






奇病毒(Mirusviruses)将疱疹病毒与巨型病毒联系起来
Mirusviruses link herpesviruses to giant viruses
翻译:周之超@UW-Madison

Article,2023-4-19,Nature,[IF 69.504]
DOI:10.1038/s41586-023-05962-4
原文链接:https://www.nature.com/articles/s41586-023-05962-4
第一作者:Morgan Gaïa,Lingjie Meng(孟令杰)
通讯作者:Tom O. Delmont
主要单位:
Génomique Métabolique,Genoscope,Institut François Jacob, CEA, CNRS, Univ Evry, UniversitéParis-Saclay
巴黎大学-萨克雷分校François Jacob研究所
Bioinformatics Center, Institute for Chemical Research, Kyoto University
京都大学化学研究所生物信息学中心
- 摘要 -
DNA病毒对细胞生物的生态学和进化有重大影响,但它们的整体多样性和进化轨迹仍然难以捉摸。在这里,我们对海洋真光层进行了系统发育指导的基因组解析宏基因组调查,发现了一个能够感染浮游生物的新门Mirusviricota (拉丁文:惊奇、奇怪),而它们是疱疹病毒(Herpesviruses)的亲属。这个大的单支系的病毒形态发生模块证明它们是典型的来自Duplodnaviria域的病毒,其多个组成部分强烈表明它们与动物感染性疱疹病毒拥有共同祖先。然而,相当一部分Mirusvirus的基因,包括疱疹病毒中缺少的标志性转录机器基因,是来自另一个病毒域Varidnaviria的巨型真核DNA病毒的密切相关的同源物。这些连接Mirusviricota与疱疹病毒和巨型真核病毒的显著的奇特属性得到了100多个环境Mirusvirus基因组的支持,包括一个接近完整的432 kbp的连续基因组。此外,Mirusvirus是在真光层中被描述为最丰富和最活跃的真核病毒之一,它编码了从一极到另一极的真核微生物感染过程中使用的各种功能。Mirusviruses的普遍性、功能活性、多样性和非典型的奇特属性表明,Mirusviricota在海洋生态系统的生态学和真核生物DNA病毒的进化中具有长期的作用。
- 引言 -
大多数双链DNA病毒被划分为两个主要领域:Duplodnaviria和Varidnaviria。Duplodnaviria包括有尾的噬菌体和相关的Caudoviricetes类的古菌病毒,以及疱疹病毒目真核生物病毒。Varidnaviria包括核细胞病毒门(Nucleocytoviricota)的大型和巨型真核DNA病毒,以及具有无尾二十面体衣壳的小型病毒。这两个域是根据非同源的病毒形态发生基因集合(病毒模块)建立的,包括那些编码结构上不相关的主要衣壳蛋白(MCPs)的基因,在Varidnaviria和Duplodnaviria中分别具有 "双果冻卷 "和HK97折叠。这两个领域在所有的生命领域都有代表的宿主,其各自的祖先的宿主被认为可以追溯到最后一个普遍的细胞祖先。
在Duplodnaviria中,Caudoviricetes的细菌和古菌成员表现出连续的基因组大小,从大约10kb到>700kb,而仅限于动物宿主的疱疹病毒则更均匀,基因组大小在100-300kb之间。疱疹病毒可能是从噬菌体进化而来的,但由于缺乏动物界以外的相关病毒,人们对其确切的进化轨迹产生了疑问。Varidnaviria的成员也表现出广泛的基因组大小,从大约10 kb到>2 Mb,但在核细胞病毒门的大型和巨型病毒与其余基因组小于50 kb的Varidnaviruses之间存在复杂性的不连续性。有人认为,核细胞病毒门是从一个较小的Varidnavirus祖先进化而来的,但其中需要获得多个信息基因(信息模块)的复杂化过程仍有待充分理解。
