






农科院张西美等-独特的土壤微生物群落构建模式和机制(附招聘启事)
https://doi.org/10.1016/j.isci.2022.103893
2022年3月18日,细胞出版社(Cell Press)旗下期刊iScience(《交叉科学》)在线发表了中国农业科学院农业环境与可持续发展研究所张西美团队的研究成果《Distinctive pattern and mechanism of precipitation changes affecting soil microbial assemblages in the Eurasian steppe》,该研究依托内蒙古草原5年的降水实验平台,利用扩增子深度测序、宏基因组测序和宏蛋白组技术,揭示了不同降水强度驱动土壤微生物群落构建的独特模式和机制,挑战了“环境变化幅度增加提高微生物群落构建过程中确定性过程相对于随机过程的重要性”的传统观点。
摘要
在全球变化背景下,降水的强度可能会增加或减少,但不同降水变化下土壤微生物群落组装的模式和机制的研究仍相对欠缺。传统观点认为,环境变化幅度的增加会提高微生物群落构建过程中确定性过程相对于随机过程的重要性,我们发现,降水强度改变30%时,确定性的竞争排斥过程确实降低了处理内分类组成的差异,但是当降水强度改变60%时,降水通过促进随机生态漂移过程(随机出生/死亡)增加了处理内分类组成的差异,这一独特的群落构建变化模式与传统观点形成了鲜明对比。不仅如此,60%降水下增加的分类学差异/随机性还转化为了基因、蛋白质和酶水平的功能差异/随机性。总体而言,我们的研究结果揭示了降水变化驱动土壤微生物群落组装的独特模式和机制,并证明了需要整合微生物分类学信息以更好地预测土壤微生物功能对降水变化的响应。
引言
土壤微生物群落可能拥有地球上最高的生物多样性,并执行着许多重要的生态系统功能。人类活动引起的气候变化可能会改变微生物群落的多样性、组成及生态系统功能,从而对气候变化产生正反馈或负反馈。温度和降水的变化属于气候变化,但与大量温度升高的相关研究相比,降水变化对土壤微生物群落的影响仍未得到充分探索。在气候变化的背景下,降水可能会以不同的强度增加或减少,但很少有研究调查土壤微生物群落对这些降水变化的响应模式和机制。由于水占细胞生物重量的60%以上,因此降水应该是构建陆地生物群落的最重要的生态因素。在大的时空尺度上,降水是比其他生态因子更重要的地上植物群落构建的驱动因子。在自然降水梯度和模拟降水实验中,降水在构建地下微生物群落方面也发挥着一定的作用。然而,对于土壤细菌群落,土壤pH值通常被认为是在不同时空尺度上群落构建的最重要因素,并且很多研究发现,不同类型的人为环境变化主要通过调节土壤 pH 而不是土壤含水量来影响土壤微生物群落。因此,以往的研究可能低估了降水对土壤微生物多样性的直接影响和关键作用。降水在构建土壤微生物群落中的重要性被低估,这表明降水可能通过尚未发现的独特途径和机制影响土壤微生物组成。
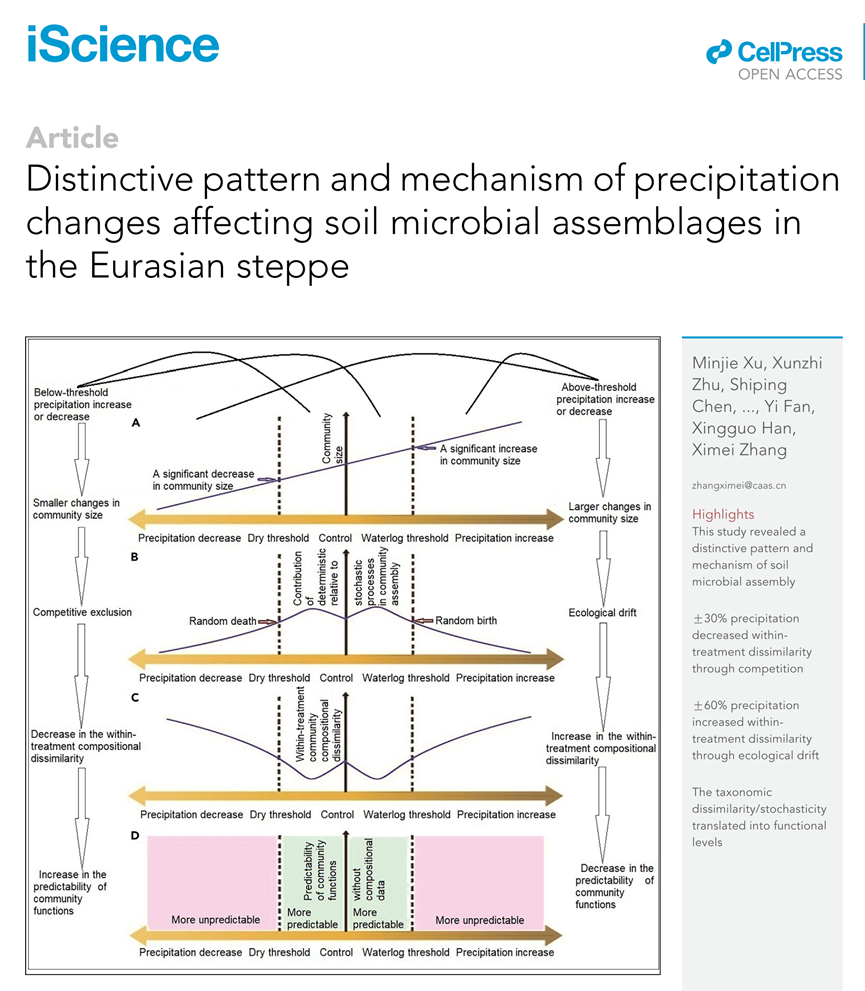
生态学家长期以来一直对群落内物种共存的机制感兴趣。传统的生态位理论认为,不同物种在一个群落中共存是因为它们具有不同的生态位,例如它们受到不同资源的限制。在一个群落内,不同的物种具有不同的适应度,它们的丰度与其适应度是一致的。随着环境条件的变化,它们的适应度会发生不一致的变化,导致它们的丰度和群落结构发生变化。一般来说,生态位理论侧重于选择的确定性影响。近年来,生态学家提出了生态中性理论,认为群落结构的变化更多地受到与生态漂变、迁移和物种形成相关的随机过程的影响,而与物种间的适应度差异无关。与物种间适应度差异相关的确定性过程,如竞争排斥和环境过滤,以及与物种间适应度差异无关的随机过程,如生态漂移和随机扩散/定殖,都被发现是潜在的驱动微生物群落组装的因素。人为环境变化可以通过调节确定性和随机过程来影响群落组装。