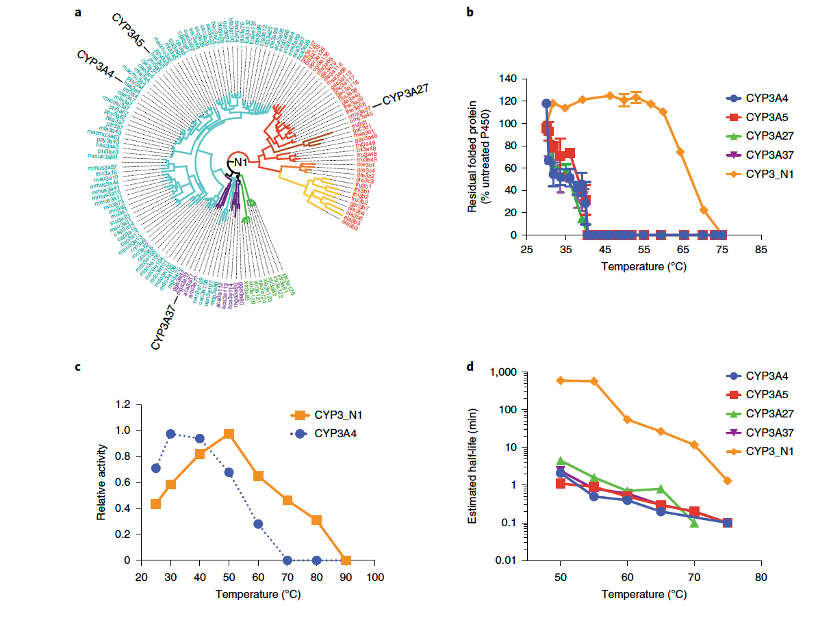 图1
图1
2. CYP3_N1的催化活性与人类CYP3A4相当
与人类CYP3A4(已充分表征)进行对比,CYP3_N1对已知的CYP3A4底物表现出相当甚至更高的活性(图2a)。通过光谱结合研究,观察到CYP3_N1与多种结构不同的配体(包括几种类固醇和大型大环内酯类抗生素)相互作用(图2b),其亲和力与CYP3A4相当。在6β-羟化睾酮和硝苯地平氧化反应中,CYP3_N1表现出更高的催化效率(补充图3和补充表5)。因此,CYP3_N1的底物多样性与人类主要药物代谢P450相似,尽管在某些底物上表现出相对活性(包括区域选择性)的变化。
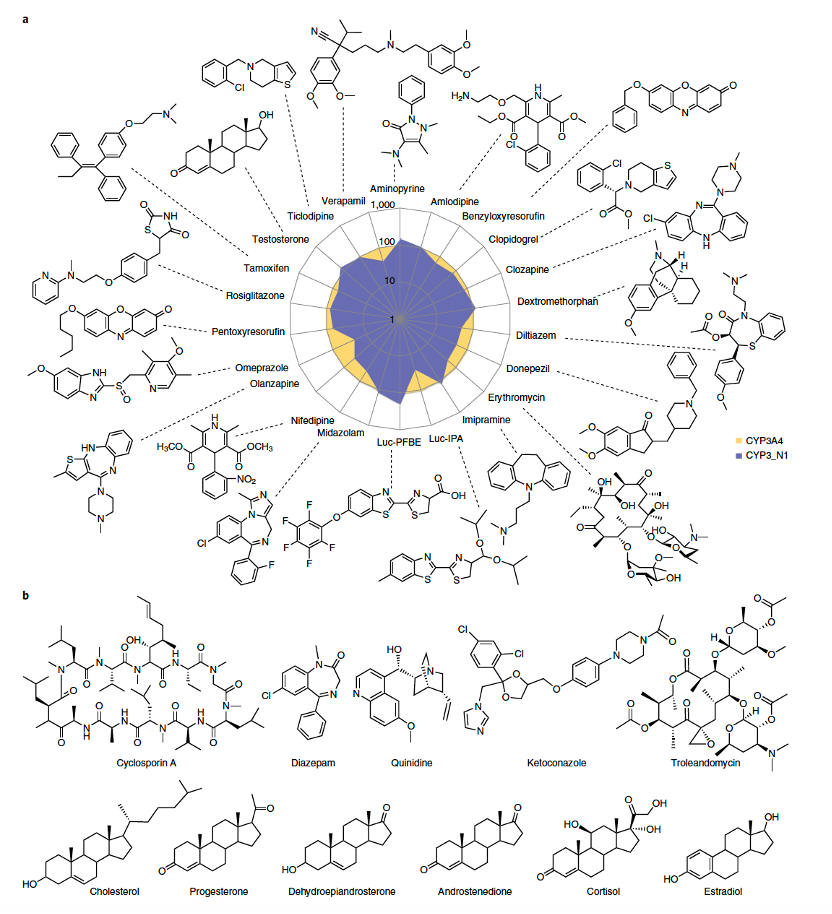 图2
图2
3. CYP3_N1通过疏水相互作用实现稳定性提升
与现存CYP形式相比,CYP3_N1在氨基酸序列上有139-169个残基差异。尽管整体氨基酸组成没有显著趋势,但CYP3_N1中不同疏水残基的比例略有变化。因此,我们假设其增强的耐热性来自于更好的疏水核包装。在CYP3_N1中,有9个替换位点被认为提高了特定折叠区域的非共价相互作用。当这些位点的残基疏水性被降低时,每个突变都会显著减少热稳定性;当9个位点同时改变时(突变体SDM8),10分钟折叠保留率(10T₅₀)下降至与CYP3A4无显著差异。然而,将逆向突变引入CYP3A4并未提高其热稳定性,甚至在某些情况下阻碍了突变体的异源表达。这一结果突显了在不同的上位效应背景下,通过理性设计实现蛋白耐热性的困难。
4. CYP3_N1是进一步定向进化的理想模板
在祖先序列重建(ASR)中,无法确定历史上确切存在的序列,而只能推测最可能的祖先。因此,为了验证CYP3家族祖先酶耐热性的推断稳健性,我们在推测不确定性最大的10个位点引入突变(图3a、3b)。这些位点的不确定性体现在推测的祖先中两种最可能残基的后验概率非常接近。实际上,我们生成了一组可能的祖先蛋白,这些蛋白可能在早期脊椎动物中存在(称为CYP3祖先云库,CYP3 ancestral cloud library)。这一祖先云库不仅代表了一组可能的祖先形式,也提供了一组突变体,其表现出不同的表型,可供定向进化实验选择最适合的起始模板。对于生物技术应用而言,表型的多样性比稳健性更重要,因为多样的表型有助于蛋白工程。