Caudoviricetes和Nucleocytoviricota(核细胞病毒门)中的病毒在阳光下的海洋(真光层)普遍存在,它们在调节浮游生物的群落组成和繁殖活动中发挥着关键作用。在这里,我们进行了一个以单一标志基因的系统发育为指导的浮游生物DNA病毒的基因组解析宏基因组调查。该调查涵盖了来自Tara Ocean考察队表层海洋样本的近3000亿条宏基因组reads。我们对数百个种群基因组进行了表征和手动整理,扩大了核细胞病毒门的已知多样性。然而,最值得注意的是,我们的调查导致发现了一类感染浮游生物的疱疹病毒的亲属病毒,形成了一个被我们称为Mirusviricota的新门。Mirusviruses具有复杂的功能特征,在真光层海洋中广泛存在,它们积极地感染真核生物,填补了我们对浮游生物生态学认识的一个重要空白。尽管与疱疹病毒有明显的进化关系,但Mirusviruses编码的基因甚至更多地与核细胞病毒门有密切关系。Mirusviricota的这些显著的奇特属性将两个遥远相关的病毒域联系起来,为真核DNA病毒的进化提供了关键的见解。
- 结果 -
海洋真核生物病毒的基因组学
DNA依赖性RNA聚合酶亚单位A(RNApolA)和B(RNApolB)是进化上的信息基因标记,出现在大多数已知的感染海洋真核微生物的DNA病毒中,到目前为止只包括核细胞病毒门。在这里,我们利用来自Tara Ocean考察队的798个宏基因组(总共2800亿条reads,产生了大约1200万个长度超过2500 nt的contigs)的大型co-assemblies,对极地、温带和热带海洋的透光区的RNApolB基因进行了全面搜索。这些宏基因组包含了8个浮游生物大小的组分,从0.8微米到2000微米,都富含真核微生物。我们使用广谱的隐马尔可夫模型(HMM)特征识别了这些片段中的RNApolB基因,随后建立了一个包含2500多个非冗余环境RNApolB蛋白序列(相似度<90%)的数据库。这些序列的系统发育信号不仅重现了海洋核细胞病毒门的相当大的多样性,而且还揭示了以前未描述的深层分支支系,显然与生命的三个领域和其他已知病毒不相联系。我们推断,这些新的支系代表了以前未知的双链DNA病毒的谱系。
我们进行了系统发育指导下的基因组解析宏基因组调查,重点是核细胞病毒门的RNApolB和新的支系,以划定其基因组背景。我们对581个长度达1.45Mb(平均约270kb)的非冗余核细胞病毒门的MAG和117个长度达438kb(平均约200kb)的新支系的非冗余MAG进行了定性和手工整理。我们将以往宏基因组调查中的海洋核细胞病毒门MAG和来自培养和细胞分选的参考基因组纳入其中,构建了一个富含大型和巨型海洋真核生物双链DNA病毒的综合数据库(此后称为全球海洋真核生物病毒(GOEV)数据库)。GOEV数据库包含约60万个基因,并提供背景信息,以确定含有新的RNApolB支系的MAG的主要生态和进化特性。
发现第三个Duplodnaviria门
新组建的核细胞病毒门MAGs包含该病毒门的大部分标志基因,对应于病毒模块和信息模块。它们扩大了Megaviricetes类中Imitervirales、Pandoravirales、Pimascovirales和Algavirales目的已知多样性。此外,其中一个新的RNApolB支系显示出一个假定的新的核细胞病毒门类群,我们称之为Proculviricetes,它由六个MAGs代表,只在在北冰洋和南大洋中被检测到(图1)。其余新的RNApolB支系的111个MAG也含有与核细胞病毒门信息模块进化相关的关键基因,包括RNApolA和RNApolB、B族DNA聚合酶(DNApolB)和转录因子II-S(TFIIS)。单基因系统发育将这些MAG置于一个(DNApolB)或多个支系(RNApolA和RNApolB)中,总是在已知的核细胞病毒门的各目之间。由于其长度较短,TFIIS的信号较弱。对四种信息基因标记物进行的稳健的系统发育推断表明,它们代表了一个单系的病毒支系,其中的几个标志基因与已知的核细胞病毒门中的基因密切相关,但又有所不同(图1)。我们把这个支系中的病毒称为Mirusviruses(mirus是一个拉丁语单词,表示令人惊讶或奇怪)。
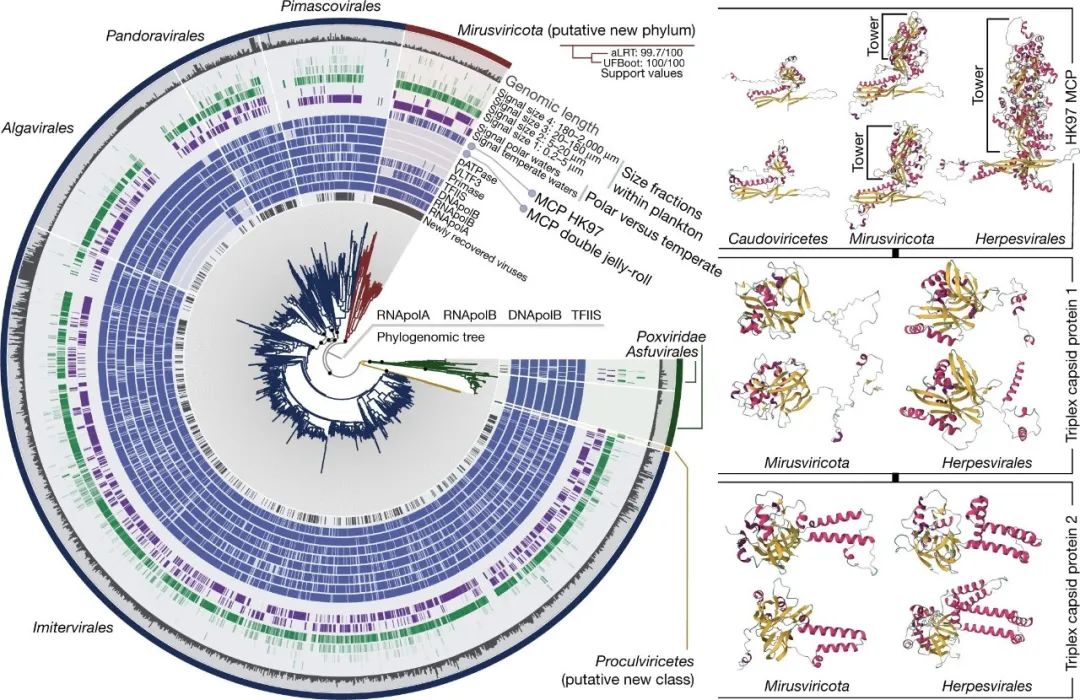
图1:核细胞病毒门、疱疹病毒目和Mirusvirus之间的进化关系
左图:使用后验平均位点频率混合模型(LG+C30+F+R10),在手工整理的RNApolA、RNApolB、DNApolB和TFIIS基因(3715个氨基酸位点)串联的基础上,从GOEV数据库(1,722个基因组)建立的最大似然系统树,并在Mirusvirus和其他病毒之间放置树根。突出显示系统发育树上面高支持度(近似似然比(aLRT)≥80和超快自举近似(UFBoot)≥95,黑色)或中支持度(aLRT≥80或UFBoot≥95,黄色)的分支点。树上装饰有补充信息的环,并用anvi'o进行可视化。右图:使用AlphaFold2获得的Caudoviricetes、Mirusvirus和herpesvirus代表的HK97 MCP的预测三维结构。蛋白质根据二级结构的特性进行了着色。该板块还显示了用同样的方法预测的Mirusvirus和疱疹病毒代表的三联体衣壳蛋白的三维结构。