例如,土壤酸化已被证明主要通过确定性过程驱动土壤微生物群落组装,而养分富集主要通过随机过程驱动淡水池塘中生产者和动物群落的组装。环境变化强度的增加被认为会对群落施加强大的选择压力,并有利于适应度更高的分类群,因此与随机过程相比,确定性过程的相对贡献会增加。鉴于降水可能是影响土壤微生物构建的最重要因素,我们提出了一个关于降水变化驱动微生物群落构建的独特模式和机制的假设(图 1)。当降水量增加(或减少)低于某个阈值并且土壤栖息地的承载能力(潜在的最大群落大小)仅发生很小的变化时(图 1A),确定性过程(即竞争排斥)的重要性在土壤微生物群落组装中会增加(图 1B),这将导致诸如嗜湿(或耐旱)微生物物种的丰度增加。反过来会导致处理组内组成差异的减少(图 1C)和群落功能的可预测性增加(图 1D)。当降水量增加(或减少)超过某个阈值时,土壤栖息地可以支持更多(或更少)的微生物个体(图 1A),这将刺激随机出生(或死亡)过程(图 1B)。因此,处理内组成差异和群落功能的不可预测性会增加(图 1C 和 1D)。换句话说,高于阈值的降水变化通过刺激生态漂变(随机出生/死亡)的随机过程来影响微生物群落的组装,而群落组成的这些随机变化可能有助于解释为什么在以往的研究中降水的重要性被低估了。
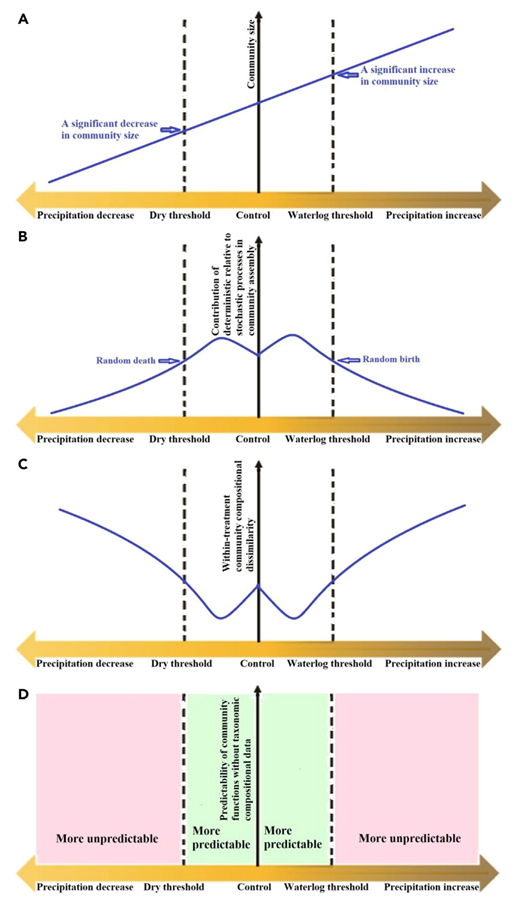
图1. 降水变化驱动土壤微生物组成改变的概念框架(A-D)
(A)降水变化如何影响群落大小的概念框架,(B)群落组装中确定性过程相对于随机过程的相对贡献,(C)处理内群落组成差异,(D)导致变化群落功能的可预测性。
为了验证提出的假说,我们在中国内蒙古的草原生态系统中进行了一项为期5年的降水实验,其中包含五种处理(减雨60%、减雨30%、对照、增雨30%、增雨60%)。根据该地区历史生长季降水量,最干燥(或最潮湿)年份的降水量低于(或高于)长期平均降水量约60%。因此,增加或降低60%降水用于高于阈值的降水变化,而增加或降低30%降水用于低于阈值的变化。我们利用16S rDNA扩增子深度测序量化了所有土壤原核生物群落(细菌和古细菌)的分类多样性,并使用宏基因组和宏蛋白质组测序测定了土壤微生物功能多样性。我们使用零模型方法将观察到的群落与随机组装的群落进行比较,并计算确定性和随机过程在驱动群落构建中的相对贡献。越来越多的研究表明,随机过程是微生物群落组装的关键驱动因素,许多人将这种模式归因于微生物群落中大量稀有物种。为了评估稀有物种对确定确定性与随机群落组装的重要性的贡献,我们改变了深度测序扩增子数据分析中包含的稀有操作分类单位 (OTU)的比例。为了确定高于阈值的降水变化影响微生物群落组装的随机过程是否是一种独特的机制,我们重新分析了先前在中国北方草原生态系统中进行的多因素实验的土壤微生物群落组成数据。以检验在浇水(高于阈值降水增加)、变暖、 N添加、磷添加、割草等13种环境变化中,浇水处理下微生物群落组装中随机过程相对于确定性过程的相对贡献是否大于其他类型的环境变化下的相对贡献。
实验设计
本研究以位于内蒙古西乌旗的干旱半干旱草地生态系统为研究对象,采集降水处理5年后的土壤样品,通过深度扩增子测序、宏基因组和宏蛋白组测序技术,分析5种降水强度(-60%、-30%、 0 (对照、自然降水)、+30% 和+60%)对土壤微生物群落构建的影响。
结果和讨论
1.±60%降水处理分别用作低于和高于阈值的变化
