







1 进化算法
在学习最优模型参数的时候,梯度下降并不是唯一的选择。在我们不知道目标函数的精确解析或者不能直接计算梯度的情况下,进化算法是有效的。
进化算法的灵感来源于自然选择,具有有利于生存的特征的个体可以世代生存,并将好的特性传给下一代;具有不利于生存的特正的个体则会被不断淘汰,最后减少甚至消失。进化是在选择过程中逐渐发生的,进化使得种群可以更好地适应环境。
下面这张图可以很好地解释进化算法的想法,一开始岛上有数量相等的白老鼠和黑老鼠,但是因为黑老鼠和土地的颜色相近,所以老鼠的天敌不易于发现这一种类的老鼠。而白老鼠由于过于显眼,在不断地被吃掉的过程中,慢慢地被淘汰了。久而久之,最后剩的多的就是黑老鼠。
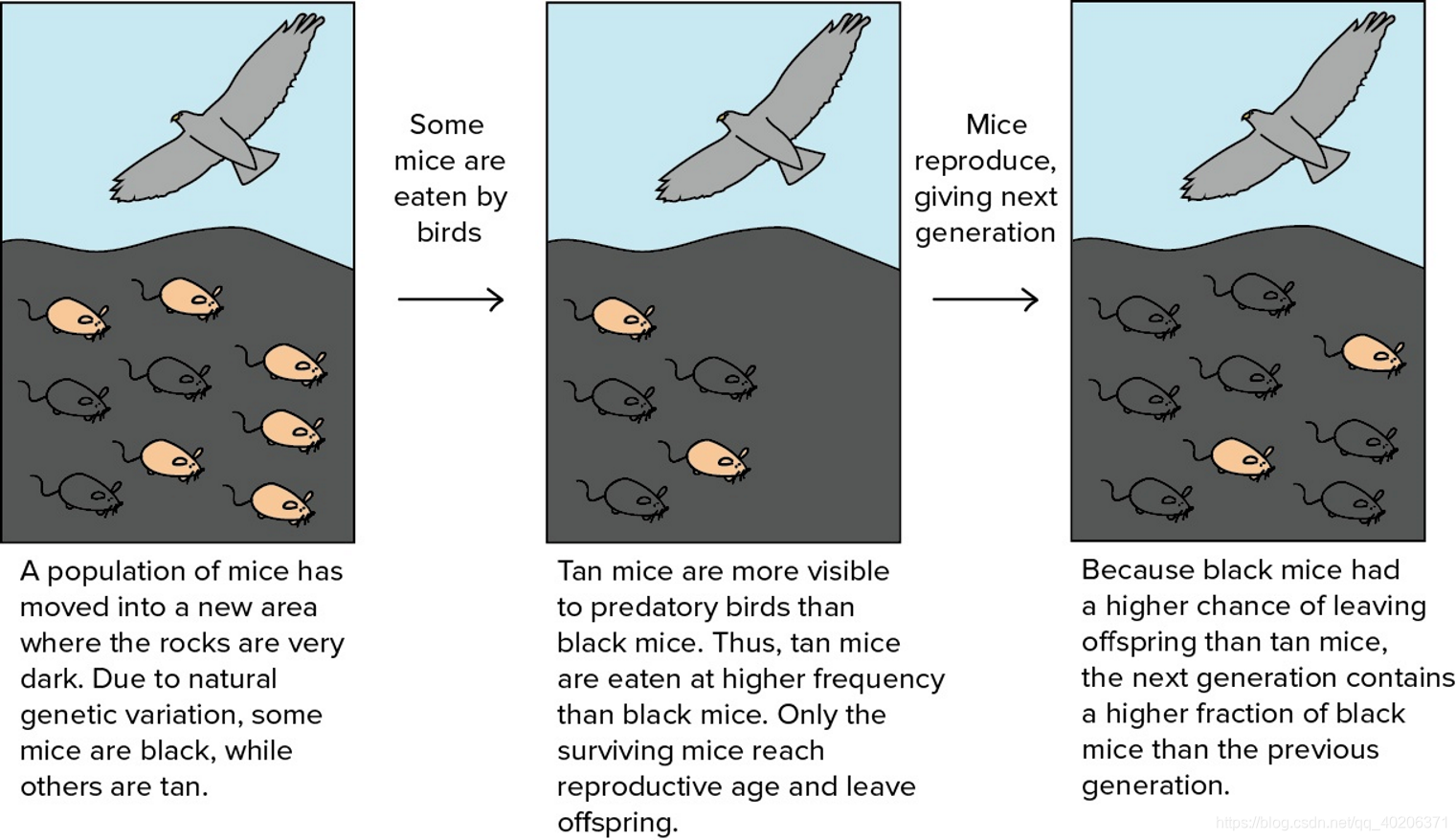
进化算法旨在优化无法直接建模的函数,核心是采样和更新。它在在每一代估计基因型的实值,然后保留最佳的基因型,其他的基因型被丢弃,精英基因被保留并产生。
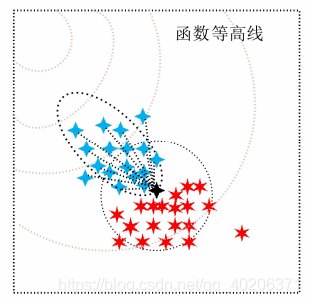
如下图所示,虚线圆是初始的正态分布,星星表示采样生成的后代,蓝色的星星表示排序后被选中的后代,红色表示没有选中的。那么虚线椭圆就是新一代的概率采样分布。
进化算法作为一种通用的优化方式,可以归纳为:
1,假设我们想优化一个函数f(x),但f(x)的梯度难以求解。只能得到在特定x下f(x)的确定值。
2,我们认为随机变量x的概率分布p(θ,x)是函数f(x)优化问题的一个较优的解,θ是分布p(θ,x)的参数。我们最终的目标是找到θ的最优配置。
3,在给定分布形式(例如,高斯分布)的情况下,参数 θ 包含了最优解的知识,在一代与一代间进行迭代更新。
4,
2 简单高斯进化策略