Mirusvirus的MAGs被分成七个不同的亚族,M1到M7(从最多到最少),其中M1和M7分别由41个MAGs和一个MAG代表(图2a)。然而,值得注意的是,它们没有可识别的核细胞病毒病毒模块的同源物,包括双果冻卷MCP。相反,使用敏感的序列和结构相似性搜索对Mirusvirus基因簇进行注释,发现在大多数这些MAG中出现了HK97折叠MCP的远源同源物(图1)。这种MCP折叠的存在,仅与Caudoviricetes和Herpesvirales共享,表明Mirusviruses属于Duplodnaviria域。与这种MCP的鉴定相一致,进一步比较HMM profile和预测的三维(3D)结构,发现了Duplodnaviria病毒模块的其余关键成分,包括终止酶(ATP酶-核酸酶,DNA包装机器的关键成分)、门户蛋白、衣壳成熟蛋白酶和三联体衣壳蛋白1和2(图1)。编码这些蛋白的基因在Mirusviruses中的存在,确定了它们是真正的大型DNA病毒,能够形成与以前已知的Duplodnaviria域的病毒颗粒相似的病毒颗粒。值得注意的是,Mirusviruses HK97-fold MCP的系统发育推断再现了最初根据DNApolB、RNApolA、RNApolB和TFIIS确定的七个亚支系(图2b),表明病毒模块和信息模块的共同演化。
Mirusviruses、疱疹病毒和Caudoviricetes之间病毒模块蛋白质的广泛序列差异和长度差异阻碍了对新扩展的Duplodnaviria域进行有意义的系统发育推断。然而,这个模块的多个组成部分提供了关键的信息,澄清了Mirusviruses的进化轨迹。首先,两个三联衣壳蛋白,形成一个异源三聚体复合物,并通过与相邻的MCP亚单位的相互作用稳定帽壳,这在整个疱疹病毒中是保守的,但在Caudoviricetes中则是缺失的。第二,在疱疹病毒的MCPs中,HK97折叠结构域被称为底层结构域,负责帽壳的形成,它被点缀上一个“塔”结构域,从组装的帽壳表面伸出。塔状结构是HK97核心折叠的A子域中的一个插入部分。在Mirusviruses中,MCP蛋白也含有一个插入的A子域,尽管其大小要小得多(图1)。到目前为止,这个塔状结构域还没有在Caudoviricetes的任何成员中被发现过,包括所谓的巨型噬菌体(jumbo phage, 即具有非常大的基因组的噬菌体)。总的来说,三倍体衣壳蛋白和MCP塔代表了标志性的特征,表明与它们的细菌病毒和古菌病毒亲属相比,Mirusviruses和疱疹病毒之间的进化关系更为密切。
使用GOEV数据库和广泛的真核生物和其他病毒谱系对DNApolB基因进行系统发育推断,支持Mirusviruses相对于所有其他已知的双链DNA病毒支系的进化距离。单分支的Mirusviruses DNApolB被定位为疱疹病毒的姐妹支系,这两个真核Duplodnaviria的支系与真核Zeta型和Delta型DNApolB序列关系最密切,共同形成一个与其他病毒的DNApolB不同的具有高支持度的支系。综上所述,Mirusviruses、Caudoviricetes和Herpesvirales的病毒模块之间存在相当大的遗传距离,Mirusviruses MCP的不同三维结构和DNApolB的不同三维结构和DNApolB系统发育推论将Mirusviruses 牢牢地定位在Duplodnaviria域内,但不在先前描述的Uroviricota(Caudoviricetes)和Peploviricota(疱疹病毒)两个门类中,我们称之为Mirusviricota。
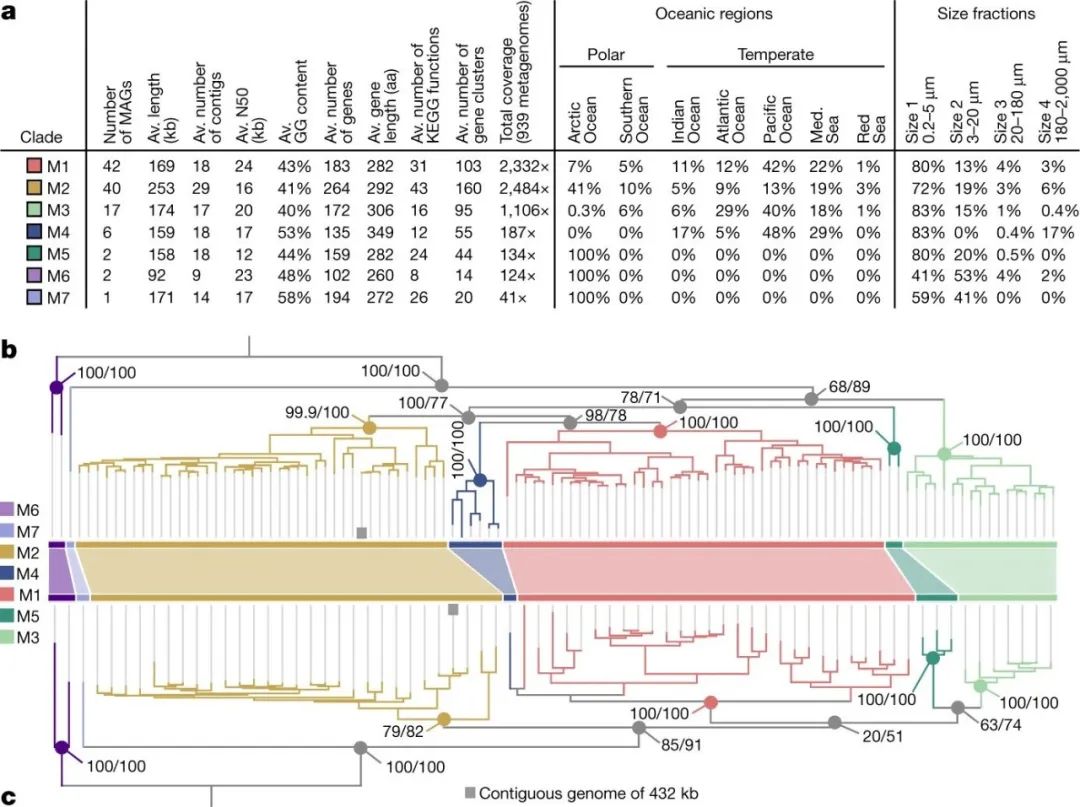
图2:Mirusvirus的基因组统计和进化
a, 七个Mirusviricota亚支系的基因组和环境统计。Av.,平均数;aa,氨基酸;KEGG,京都基因和基因组百科全书;N50,捕获总装配大小的50%所需的最短contig长度;Med,地中海。b,使用LG+F+R7模型,根据Mirusviricota MAGs的四个标志信息基因(编码RNApolA、RNApolB、DNApolB和TFIIS的基因;3715个氨基酸位置)建立的最大可能性系统发育树;c,使用LG+R6模型,根据MCP(701个氨基酸位置)的最大可能性系统发育树。两棵树都是在M6支系和其他支系之间生根。节点上的数值代表由Shimodaira-Hasegawa-like的aLRT(1,000次重复;左边的分数)和UFBoot(1,000次重复;右边的分数)计算的分支支持度(满分100分)。