本研究的降水处理包括5个水平(-60%、-30%、0(对照、自然降水)、+30%和+60%),每个水平4个重复。在处理5年后收集土壤样品。降水量的增加显著增加了土壤含水量、土壤速效N含量、植物生物量和微生物生物量C和N含量,并降低了土壤温度(p<0.05;图2);然而,降水的增加对土壤pH值、土壤有机碳和总氮含量、溶解有机碳含量和植物丰富度没有显著影响。虽然-30%的降水使土壤含水量仅减少了10.92%,-60%的降水使土壤含水量减少了26.57%(>2*10.92%;图2A);+30%的降水仅增加了4.44%的土壤含水量,但+60%的降水增加了15.46%(>2*4.44%;图2A)。同样,尽管-30%的降水仅将微生物量N含量降低了8.82%,-60%的降水将其降低了55.45%(>2*8.82%;图2K)。这些结果表明,土壤含水量和微生物量似乎对30%的降水处理具有一定的缓冲能力。因此,30%的降水改变被用作阈值以下降水变化,它仅在很小程度上改变了土壤生境的承载能力(潜在的最大群落大小),而60%的降水改变被用作高于阈值的降水变化,这极大地改变了承载能力,支持我们在图1A中的假设。
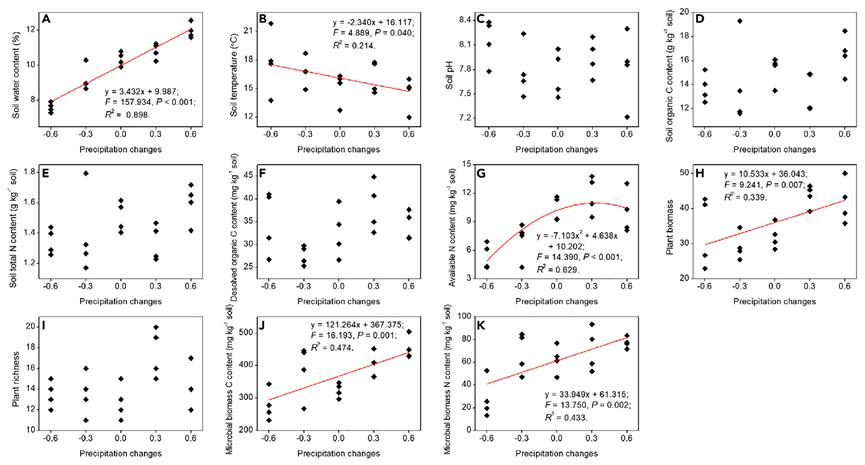
图 2 降水变化对生物和非生物指标的影响
2.低于和高于阈值的降水变化分别减少和增加了处理内的组成差异
为了检测相对稀有的微生物物种和精确测定微生物群落组成,我们对五个降水强度水平的四个重复群落进行了针对16S rDNA序列的深度扩增子测序。从20个细菌群落中获得了716,472–851,990条序列,从20个古菌群落中获得了148,106–244,669条序列。这些序列用97%的相似性阈值划分为OTU。在样品中共观察到13205个细菌OTU和214个古细菌OTU,平均每个群落有9420个细菌OTU和96个古细菌OTU。单因素方差分析表明,实验降水梯度对细菌和古细菌OTU丰富度的影响不显著(p>0.05),并且在稀有OTU数量增加的情况下,该结果仍是稳健的。这些结果表明,实验降水不影响土壤微生物OTU丰富度。
对土壤细菌和古菌群落分别计算了加权和未加权Unifrac距离,代表群落之间的系统发育距离。PERMANOVA(permutational multivariate analysis of variance)是一种置换方法,用于在任意距离度量的基础上测试ANOVA实验设计中变量对因素的响应,PERMDISP(permutational analysis of multivariatedisperss)是一种基于任意距离或差异比较组间离散程度的方法。PERMANOVA和PERMDISP分析结果均表明降水变化对土壤细菌和古菌群落的系统发育结构没有显著影响(p>0.05;表S2)。我们计算了基于OTU 相对丰度的Bray-Curtis差异矩阵来表征群落组成的变化。PERMANOVA分析结果显示降水处理不会导致细菌或古细菌群落组成发生任何方向性/系统性变化(p>0.05;图S1)。然而, PERMDISP显示降水梯度对处理内的组成变化有显著影响(p<0.01;图S1)。总体而言,这些结果表明降水梯度对微生物群落组成具有非定向/非系统影响。换句话说,虽然一个处理的所有重复群落的整体组成与对照没有差异,但一个处理的单个重复群落的组成可能与对照的整体组成不同,并且该处理的四个重复群落的组成可能不同。某种处理以不一致和不可预测的方式偏离了对照的整体构成。这是一个有趣的观察结果,因为经常观察到实验处理会定向/系统地改变群落组成,而不是非定向/非系统地改变群落组成。例如,加氮处理后的土壤微生物群落组成相对于对照有方向性变化。
为了检验群落组成对降水梯度的非系统响应是否源于大量稀有OTU,我们通过改变分析中包含的稀有OTU的数量来分析群落数据(例如,包括相对丰度>1%、>0.001甚至更低相对丰度的OTU)。结果表明稀有OUT的数量变化没有改变PERMANOVA不显著和PERMDISP显著的结果(表S2),这表明常见和稀有的OTU在对降水变化的响应方面没有差异,这一发现也证实了群落组成的反应不是系统性的。
在所有13,205个细菌OTU中,47.38%的OTU的相对丰度在五个降水强度水平中有显著差异(Levene检验:p<0.05;表S3),与PERMDISP结果一致;只有6.11%的OTU的相对丰度与土壤含水量显著相关(Pearson相关性:p<0.05;表S3),与PERMANOVA结果一致。同样,在214个古细OTU 中,75.70%的OTU相对丰度在五个降水强度间有显著差异,只有3.74%的OTU相对丰度与土壤含水量显著相关(表S4)。总体而言,这些结果证实了微生物群落组成对降水梯度的非系统响应。