为了设计这一云库突变体,编码增强型黄色荧光蛋白(eYFP)的C-端融合标签被引入。在所采样的1,023个突变体中,77%成功表达,细胞显示eYFP荧光。这些表达的突变体在大肠杆菌中均检测到可检测水平的P450,平均产量为550±270 nmol l⁻¹(平均值±标准偏差)。虽然大多数突变体显示出与CYP3_N1相当的耐热性,但222个突变体表现出显著更高的耐热性,其10T₅₀高达75.8°C。筛选一部分突变体发现,其对典型CYP3A4底物的活性表现出多样化的特性,表明该库功能多样。
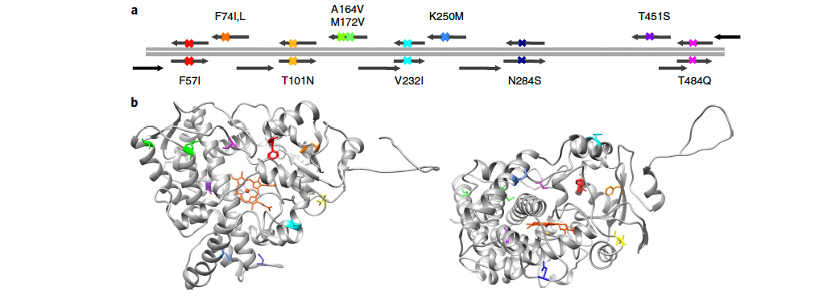 图3
图3
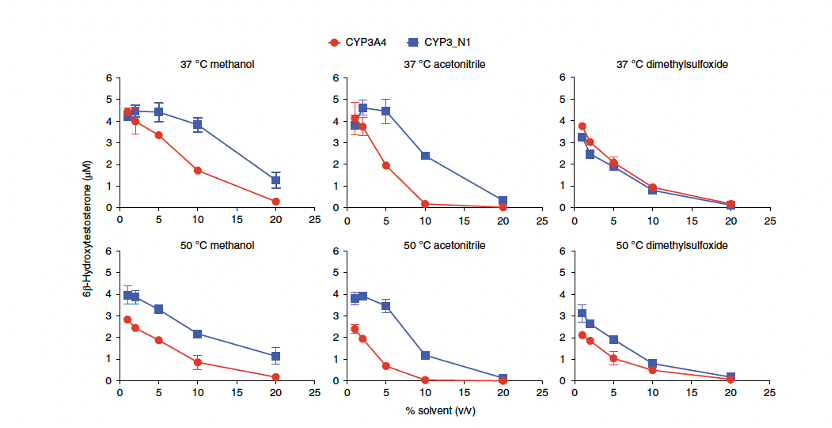 图4
图4
5. 祖先CYP3酶表现出增强的溶剂耐受性
酶催化的经济性还依赖于底物负载能力的提升。对于许多药物等亲脂性底物,在有机溶剂存在下进行反应可能更有利。为了测试溶剂对CYP3_N1的影响,并排除人源NADPH-细胞色素P450还原酶(hCPR)的干扰作用,实验使用氧代替物cumene过氧化氢来支持反应。结果显示,与CYP3A4相比,CYP3_N1对甲醇和乙腈的耐受性显著增强,但对二甲基亚砜无明显提升(图4)。在37°C下,CYP3_N1在10%(v/v)甲醇和乙腈中分别保持91±7%和63±6%的活性,而CYP3A4仅分别保持39±2%和4±2%的活性。
6. 该方法在其他蛋白家族中的适用性
为了验证这一方法的通用性,我们进一步研究了一种功能上无关的酶——酮酸还原异构酶(KARI)。KARI将2-乙酰乳酸转化为2,3-二羟基异戊酸,或将2-乙酰-2-羟基丁酸转化为2,3-二羟基-3-甲基戊酸(EC 1.1.1.86)。数据库中大多数II类KARI序列来自于真正细菌(主要是变形菌门和拟杆菌门),其余少数来自疣微菌门、纤维杆菌门和螺旋体门。因此,推测分析了这些群体的共同祖先(图5a)。通过差示扫描荧光法测得祖先II类KARI的熔点温度(Tm)为59°C,相较于现存的大肠杆菌和水稻II类KARI(Tm分别为42°C和44°C)显著提升(图5b)。此外,该祖先酶对2-乙酰乳酸的比活性高出8倍,尽管其米氏常数(Km)比现存大肠杆菌形式高出5倍(图5c)。此外,该祖先酶的10分钟折叠保留率(10T₅₀)也有所提升,尽管其温度最适值和异丁醇耐受性并未优于大肠杆菌形式。