Mirusvirus在功能上很复杂
111个Mirusviricota MAGs共包含22,242个基因,分为35个核心基因簇,至少有50%的MAGs存在;1,825个非核心基因簇;最后是9,018个在GOEV数据库中没有近亲的singletons。核心基因簇提供了一个窗口,让人们了解到Mirusvirus各亚族之间共享的关键功能。除了上述病毒模块和信息模块的核心成分外,它们还对应着与DNA稳定性(H3组蛋白)、DNA复制(DNA复制许可因子、谷胱甘肽/核苷酸还原酶、Holliday接头分解酶和3′修复外切酶1)、转录(TATA结合蛋白)、基因表达调节(赖氨酸特异性组蛋白脱甲基酶1A)有关的功能、 RNA的转录后修饰(RtcB-like RNA-splicing ligase)和蛋白质的转录后修饰(putative ubiquitin protein ligase),蛋白质降解(trypsin-like,C1和M16-family peptidases),细胞生长控制(Ras相关蛋白),检测外部信号(sensor histidine kinase)和光敏感受体蛋白(heliorhodopsins)。因此,Mirusvirus编码了一个精心设计的工具包,可以实现对其宿主的细胞生物学和能量潜力进行微调,以实现最佳的病毒复制。最后,根据与参考数据库中的蛋白质的序列或结构比较,10个核心基因簇不能被赋予任何功能,有待于进行实验性的功能鉴定。
根据基因簇的定量出现,对Mirusviricota MAGs和来自培养的参考病毒基因组(包括核质病毒、疱疹病毒和Caudoviricetes)进行聚类,突出了Mirusviruses和疱疹病毒之间强烈的功能区分,反之,Mirusviruses和核质病毒之间有强烈的功能相似性。因此,从功能上讲,与疱疹病毒相比,Mirusviruses更接近于核质病毒(其中许多病毒也广泛存在于海洋表面,见图1)。为了进一步探索感染真核生物的海洋病毒的功能景观,我们利用整个GOEV数据库,根据基因簇的定量出现情况对其基因组进行了聚类。Mirusviruses聚在一起,并根据系统发育信号进一步组织成亚支系。相比之下,该分析强调了核质病毒的复杂功能构成,一些支系(如Imitervirales和Algavirales)被分成多个群。除了信息模块的核心成分外,连接Mirusviricota和Nucleocytoviricota基因组相当大一部分的基因簇以参与DNA复制的功能为主:谷氨酰胺/核苷酸还原酶、Holliday接头解析酶、增殖细胞核抗原、dUTP酶和DNA拓扑异构酶II。共同分享的功能还包括Ras蛋白、patatin样磷脂酶(脂质降解)、肽酶C1、泛素羧基末端水解酶(蛋白质活性调节)和Evr1/Alr家族(细胞膜Fe/S蛋白的成熟)。因此,这两个门类之间的功能连接远远超出了信息模块的范围。另一方面,数以百计的基因簇和功能在Mirusviruses或核质病毒群中明显富集,展现了这两个类群的不同生活方式。Mirusviruses中的核心基因簇在核质病毒基因组中的代表性明显较低的包括类胰蛋白酶(Mirusviruses中占73%,核质病毒中占9%)和M16家族(60%对2%)肽酶、TATA结合蛋白(59%对0%)、heliorhodopsin(64%对5%)和组蛋白(54%对2%)。组蛋白和heliorhodopsin的系统发育推论指出,这些基因在Mirusviruses和核质病毒都有复杂的进化历史,并且病毒支系和海洋浮游真核生物之间有多次水平转移事件。此外,Micromonas的一种heliorhodopsin可能来源于一种Mirusviruses,这表明Mirusviruses与核质病毒一起,通过基因流的方式对浮游真核生物的进化做出了贡献。
Mirusviruses是丰富和活跃的