PERMDISP 揭示了土壤细菌和古细菌群落的一致模式,即与对照相比,降水强度改变30%降低了处理内组成差异,而降水强度改变60%增加了处理内部的组成差异(图3A和3B)。这种模式在所分析的微生物群落包含稀有OTU数量增加时依然存在(图S2)。与±30%降水处理下处理内组成差异的降低一致,29.53%的细菌OTU和63.55%的古细菌OTU在两种处理下的相对丰度差异最小。与±60%降水处理下处理内组成差异的增加一致,28.50%的细菌OTU和41.59%的古细菌OTU在这两种处理下的相对丰度差异最大(表S3和S4)。这种模式支持我们在图1C中的假设,即低于阈值的降水变化(±30% 降水)降低了处理内的成分差异,而高于阈值的降水变化(±60% 降水)增加了处理内的差异。
3.低于和高于阈值的降水变化分别增加了确定性和随机过程的相对重要性
Mantel检验用于识别解释微生物群落组成变化的因素(植物群落指数和土壤理化指数)。细菌和古菌群落组成的变化与土壤含水量、温度、pH、速效氮含量和植物群落组成显著相关(p<0.05;表S5)。然而,Mantel检验的r值通常小于0.5,这意味着这些土壤和植物指标无法解释微生物群落组成的大部分变化。VPA结果也表明,细菌和古细菌群落组成的大部分变异(75.7-85.7%)不能用测量的环境变量来解释(图S3),这表明随机过程和/或未测量的环境变量在土壤微生物群落组装中发挥了重要作用。
为了分析确定性与随机过程在每种处理下对土壤微生物群落组装过程的相对贡献,我们使用零模型方法检验了细菌和古细菌群落的处理内差异(β多样性)与随机组装群落的预期差异之间的一致性。该方法控制了α和γ多样性的影响,因此可以确定生态过程对β多样性的影响。对于对照的群落,观察到的差异矩阵和预期的差异矩阵之间没有显著差异(p>0.05;表 S6),这表明随机过程是对照的微生物群落组装的主要驱动力。在对其他生态系统的研究中发现的微生物群落组成受到随机过程控制的普遍现象通常被认为是由于存在大量稀有物种。然而,我们的研究在稀有OTU数量增加的情况下,观察到的差异和预期的差异之间一致的结果依然成立(表S6),这表明微生物群落组装的随机性不是主要由大量稀有物种引起的。因此,微生物群落组装的随机性可能是由微生物的其他特征决定的,例如它们的小体型和短的世代时间;然而,这个假设需要证实。
对于其他四种降水处理,观察到的处理内差异与随机组装群落的预期差异之间的差异在40例(2个微生物组*5个OUT稀有种水平*4个降水梯度)中有35例不显著(表S6),这表明随机过程是不同降水强度下土壤微生物群落组装的主要驱动力。因此,40个案例中只有5个差异显著(p<0.05;表S6)的结果表明,降水变化仅对驱动土壤微生物群落组装的随机与确定性过程的相对贡献产生了微弱的影响。
为了进一步研究降水变化的影响,使用零模型方法对所有降水强度下所有地块的群落数据进行分析,以计算SES(标准化效应大小)值。PERMDISP揭示了土壤细菌和古细菌群落的一般模式,其中±30%的降水增加了SES值,而±60%的降水降低了SES值(图3C和 3D),并且这种模式在增加分析中所包含的稀有OTU数量时是不变的(图S4)。这种模式表明,±30%降水增加了确定性过程相对于随机过程的贡献,而±60%降水增加了随机过程相对于确定性过程的贡献。我们使用直接计算方法进一步检查了结果,这些结果与 SES值的变化一致,从而证实了图3C和3D中的模式(详见数据S1、图S5 和表S7)。
图 3 降水变化影响群落组成和生态驱动力(A-D)