![]()
![]()

即 在分布中随机采样

4,
5,重复(2)-(4)步直到结果足够好
3 协方差自适应进化策略CMA-ES
3.1 基本原理

协方差矩阵自适应演化策略(CMA-ES)通过使用协方差矩阵 C 跟踪分布上得到的样本两两之间的依赖关系,解决了上述这一问题。新的分布参数变为了:
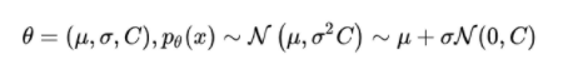
3.1.1 协方差矩阵性质
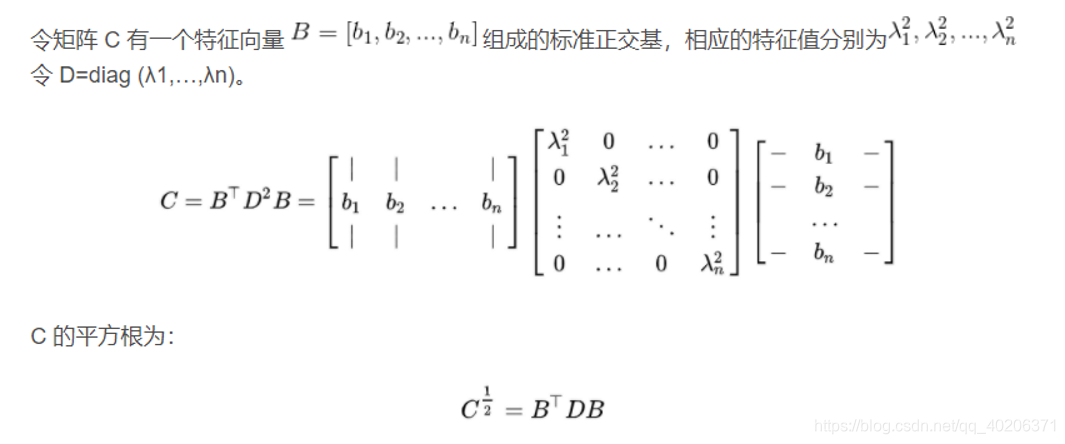
1,始终都是对角阵
2,始终都是半正定矩阵
3,所有的特征值为非负实数
4,所有的特征向量都是正交的
5,特征向量可以组成一个Rn的标准正交基
3.1.2 本节涉及符号及其意义

是在分布
上随机采样;
是在分布
上随机采样。
是第t代的平均值;
是第t代采样时候的步长;
是第t代的协方差矩阵。
和
是
进行特征值分解的结果(见3.1.1)。
是第t代步长σ和协方差矩阵C的进化路径
分别是μ,σ,pc,C的两种路径(后文会涉及)的学习率
是步长σ更新时候的阻尼系数
3.1.3 更新均值
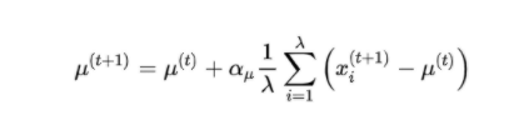

μ的学习率为1的时候,就是
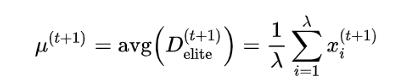
3.1.4 控制步长



即 ~N(0,I) (第二行式子是因为每一个yi都是N(0,C)中采样的)
![]()

有了演化路径之后,我们就可以根据演化路径来更新我们的σ进化路径了

注:这里公式有一处笔误,第二行根号里面第一项为

注:这里调整步长的思想来源于CSA-ES(累积步长调整策略),这种思路使用多代来记录同一条路径,即进化路径。演化路径是连续几代路径的总和,在每次迭代中对多个精英子代的搜索方向做加权平均,相反分量相互抵消,相同分量进行叠加。如果成功的步骤都朝着相同的方向发展累加起来,那么该方向的发展路径就会相对较长,我们可以用一个较大的步长来代替该方向的总进化距离;反之,则需要简短步长,即:

3.1.4.1 Polyak算法回忆

3.1.5 调整协方差矩阵


看起来是可以的,但是只有当我们选择出的种群足够大,上述估计才可靠。然而,在每一代中,我们希望使用较小的样本种群进行快速的迭代。这就是 CMA-ES 发明了一种更加可靠,但同时也更加复杂的方式去更新 C 的原因。它包含两种独立的演化路径:


注:n是每个样本的维度,λ是每一代取的精英样本数



3.2 总体CMA-ES
这是一篇论文里面的CMA-ES的总体方案;第一步取值;第二步挑精英值;第三步,更新均值μ;第四步更新 步长σ需要的演化路径;第五步更新步长;第六步更新path2 需要的C的演化路径;第七步我个人觉得不需要?(同时在另一篇论文“基于改进的CMA_ES的仿真足球机器人的行走优化”里面,也是直接第八步的,这个欢迎讨论哈!);第八步更新协方差矩阵



















 716
716

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言











