






 点击蓝字↑↑↑“微生态”,轻松关注不迷路
点击蓝字↑↑↑“微生态”,轻松关注不迷路
编译:微科盟小舍,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
固氮微生物通过缓解氮限制来调节海洋生产力。迄今为止,化学自养细菌被广泛认为是寡营养海洋和陆地生态系统中的主要的固氮菌。然而,在富含有机质的生境中,化学自养生物对固氮作用的贡献尚不清楚。本研究采用宏基因组学和宏转录组学方法,结合培养实验,研究了漳州红树林沉积物中的固氮菌的多样性、分布和活动。理化分析表明,所研究的红树林沉积物是典型的富碳、富硫、氮限制和低氧化还原的海洋生态系统。这些沉积物中含有广泛的固氮酶基因系统发育变种,包括类群I-III和VII-VIII。出乎意料的是,包括弯曲菌(Campylobacteria)、γ-变形菌(Gammaproteobacteria)、ζ-变形菌(Zetaproteobacteria)和Thermodesulfovibrionia在内的多种化能自养分类群是0-18 cm沉积物层中主要的和活跃的固氮菌。相比之下,18-20 cm层以化学自养分类群脱硫菌门和Halobacteriota的活性固氮菌为主。对MAGs的进一步分析表明,主要的化能自养菌可以通过耦合氢、还原硫和铁的氧化,与氧、硝酸盐和硫还原来固定氮。培养实验进一步证明了,化能自养弯曲菌的成员具有氢和硫氧化驱动的固氮能力。活性测定证实,栖息在红树林沉积物中的固氮菌优先从各种还原性无机化合物中吸收能量,而不是从有机物中吸收能量。总体而言,本研究结果表明,在红树林沉积物中,化能自养菌而非异养菌是主要的固氮菌。本研究强调了化能自养生物在碳主导的生态系统中的重要性。
论文ID
原名:Chemolithoautotrophic diazotrophs dominate dark nitrogen fixation in mangrove sediments
译名:化能自养固氮菌在红树林沉积物中主导暗氮固定作用
期刊:The ISME Journal
IF:10.8
发表时间:2024.6
通讯作者:邵宗泽、董西洋、姜丽晶
通讯作者单位:自然资源部第三海洋研究所
DOI号:10.1093/ismejo/wrae119
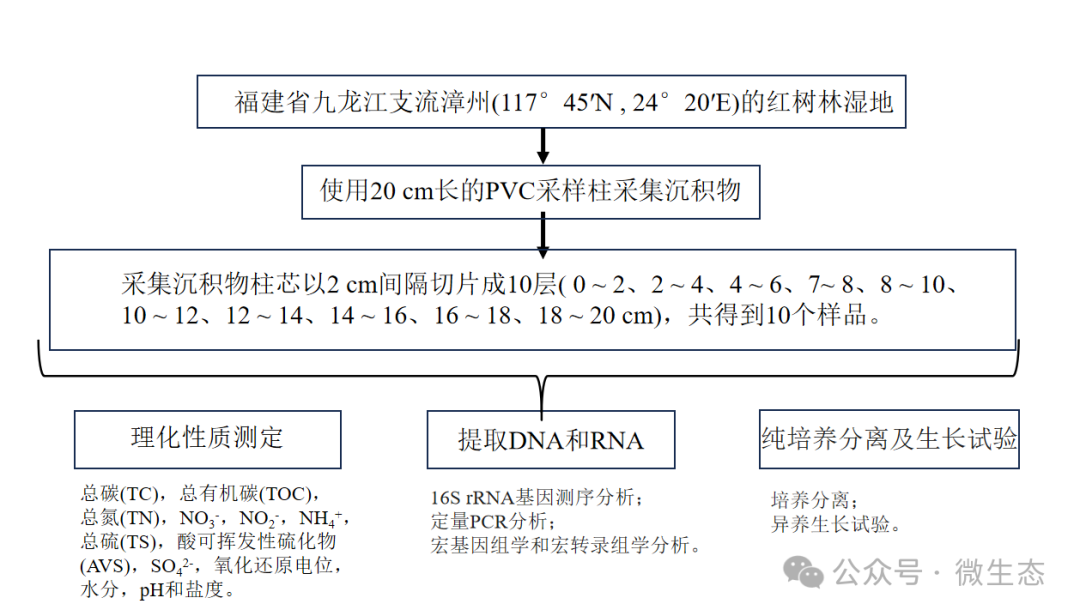
实验设计
结果和讨论
1 红树林沉积物的理化特征
本研究测定了漳州红树林沉积物中总碳(TC),总有机碳(TOC),总氮(TN),NO3-,NO2-,NH4+,总硫(TS),酸可挥发性硫化物(AVS)、SO42-,氧化还原电位、水分、pH和盐度,以表征其在0 ~20 cm深度的垂直分布(图S3)。在所有深度都观测到高浓度的TC和TOC(2.41~2.59%和2.10~2.35%),而TN浓度较低,范围为0.12~0.14% (图S3A-C)。碳氮比在0~20 cm层的变化范围为18.39~21.97,与其他红树林沉积物生态系统相似,而与潮滩、微咸水和淡水生境中的碳氮比不同。无机氮包括NO3-(0.14~0.65 mg/kg)和NO2-(0.01~0.04 mg/kg)的浓度较低,且随深度的增加而降低(图S3D,E)。各深度NH4+浓度变化范围为1.27~3.23 mg/kg(图S3F)。在其他红树林生境中也报道了类似的低铵态氮浓度,强调了漳州红树林表层沉积物中的氮限制。