据我们所知,Mirusviricota是Duplodnaviria中第一个被发现在阳光下的浮游生物中广泛而丰富的真核生物感染谱系。事实上,在Tara Ocean的143个站点中,有131个站点检测到Mirusviruses,包括从北极到南极的站点。它们主要出现在0.2-5微米(占整个Mirusviruses宏基因组信号的76.3%)和3-20微米(15.4%)大小的组分,涵盖了单细胞浮游真核生物的高度多样性(图1和2)。在我们研究中所考虑的Tara Ocean宏基因组中,海洋核质病毒MAGs和培养基因组在GOEV中的总平均覆盖率比Mirusviruses MAGs高15倍,反映了目前这两个门类之间基因组单位的不平衡(1,706对111)。然而,与所有核质病毒门的目相比,Mirusviruses的中位累积平均覆盖率较高,但Algavirales病毒目明显例外。因此,Mirusviruses 是迄今为止在阳光下的海洋中最丰富的真核生物病毒之一。
Mirusviruses 不仅丰富,而且在浮游生物中也非常活跃。事实上,在GOEV中仅包含3.8%基因的Mirusvirus MAGs,在这个基因组数据库中却占Tara Ocean宏转录组信号的13%。这种大量的原位转录组信号强调了Mirusviricota与海洋系统中真核生物病毒-宿主动态的相关性。与中深海区(深度大于200米)相比,Mirusviruses 在阳光下的海洋(尤其是富含叶绿素的透光层)最为活跃,在0.2-20微米的细胞范围内(图3),这与宏基因组的信号一致。Mirusviricota的35个核心基因簇占宏转录组信号的20%(包括12%的7个衣壳蛋白),其余信号与非核心基因簇(43%)和单基因(37%)有关。因此,高度多样化的基因(近10,000个单基因被鉴定出来)似乎在海洋真核微生物感染过程中对Mirusviricota的功能活动起着关键作用。
Mirusviruses有不同的生物地理分布(例如,有些只在北冰洋发现),然而它们的35个核心基因在有宏转录组信号的样本中表达水平相似,表明无论纬度或亚支系如何,都有相对同质的功能生活方式(图3)。表达水平最高的是编码衣壳蛋白的基因,其比例再现了疱疹病毒衣壳中相应蛋白的比例(例如,与三联蛋白或门户蛋白相比,HK97 MCPs更多)。编码新型heliorhodopsin和组蛋白的基因也有较高的表达量,这表明在感染期间有重要的功能作用。总的来说,Mirusviruses的生物地理学和原位转录组模式表明,它们在温带和极地水域都积极感染丰富的海洋单细胞真核生物。
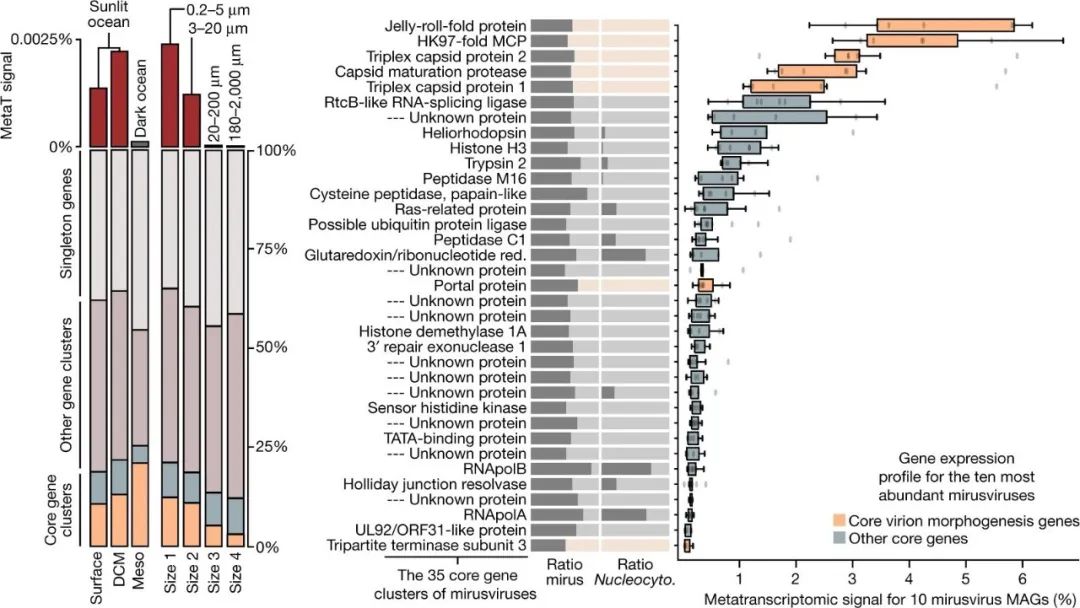
图3:感染期间Mirusviruses的原位表达概况。
左图:Tara Ocean宏转录组中不同基因类别的Mirusviruses MAGs的整体宏转录组信号总结。DCM,深层叶绿素最大值层;Meso,中层(200米以下的顶部暗海层)。右图,35个Mirusviricota核心基因簇的出现比例汇总,为Mirusvirus MAGs(mirus)比上Nucleocytoviricota(Nucleocyto.)的比例。该面板还显示了对应于35个核心基因簇的基因和在Tara Ocean宏基因组中10个最丰富的Mirusviruses中出现的整体宏转录组信号的框图。百分比值是以基因组为中心的,当考虑到在相应的基因组中发现的所有基因的累积平均覆盖率(所有宏转录组的总和)时,对应于一个基因的平均覆盖率的百分比(所有宏转录组的总和)。框图中的中心线表示中位数;框限表示第25和75个百分位数;晶须从第25和75个百分位数延伸到1.5倍的四分位数范围;离群值用点表示(n = 10点)。Red.,还原酶。
Mirusviruses连接两个病毒域
为了进一步验证Mirusviruses的基因组内容,并排除人为杂交的可能性,我们为新发现的Mirusviricota MCP创建了一个HMM,并以它为诱饵在其他数据库中搜索完整的基因组。首先,我们在表层海洋中尺寸小于0.2微米的部分的综合病毒基因组资源(全球海洋病毒组2,GOV2)中只发现了两个Mirusviricota MCP,这表明该支系的大多数病毒尺寸大于0.2微米。我们随后在一个包含数百个来自表层海洋0.2-3微米大小部分的宏基因组组合的数据库中筛选出了Mirusviricota MCP。我们在地中海发现了一个连续的Mirusviricota基因组(355个基因),隶属于M2支系,长度为431.5 kb,仅比最长的Mirusviricota MAG短6 kb(图2b,c)。它的基因再现了Mirusviruses的核心功能(例如拓扑异构酶II、TATA结合蛋白、组蛋白、多种日光蛋白、Ras相关GTP酶、细胞表面受体、泛素和胰蛋白酶),与Nucleocytoviricota HMMs相比,其中80个基因有明显的命中率。最关键的是,不仅Mirusviricota的信息模块(DNApolB、RNApolA、RNApolB和TFIIS)和病毒模块(HK97-fold MCP、终止酶、门户蛋白、壳成熟蛋白酶和两个三重壳蛋白)的所有标志基因都存在,而且它们在整个基因组中也相对均匀地出现。因此,这个近乎完整的连续基因组完美地再现了仅在Mirusviruses和疱疹病毒之间共享的标志性病毒模块特征,以及Mirusviruses和核质病毒之间共享的信息模块(图4)。