(A、B)降水变化对 PERMDISP 揭示的土壤细菌和古细菌群落的处理内成分差异和(C、D) SES标准效应大小值的影响。对于多重比较,显著 (p<0.05) 差异用大写字母区分。误差线代表标准误差。分析中包括相对丰度>0.01%的OTU。另请参见图 S1-S5 和 S8、数据 S1 以及表 S1、S2、S3、S4、S5、S6、S7 和 S8。
±30%降水下,确定性过程相对于随机过程的贡献增加与环境变化促进确定性群落组装的传统观点一致。环境过滤和竞争排斥都是确定性过程。虽然环境过滤会选择具有相似生态位的物种并减少处理内群落差异,但种间竞争会排除具有相似生态位的物种并选择具有不同生态位的物种,从而增加处理内差异。在所有观察到的处理内差异与随机组装群落的预期差异显著不同的五个案例中,有四例发生在±30%降水强度下,且观察到的处理内差异显著大于随机组装群落的预期差异(p<0.05;表S6),表明±30%降水增加了竞争排斥的确定性过程而非环境过滤过程的贡献。总体而言,这一结果支持了我们在图1B中的假设,即低于阈值的降水变化将导致土壤微生物群落组装中竞争排除的确定性过程的重要性略有增加。
±60%降水下,随机过程相对于确定性过程的贡献增加与增加环境变化的强度会增加确定性过程在群落组装中的重要性的传统观点不同。生态随机过程通常包括物种形成、物种扩散和定殖以及生态漂移(随机出生和死亡)。在这里,OTU被定义为具有>97%相似性的16S rDNA簇。尽管在5年的实验时间范围内,一些快速进化的基因可能会发生遗传变化,但16S rRNA基因非常保守,以至于进化对相似度>97%的OUT的物种形成没有明显的影响。换句话说,随机过程的贡献增加与物种形成过程无关。尽管实验处理不影响OTU的扩散,但它们影响了土壤理化条件(图2),从而影响了迁移OTU的定殖成功。我们使用Hubbell的中性模型估计了OTU扩散/定植对微生物群落组装的贡献。在这些处理中,土壤细菌和古细菌OTU迁移率为0.36-0.40(表S8),表明随机分散/定殖过程在土壤微生物群落组装中发挥了作用。然而,不同降水变化之间的OTU迁移率没有显著差异(表S8),表明随机扩散/定殖过程不是随机过程贡献增加的主要驱动力。相对于+30%(或-30%)降水(图2K),在+60%(或-60%)降水下微生物生物量的急剧增加(或减少),与+60%(或 -60%)降水下随机过程贡献增加的结果相吻合。换句话说,+60%(或-60%)的降水改变了土壤栖息地,从而支持更多(或更少)微生物个体的生长,并导致随机出生(或死亡),从而导致相对于确定性过程的随机过程的重要性增加。在未来的研究中,需要更先进的技术和/或改进的实验设计来直接确定原位微生物的出生/死亡率。总体而言,这一结果支持了我们在图 1B 中的假设,即高于阈值的降水变化通过刺激生态漂变的随机过程(随机出生/死亡)来影响土壤微生物群落的组装。
4.分类随机性转化为功能随机性并影响其可预测性
为了研究由降水变化引起的土壤微生物群落的处理内分类组成差异的变化是否转化为功能差异的变化,我们对土壤微生物宏基因组和宏蛋白质组进行了测定,并测量了酶活性。与分类组成的模式一致,没有观察到功能组成对降水变化的系统反应(PERMANOVA:宏基因组,F = 1.157,p = 0.315;元蛋白质组,F = 1.038,p = 0.390;酶,F = 0.751,p = 0.676);仅观察到非系统反应(PERMDISP:宏基因组,F = 5.389,p = 0.011;元蛋白质组,F = 5.988,p = 0.006;酶,F = 5.456,p = 0.007;图4A-4C)。特别是,与对照相比,±30%降水处理导致基因、蛋白质和酶的处理内差异小幅下降,而±60%降水导致这些处理内功能差异大幅增加(图4A–4C),这与处理内分类学差异的变化一致(图3A和3B)。此外,Mantel检验表明,基因、蛋白质和酶的差异与占土壤生态系统宏基因组97.64±0.06%的土壤细菌群落的分类差异显著相关(p<0.05;图4D-4F)。