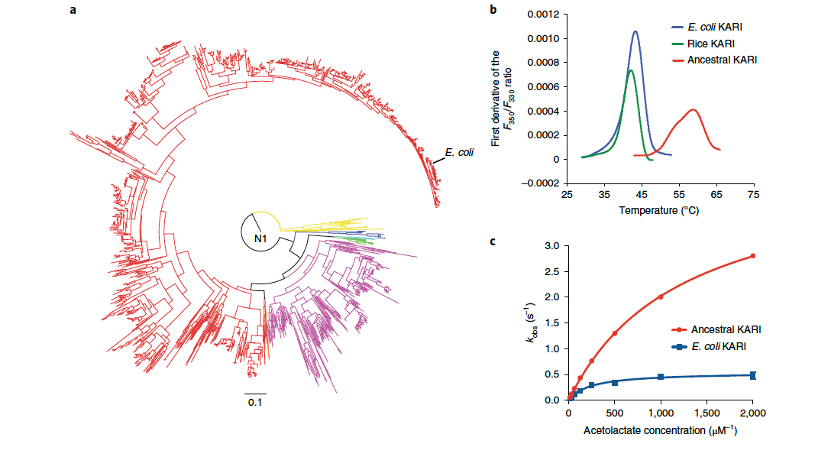 图5
图5
讨论
热稳定性是商业用途蛋白的关键特性。在生物催化中,热稳定性可以延长酶的使用寿命,从而降低成本并提高产量。在更高温度下运行反应可以增加底物负载,减少生物反应器中微生物污染的可能性,从而提高工艺的经济性。即使T₅₀值(在特定温度下50%蛋白保持折叠状态的时间)仅提高5-10°C,也对工业应用具有重要意义。
本研究通过对CYP3_N1祖先酶的重建,取得了约30°C的耐热性提升。这一结果可与针对CYP102A1(CYP3家族中的另一成员)的耐热性工程研究成果相比较。CYP102A1是过去二十年中工业应用中广泛研究的酶,其最稳定的突变体表现出10T₅₀为69.7°C,而CYP3_N1的10T₅₀为71.4°C。需要注意的是,CYP102A1突变体源自具有较高初始Tm值(10T₅₀为49–55°C)的现存蛋白,而本研究中的CYP3A现存形式初始10T₅₀值仅为35–38°C。
现有方法与祖先序列重建的比较
传统的蛋白稳定性增强方法通常包括随机、定点和重组突变的定向进化。这些方法无需结构信息即可取得改进,但通常需要多轮突变和筛选,且需大量时间和资源投入以获得逐步提高的突变体。理性突变和计算设计也取得了一些成功,但需要对蛋白结构-功能关系有深刻的理解。此外,高斯过程景观建模在CYP102的研究中取得成功,但依赖于以往研究生成的训练数据。
相比之下,本研究利用祖先序列重建,仅需一次推测操作即可得出祖先形式,无需任何先验的结构信息或热稳定性数据。通过分析目标蛋白(或亚家族)的大量序列,本方法可应用于任何已知多个序列并被测序的蛋白家族。
祖先序列的潜力
以往研究表明,从推测的祖先中提取的突变可以在一定程度上提高蛋白的稳定性。然而,我们的研究进一步表明,即使是相对较新的祖先(例如来自早期脊椎动物或四足动物)也可通过重建实现约20–30°C的热稳定性提升,同时保留功能多样性。