一方面,根据其病毒模块,Mirusviruses属于Duplodnaviria域。另一方面,它们的标志性信息基因在核质病毒门中普遍存在同源物,序列相似度之高出乎意料。这些结果有力地表明,这种信息模块起源于巨型病毒(巨型病毒起源假说;图4b)或Mirusviruses(Mirusviruses起源假说;图4c),然后在它们两个域之间转移,很可能是在病毒和原真核生物宿主之间相应基因的长期共同进化之后。因此,Mirusviruses不仅是真核浮游生物生态学的组成部分,而且还填补了我们对双链DNA病毒的两个主要域的进化轨迹的关键空白。
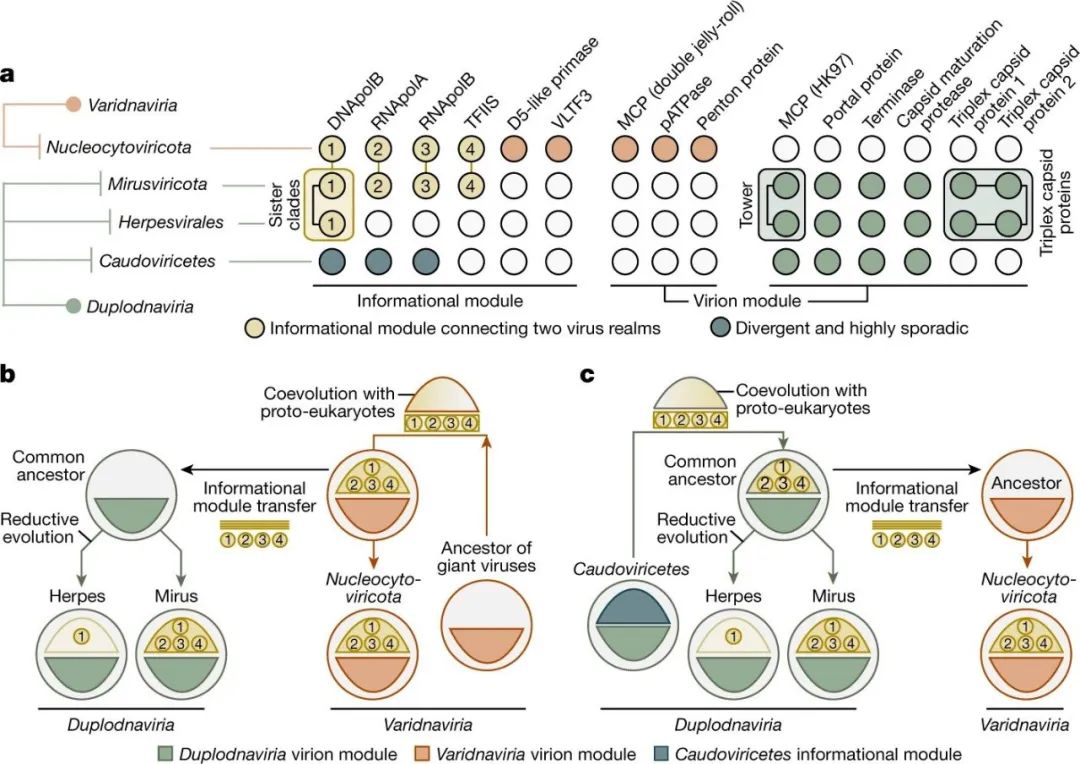
图4:真核信息模块的进化轨迹。
a,核质病毒、Mirusviruses、疱疹病毒和Caudoviricetes的信息模块和病毒模块的标志基因的出现摘要。具有强烈进化关系的信息模块基因用一条线连接。含有指向Mirusviruses和疱疹病毒之间共同的真核生物病毒祖先的信息的基因被框起来。b,c, 描述了两种进化情况,在这两种情况下,Duplodnaviria和Varidnaviria域内的真核生物感染病毒的信息模块首先出现在Nucleocytoviricota(巨型病毒假说)或Mirusviruses(Mirusviruses假说)的祖先。
- 讨论-
我们对五个大洋和两个海洋表面的浮游生物进行了系统发育指导的基因组解析宏基因组调查,发现了一个大型真核DNA病毒的主要支系,其基因组的长度可以达到400 kb以上,它们在阳光下的海洋中是多样的、普遍的、活跃的。这个被称为Mirusviricota的支系对应于Duplodnaviria域中的一个假定的新门,到目前为止只包括感染细菌和古细菌的Caudoviricetes和感染动物的Herpesvirales。Mirusviricota门至少分为七个亚支系,可能对应于不同的科。尽管Mirusvirus和疱疹病毒都是感染真核细胞的双链DNA病毒,但它们表现出非常不同的基因组特征。最值得注意的是,Mirusvirus大大偏离了以前所有其他特征的DNA病毒组,其病毒形态发生模块(最高级双链DNA病毒分类的决定性特征)隶属于Duplodnaviria域,而信息模块则与Varidnaviria域内的大型和巨型病毒密切相关。这些明显的奇特属性在一个431.5kb的近乎完整的连续基因组中得到了重现。Mirusviricota的发现提醒我们,我们还没有掌握关键生态系统(如我们的海洋表面)中最丰富的双链DNA病毒的全部生态和进化复杂性。
Mirusviruses在阳光下海洋的各个区域相对丰富,它们积极感染尺寸小于20微米的真核浮游生物,并表达各种功能。Mirusviricota有一个统一在一起的、复杂的推断生活方式,包括独特的特征(许多核心基因只在这个门中发现),但也与大型和巨型真核varidnavirus病毒的生活方式有很大的重叠。这些共同的功能远远超出了信息模块,包括生态系统和宿主的特定基因,这些基因可能是在这两组病毒之间横向转移的,也可能是在进化过程中的不同时间点从共同的宿主那里通过趋同进化获得的。例如,两个门之间共享的蛋白酶(patatin-like phospholipase)已经被认为可以促进核质病毒基因组向细胞质和细胞核的运输。与核质病毒相比,Mirusviruses中富集的功能包括系统发育不同的H3组蛋白(参与真核细胞内染色质形成的蛋白)和heliorhodopsins(光敏受体蛋白,在感染期间可被巨大病毒用作质子通道)。总之,生物地理学模式、功能基因汇辑和宏转录组信号表明,Mirusviruses利用一种以前被忽视的生活方式影响着关键海洋真核生物的生态。