TS和AVS的浓度随着沉积物深度的增加而增加,在18~20 cm层(图S3G,H)处达到峰值,而SO42-浓度则呈现降低的趋势,在0~6 cm层(图S3I)浓度最高。硫代硫酸盐(S2O32-)仅在表层沉积物(在0-2 cm和2-4 cm层中分别为~40 μM和~20 μM)中检出。顶部0~2和2~4 cm氧化还原电位(Eh)分别为320和252 mV,在4 cm以下随深度急剧降低,最深层(图S3J)处达到最低值-120 mV。在所有测量参数中,TN、NO3-、NO2-和SO42-与深度(P<0.05)呈负相关,而TS和AVS与深度(P<0.05)呈正相关(图S4)。总的来说,这些发现强调了漳州红树林表层沉积物的特点是碳和氮的不平衡,同时还含有丰富的还原性硫化物。
2 不同沉积物深度中固氮基因的多样性、分布和活性
nifH基因编码一种必需的固氮酶蛋白,常用于固氮菌在不同环境中的多样性和流行率研究。从红树林沉积物样品中提取的宏基因组组装的连续克隆系注释显示,共有154个非冗余的nifH同系物属于固氮酶超家族(图1A;表S1)。除了类群IV到类群VI的类固氮酶外,这些同源物被归类到典型的固氮酶序列(Ⅰ~Ⅲ类群)和两个新提出的谱系,即Ⅶ和Ⅷ类群(图1A)。基于固氮酶保守基序的分析,两个新的谱系也被认为是真正的nifH,详见下文。这些结果表明红树林沉积物比以前认为的的红树林沉积物含有更多样的固氮酶基因。
 图1 .红树林沉积物中154个固氮酶亚基NifH的非冗余同源物的概述。A,NifH氨基酸序列的最大似然法系统发育树,将nifH同系物分为典型类群I~III,固氮类类群IV~VI,以及新归属的类群VII和VIII。本研究获得的NifH序列用蓝色高亮显示,重氮营养型MAGs序列用恒星标记。标尺栏表示平均每个站点有替换。B,不同分类群nifH基因的相对丰度(左)和不同红树林沉积物样品中基因丰度的总值(右)。C,不同分类群nifH基因的相对转录本丰度(左)和不同沉积物样品中转录本表达量的总值(右)。不同分类群以不同颜色为代表。“其他”类别代表未分配的序列。
图1 .红树林沉积物中154个固氮酶亚基NifH的非冗余同源物的概述。A,NifH氨基酸序列的最大似然法系统发育树,将nifH同系物分为典型类群I~III,固氮类类群IV~VI,以及新归属的类群VII和VIII。本研究获得的NifH序列用蓝色高亮显示,重氮营养型MAGs序列用恒星标记。标尺栏表示平均每个站点有替换。B,不同分类群nifH基因的相对丰度(左)和不同红树林沉积物样品中基因丰度的总值(右)。C,不同分类群nifH基因的相对转录本丰度(左)和不同沉积物样品中转录本表达量的总值(右)。不同分类群以不同颜色为代表。“其他”类别代表未分配的序列。
从表层到18 cm,nifH基因丰度在20.1~34.1个/百万(GPM)之间,峰值为18~20 cm层的47.8个GPM(图1B)。在门水平上,红树林沉积物中最丰富的假定固氮菌分别为脱硫杆菌(Desulfobacterota)、假单胞菌(Pseudomonadota)和弯曲菌(Campylobacterota)(图1B)。在纲水平上,γ变形菌(Gammaproteobacteria)、弯曲菌(Campylobacteria)和脱硫杆菌(Desulfobacteria)是最主要的纲(图S5A)。尽管在14-16 cm层没有转录本,但在其他层检测到nifH转录本,表明大多数沉积物深度都存在固氮活性(图1C)。在nifH转录本高的4个层中,弯曲菌(Campylobacterota)、硝化螺旋菌(Nitrospirota)和假单胞菌(Pseudomonadota)出现了潜在的优势固氮类群(图1C)。具体来说,这些群落之间存在生态位分化。在6-8和10-12 cm层,nifH转录本主要隶属于弯曲菌类(图S5B),在12-14 cm层,nifH转录本在Thermodesulfovibrionia类中高表达。在16-18 cm层中,nifH转录本在γ变形杆菌和弯曲菌中的数值较高(图S5B)。