与显著的PERMDISP结果一致(图4A-4C),24.03%的基因、25.11%的蛋白质和33.33%的酶具有在五个降水强度下显著异质的相对丰度,并且相对丰度的最大差异经常发生在+60%或-60%降水处理中(表S10、S11和 S12)。此外,土壤微生物群落基于组装序列(the assembled sequences)(图S6A)处理内基因差异和基于所有注释和未注释短序列(reads)(图S6B)的处理内差异与基于注释序列的基因差异(图4A)的处理内基因差异和分类学差异(图3A和3B)具有相似的模式。总体而言,这些结果支持我们在图 1D 中的假设,即观察到的分类随机性变化转化为功能随机性,从而影响功能可预测性。
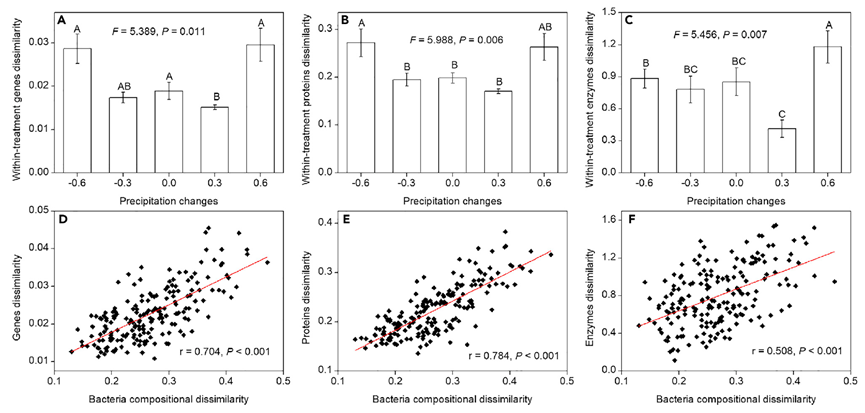
图 4 降水变化影响群落功能
(A-C)PERMDISP揭示的降水变化对土壤微生物群落基因、蛋白质和酶的处理内差异以及(D-F)Mantel检验揭示的基因、蛋白质、酶差异和细菌组成差异之间的关系的影响。对于多重比较(A–C),显著性(p<0.05)差异用不同的大写字母表示。误差线代表标准误差。另请参见图S6和表S9、S10、S11和S12。
结构方程模型(SEM)用于确定土壤理化指标、植物群落指标和微生物分类组成如何与微生物群落功能指标相关(图S7)。最终的SEM模型充分拟合了描述这些功能指标的解释变量数据(宏基因组,c2 = 3.24,p = 0.36;元蛋白质组,c2 = 3.24,p = 0.52;酶,c2 = 3.24,p = 0.52;标准化路径系数在图5中给出)。在这些确定的解释变量中,细菌分类组成是所有三个功能指标的标准化路径系数的绝对值最大的变量(0.64-0.94;图5)。这些结果表明,需要土壤微生物群落的分类信息来有效预测土壤微生物群落对降水变化的功能响应。
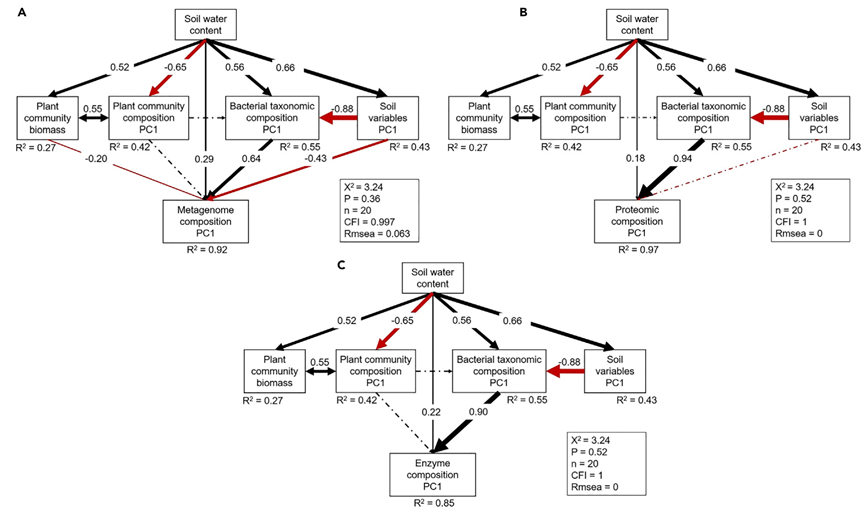
图 5. 群落功能的解释变量
(A-C)土壤理化指标、植物群落指标和细菌分类组成对微生物群落功能指标影响的结构方程模型分析(A,宏基因组;B,蛋白质组;C,酶)。实线箭头处的数字(p<0.05)是标准化路径系数(相当于相关系数),箭头宽度表示关系的强度。虚线箭头表示不显著的关系(p>0.05)。接近三个函数变量的数字(R2)表示模型解释的方差。另请参见图S6和S7以及表S10、S11和S12。
5.随机过程是阈值以上降水变化驱动微生物群落组装的独特机制
鉴于在±60%降水下,随机过程相对于确定性过程的贡献增加与传统的认识不同,我们假设这可能代表了一种新的机制,通过该机制,高于阈值的降水变化影响土壤微生物群落组装。为了验证这一假设,我们重新分析了在同一草原生态系统中进行的涉及13种环境变化实验的土壤微生物群落组成数据(参见数据 S2 中的实验设计细节),来检验与其他环境变化相比,阈值以上降水变化是否对细菌群落大小和随机生态过程的影响更大。多因素试验的增雨处理与植物生长季(5月至8月)的降水自然水平相比,5年处理的降水量大约高出+54.43%。每种环境因素都具有独特属性,因此每个环境因素的变化都可能对土壤微生物群落产生独特的影响。例如,气候变暖和养分富集分别通过增加土壤温度和养分含量来影响微生物群落。环境变化的强度也可能不同,环境变化强度的变化会影响群落组成确定性变化的幅度。为了控制13种环境变化强度的影响并描述其不同特性的效应,我们使用以下公式来标准化细菌群落组成的随机变化:[群落组成的随机变化的绝对值] /[群落组成的确定性变化的绝对值]。土壤细菌群落大小的变化使用以下公式进行标准化:[群落大小相对于对照的百分比变化的绝对值] /[群落组成的确定性变化的绝对值]。