我们推测,祖先蛋白的耐热性可能不是直接受到自然选择压力的结果,而是从远古时代高温环境中逐渐遗留的特性。在海洋温度可能达到55–60°C的古代环境下,祖先酶的耐热性被保留下来。随着时间推移和中性漂变,这些特性逐渐被弱化。
溶剂耐受性与热稳定性
有机溶剂可能通过剥离关键水分子或改变蛋白质表面、活性位点或核心的分子间相互作用强度而影响其结构和活性。尽管溶剂耐受性与热稳定性不一定相关,已有研究表明两者可能共存,包括在极端酶中。耐热酶中增强的疏水核包装可能有助于减少暴露于有机溶剂的风险,这一观察与本研究祖先CYP3酶的表现一致。
工业和生物学应用
通过本文提出的ASR方法生成的耐热酶为生物物理表征和进一步的蛋白工程提供了稳健的模板。使用祖先作为模板进行重新进化,可以在后续的工程中更有效地缓冲有害突变。此外,祖先酶可以避免现存形式重组时常见的上位效应问题。我们将此方法称为**“用于定向进化的祖先组合文库”(CLADE)**,可用于构建寿命足够长以满足生物催化、合成生物学等领域需求的酶。
对于细胞色素P450酶,本文开发的耐热祖先形式为工程专门化单加氧酶提供了急需的起点。这些酶在制药工业中有广泛应用,包括用于药物代谢物的生产以辅助结构鉴定。目前,重组人CYP3A4和工程化的CYP102突变体被广泛使用。本文提出的方法将显著扩展可用于制药开发的候选酶形式,并推动P450在精细化学品合成、生物传感器及污染环境修复等领域的应用。
对于KARI酶,通过ASR提高其热稳定性和活性,可为进一步工程提供优化的起点。本文所示的祖先KARI在50°C时的比活性(17 U mg⁻¹)明显高于现存极端生物来源的KARI变体。这为其应用于生物燃料生产等工业过程提供了更强的竞争力。
(李俊锋摘译)
原文链接:https://doi.org/10.1038/s41929-018-0159-5
高颜值免费 SCI 在线绘图(点击图片直达)
最全植物基因组数据库IMP (点击图片直达)

往期精品(点击图片直达文字对应教程)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|















































 1924
1924

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