疱疹病毒目和核质病毒门的病毒分别属于两个古老的病毒谱系,即Duplodnaviria和Varidnaviria,其相应的祖先可能早于最后的普遍细胞祖先(LUCA)。然而,这些突出的感染真核细胞的双链DNA病毒支系的确切进化轨迹和各自最新的共同祖先的身份仍然难以捉摸,部分原因是缺乏已知的中间状态。特别令人费解的是无处不在的Caudoviricetes和Herpesvirales之间的差距,前者在功能的复杂性和基因重组的丰富性方面可与Nucleocytoviricota相媲美,后者仅限于动物宿主,并且缺乏转录机制和进行核复制。Mirusviricota的鉴定将Duplodnaviria的存在范围从动物扩大到真核浮游生物宿主,强烈地表明它们与真核生物的早期进化联系。在Mirusviricota和Herpesvirales中塔状结构域的存在和位置与两个三倍体外壳蛋白的保存相结合(这些都不存在于已知的Caudoviricetes中)(见图1)强烈地表明这些真核生物病毒的共同祖先,而不是从不同的Caudoviricetes支系独立进化而来。Mirusviricota信息基因的深分支定位证明了一次或多次早期历史转移(图1),以及与其他DNA病毒支系相比,两个真核Duplodnaviria支系之间DNApolB的密切相似性提供了补充信息。由于塔状结构域的尺寸较短,并考虑到与单细胞真核生物相比,动物的出现时间较晚,Mirusviricota可能更接近感染真核生物Duplodnaviria的祖先状态。因此,Mirusviruses指向疱疹病毒的浮游祖先;疱疹病毒可能经历还原性进化,最明显的是失去转录机制,并专门用于感染动物细胞。
同样令人费解的是大型和巨型核质病毒门基因组与相对简单的Varidnavirus之间的进化渠道,这些Varidnaviruses的病毒体形成和基因组复制的基因组合不多(那些感染细菌和古菌的变种病毒,以及噬病毒体、腺病毒科,或其他雅拉病毒和多角体病毒)。有人推测,其中一些简单的Varidnaviruses可能代表了噬菌体和来自核质病毒门的真核巨型病毒之间的进化中间产物。浮游生物内的Mirusviruses的基因组复杂性,以及它们与核质病毒门共享的核心功能提供了进一步的见解。信息模块,可能还有其他功能,可能已经从核质病毒转移到了Mirusviruses的祖先(巨型病毒起源假说),促进了感染真核生物Duplodnavirus的复杂化。在这种情况下,核质病毒可能将其病毒模块与共同感染同一宿主的未定性的Duplodnavirus的病毒模块互换,同时保留了精心设计的信息模块。然而,我们的数据并不排除同样发人深省的可能性,即信息模块从一种Mirusviruses转移到核质病毒更简单的祖先(Mirusviruses起源假说)。这种情况可以帮助解释从 "小 "Varidnaviruses到极其复杂的核质病毒的进化飞跃。不管考虑的是哪种假说,Mirusviruses都阐明了两个域的真核双链DNA病毒的进化轨迹。
总的来说,Mirusviruses的普遍性、功能的复杂性和经过验证的转录活性表明,Mirusviruses在海洋生态系统的生态学中起着突出的作用。这个假定的门不仅扩大了我们对浮游生物生态学的理解,而且还提供了对病毒进化的新见解。尽管Mirusviruses可能早于疱疹病毒的出现,但Mirusviruses在浮游生物中的起源时间线(在巨型真核病毒出现之前或之后)还没有被阐明。今后,更多的功能和基因组特征,加上培养和环境细胞分选以确定宿主,将进一步促进我们对Mirusviruses在海洋和其他地方的生活方式和突出地位的评估。
- 深度解析 -
作者利用在全球五个大洋和两个海洋表面的Tara Ocean的宏基因组数据集,以RNApolB(核质病毒门specific的informational protein)为指针调取了潜在的核质病毒门病毒。发现了两类新的类群,一个为Proculviricetes类群(极地类群),仅包括北冰洋和南大洋的少数几个病毒基因组,另一个为Mirusviricota,分布广泛,表达活性高,多样性丰富。文章以四个核质病毒门的informational protein为标志序列分析了Mirusviricota的系统发育定位,发现了这一类群与属于核质病毒门多个目的谱系有亲缘关系,但独立分支开来。Mirusviricota被发现有嵌合的基因组特性,它的informational module(信息模块)与核质病毒门相近,但是virion module(病毒模块)与疱疹病毒相近。Mirusviricota的衣壳MCP是与疱疹病毒(属于Duplodnaviria域)相似的HK97折叠,并且病毒模块的其他关键组分也属于Duplodnaviria,这表明Mirusviruses是属于Duplodnaviria域的一类新病毒。同时Mirusviricota的MCP反应出和四个信息模块蛋白一样的系统发育分支,这表明Mirusviruses的病毒模块和信息模块是一起进化的,是从进化祖先那里共同得到并流传下来的。