结果显示,细菌群落大小的标准化变化与细菌群落组成的标准随机变化呈正相关(斜率 = 0.056,F = 35.420,p < 0.001,R2 = 0.763;图6B),这种正线性关系即使在细菌群落大小标准化变化值最大的两个点(浇水、浇水+变暖)被排除在外的情况下依然成立(斜率 = 0.032,F = 4.658,p = 0.059,R2 = 0.341)。这些结果表明,高于阈值的降水增加导致土壤生境支持更多的微生物个体(更大的群落规模) ,从而刺激了随机出生过程,增加了随机过程对微生物群落组装的相对贡献。在所有五种与降水相关的环境变化中,有三种细菌群落组成的标准随机变化值相对较大(浇水、浇水+变暖和割草+浇水;图6A),表明高于阈值的降水增加对细菌群落大小和随机出生过程具有与其他因素相比更大的影响。另外两个细菌群落组成标准随机变化相对较小的降水相关环境变化均与N相关(N添加+浇水,割草+N 添加+浇水;图6A),表明降水升高的作用被N添加抵消。在所研究的草原生态系统中,添加氮通常会通过降低土壤pH 值,从而减小土壤细菌群落大小并改变细菌群落组成,因此,N添加和浇水对细菌群落大小有相反的影响,这表明当同时添加氮和浇水时,这些处理的效果会被抵消。细菌群落组成的标准化随机变化也因变暖而升高(图6A),其生态后果需要进一步研究。无论如何,这些结果表明,高于阈值的降水增加及其与其他因素的联合作用通过独特的随机出生过程影响了土壤微生物群落的组装。
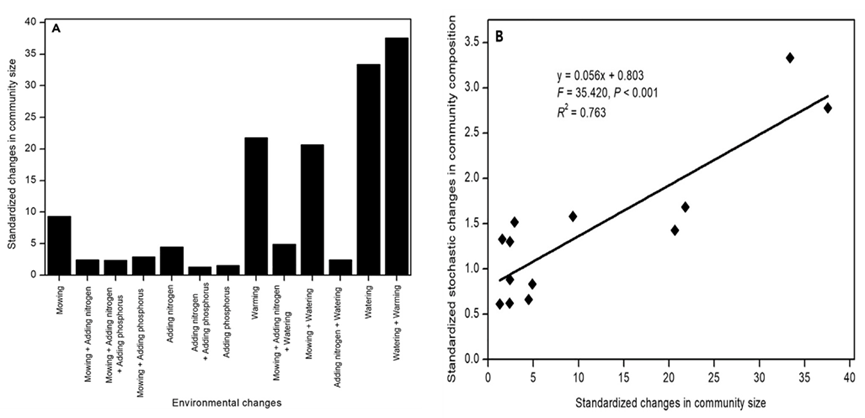
图 6. 降水变化影响土壤微生物组合的独特模式和机制的证据
(A)13种人为环境变化下土壤细菌群落大小的标准化变化,(B)及其与细菌群落组成标准化随机变化的关系。另见数据 S2。
结论
我们揭示了降水变化影响欧亚草原土壤微生物群落组装的独特模式和机制,并证实了我们所有的假设(图1)。简而言之,低于阈值的降水变化增加了竞争排斥的确定性过程在土壤微生物群落组装中的重要性,减少了处理内的组成差异,从而降低了处理内的功能差异。这导致了不需要微生物分类组成数据,仅使用土壤理化指标就能很好的预测群落功能。相反,高于阈值的降水变化促进随机出生(或死亡)过程,增加了处理内的组成差异,并使群落功能变得更加不可预测,从而需要使用分类组成数据预测群落功能。
我们这项研究中的重点是诸如不同土壤有机质分解代谢途径相对丰度(图S8)的群落功能组成,因此,分类组成的随机性会导致功能的随机性和不可预测性。如果我们专注于整个群落的功能(例如,群落呼吸)或单个功能(例如,N固定),则功能的大小将更可预测。例如,群落呼吸量可能与降水梯度呈线性趋势,这与微生物群落大小的模式相似(图2K)。降水变化对各种整体功能和特殊功能的影响和机制有待进一步研究。
不同生态系统(例如,从赤道到北极)的自然降水梯度如何影响随机过程和确定性过程对土壤微生物群落组装的相对贡献也需要进一步研究。在这里,我们假设随机出生过程会导致更多的突变甚至物种形成,随机死亡过程会导致大时空尺度上的物种灭绝。由于随机效应和现象更容易被忽视;因此,高强度降水变化的独特随机机制表明,在之前的研究中,降水在驱动土壤微生物组合中的重要性可能被低估了。特别是,降水或土壤含水量对微生物群落组装的影响可能部分被土壤pH值掩盖。例如,在本研究的对照群落中,古菌群落组成PCoA(主坐标分析)轴1分别与土壤含水量和pH值存在显著的正负关系(表S7),这表明土壤环境小空间尺度的异质性导致微生物群落组成的变化。因此,梳理土壤pH值的影响对于表征降水变化影响土壤微生物组装的随机机制非常重要。
研究的局限性
在本研究中,30%和60%分别被用作低于和高于阈值的降水变化。然而,确定确定性过程让位于随机过程的确切阈值需要进行更多水平的降水实验,而且降水增加和减少的阈值很可能是不同的。阈值可能具有特殊的生理意义,在这里,我们假设微生物可以适应消耗增加的资源(或容忍资源短缺)以响应低于阈值的降水增加(或减少),并且它们无法在生理上适应高于阈值的降水增加(或减少),但他们可以通过出生过程增加其丰度来消耗额外的资源(或通过减少其丰度来处理资源短缺;即死亡过程)。此外,需要更多的时间序列样本来监测未来研究中的随机出生/死亡过程。
通讯作者张西美简介

l中国农业科学院农业环境与可持续发展研究所研究员,博导。
l国家万人计划“青年拔尖人才”,农业农村部“杰出青年农业科学家”,中国农业科学院“农科英才”。
l长期从事土壤微生物多样性的维持和调控机制、土壤微生物地理学、土壤微生物的生态系统功能、植物-微生物互作研究。
l研究成果以第一作者或通讯作者发表在The ISME Journal、Global Change Biology、iScience、Soil Biology and Biochemistry、Science China - Life sciences、mSystems、Environmental Microbiology、Molecular Ecology等专业期刊上。
l正在主持国家自然科学基金区域创新发展联合基金重点支持项目、国家自然科学基金面上项目等项目。
附:中国农科院高薪诚聘生物信息学研究助理(来年优先读博)
一、团队介绍
张西美老师团队隶属于中国农业科学院农业环境与可持续发展研究所,研究组组长张西美研究员长期从事土壤微生物多样性的维持和调控机制、土壤微生物地理学、土壤微生物的生态系统功能研究。研究成果以第一作者或通讯作者发表在The ISME Journal、Global Change Biology、Soil Biology and Biochemistry、iScience等专业期刊上。个人主页:http://ieda.caas.cn/rck/276832.htm。
团队的研究方向为利用扩增子和鸟枪法宏基因组的高通量测序技术,研究土壤微生物多样性、生物地理学、微生物进化等。现因课题需要,招聘生物信息学专业或具有扩增子、宏基因组分析经验的研究助理1名,办公地点为北京市海淀区中关村南大街12号。
二.申请条件
(1)生物信息学专业或具有扩增子、宏基因组分析经验,具有相关专业的硕士学位,具有团队协作精神;
(2)熟悉高通量测序数据分析和整合,具有微生物高通量组学大数据分析经验者优先考虑;
(3)熟练运用R/Python/Perl/C/Matlab等编程语言,熟悉Linux/Unix系统环境;
(4)品学兼优,身体健康,年龄在35周岁以下。
三、薪酬待遇
(1)执行国家、中国农业科学院及农业环境与可持续发展研究所有关科研助手的相关规定;
(2)工资每月8k-1.2k,具体面议。
(3)优先支持攻读本研究组2023年度博士。
四、应聘方式
请申请者提供以下材料:
(1)本人简历(包括本科、硕士的就读时间、学校、院系和专业和导师);
(2)教育经历:本科、硕士毕业论文pdf版本;
(3)研究成果(发表论文pdf等);
(4)擅长的生物信息学分析相关技能和参与项目。
五、联系方式
有意向者请将报名材料发送至张老师邮箱: ximeizhang@sohu.com;也欢迎致信咨询项目详情。
收到您的简历后,课题组会尽快审核申请材料,对于符合要求并通过初审者,将会通知安排面试。招聘启事在2022年度长期有效。
猜你喜欢
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
文献阅读 热心肠 SemanticScholar Geenmedical
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份,另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。

学习16S扩增子、宏基因组科研思路和分析实战,关注“宏基因组”
点击阅读原文,跳转最新文章目录阅读

















 274
274

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