然而在功能上面,Mirusviruses却和核质病毒更相近;连接它们的除了信息模块的蛋白以外,还有一系列关于DNA复制功能的蛋白。其中组蛋白和heliorhodopsin的进化分析表明,这两个蛋白在Mirusviruses和核质病毒中都有复杂的进化历史,其中包含有与海洋浮游真核生物之间的多次水平基因转移事件。比如,一种单细胞的海洋绿藻中的heliorhodopsin可能来源与Mirusviruses。这说明在长期的进化历史当中,Mirusviruses和核质病毒可能和它们的宿主或者密切接触的邻居产生过基因的交换。
转录组的数据表明Mirusviruses有与核质病毒目水平类群相近的表达水平,分布于不同纬度的海洋表层区域。同时,Mirusviruses的non-core genes和singleton genes的较广泛表达水平也表明这些基因的多样性可能在Mirusviruses感染海洋真核微生物活动中起到关键的作用。作者为了排除在病毒组binning中人为杂交的可能性,以MCP蛋白为诱饵钓取了0.2-3微米组分海洋宏基因组数据库,发现了一个连续的Mirusviruses基因组。信息模块和病毒模块均匀地在基因组中分布。这个连续的、近乎于完整的基因组再次验证了Mirusviruses的嵌合基因组特征,排除了人为引入的可能。
作者提出了信息模块水平转移的两种假说:(1)信息模块起源于巨型病毒(核质病毒)的祖先,然后再水平转移到Mirusviruses和疱症病毒的祖先,最后Mirusviruses和疱症病毒分别含有信息模块的一部分。(2)信息模块起源于Mirusviruses,然后再水平转移到巨型病毒的祖先。现在的Mirusviruses和疱症病毒分别含有信息模块的一部分。不过这两种假说共同点是信息模块都是病毒在和原真核生物相互作用的长期进化历史阶段以后形成的。
对于Duplodnavirus和Varidnavirus这两大域内真核DNA双链病毒的进化,Mirusviruses的发现都提供新的见解。在Duplodnavirus里面,Caudoviricetes和Herpesvirales(疱疹病毒)差异明显,前者有复杂的功能和重组特性,而后者是简单的感染动物类群的病毒。Mirusviruses和疱疹病毒相似的MCP结构证明,这两个真核生物感染病毒来自于同一个祖先。很可能,感染真核简单浮游生物的Mirusviruses在进化上先于疱疹病毒,而疱疹病毒经历了还原性进化,丢掉了转录机制,专门感染动物细胞。在Varidnavirus里面,核质病毒与其他简单的Varidnavirus有进化的“鸿沟”。在Mirusviruses起源假说里面,核质病毒可能有比较简单的祖先,信息模块转移后,核质病毒发展成现在拥有精细的信息模块和功能的复杂病毒。这种假说可以解释Varidnavirus内简单病毒到复杂的核质病毒的进化转变。
信息模块水平转移的具体情况有待于今后的研究阐明。同时Mirusviruses和核质病毒在感染浮游生物的进化历史中出现的先后顺序有待以后解决。同时,Mirusviruses的具体宿主情况也不太清楚。这些都依赖于以后细胞培养、基因组/转录组研究和功能特征鉴定等方面的手段来研究解决。
参考文献
Gaïa, M., Meng, L., Pelletier, E. et al. Mirusviruses link herpesviruses to giant viruses. Nature (2023). https://doi.org/10.1038/s41586-023-05962-4
- 作者简介 -
通讯作者

法国国家科学研究中心
Tom O. Delmont
研究员
Tom O. Delmont,法国国家科学研究中心、巴黎大学-萨克雷分校François Jacob研究所,微生物生态研究员, TARA Oceans consortium欧洲员,前芝加哥大学anvi'o team(A. Murat Eren (Meren) team)成员,全球海洋系统生态学和进化研究联合会成员。
猜你喜欢
iMeta简介 高引文章 高颜值绘图imageGP 网络分析iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文

















 2265
2265

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