考虑到占优势且具有转录活性的固氮菌主要是属于化能自养的弯曲菌门和硝化螺旋菌,因此本研究进一步探索,以确定nifH与红树林沉积物中碳固定基因(如,aclB、rbcL和acsB)、硫氧化基因(如,sqr和soxB)或氢氧化基因(hydB)之间的相关性。结果表明,nifH的转录活性与上述基因之间存在强而显著的相关性(图S6),这表明在红树林沉积物中固氮菌和假定的化能自养种群之间存在着牢固的联系。因此,假设这些化能自养微生物可能构成红树林沉积物原位固氮的关键活性类群。
3 在红树林沉积物中鉴定出潜在的固氮微生物
通过宏基因组组装和分箱策略,本研究恢复了180个细菌(n=172)和古菌(n=8)群体基因组,其完整性>50%,污染率<10%,基于GTDB分类学(表S2),分属于24个门。在这些基因组中,36个MAGs(33个细菌和3个古菌MAGs)跨越12个门,被鉴定为潜在的固氮微生物,包括脱硫菌门(n=10)、Pseudomonadota(n=6)、Campylobacterota (n=4)、Chloroflexota (n=3)、Halobacteriota (n=3)、Myxococcota (n=3)、Bacteroidota (n=2)、Methylomirabilota (n =1)、Nitrospirota (n=1)、Spirochaetota (n=1)、Schekmanbacteria (n=1)和SZUA-182 (n=1)(图2A;表S3)。系统发育分析显示,这36个MAGs的固氮酶被分为I、II、VII和VIII类群(图2B),所有这些MAGs在NifH、NifD和NifK(图S7-9)之间具有保守的活性位点。此外,来自相同类群的NifH序列并没有聚成一个单独的分支(图2A,B),这可能是由水平基因转移(HGT)解释的。
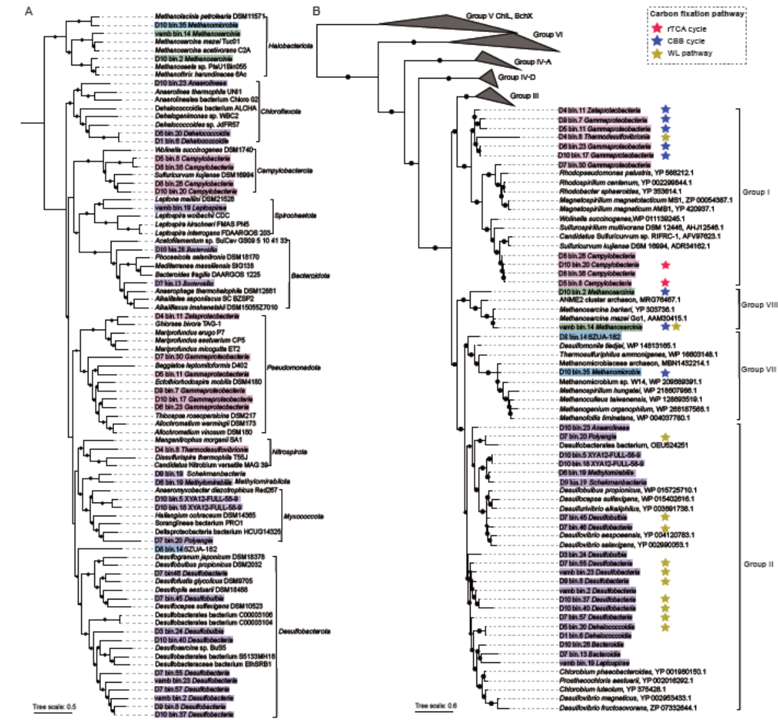 图2 固氮MAGs及其NifH蛋白序列的系统发育树。A,基于保守蛋白序列的串联,构建了包含固氮基因的36个MAG的最大似然法系统发育树。MAG根据其在门水平上的NifH同源群进行颜色编码。B,NifH蛋白序列在MAGs内的最大似然法系统发育树。不同颜色的星表示不同的固碳途径,rTCA为红,CBB为蓝色,WL为黄色。对于两棵树,在节点处表示bootstrap值大于50%,标尺条表示平均每个站点的替换数。
图2 固氮MAGs及其NifH蛋白序列的系统发育树。A,基于保守蛋白序列的串联,构建了包含固氮基因的36个MAG的最大似然法系统发育树。MAG根据其在门水平上的NifH同源群进行颜色编码。B,NifH蛋白序列在MAGs内的最大似然法系统发育树。不同颜色的星表示不同的固碳途径,rTCA为红,CBB为蓝色,WL为黄色。对于两棵树,在节点处表示bootstrap值大于50%,标尺条表示平均每个站点的替换数。
在36个潜在的固氮MAGs中,大多数(21/36)编码碳固定途径,至少有60%的基因和所有的关键酶(图2B;S10)。还原柠檬酸(rTCA)循环由弯曲菌(图2B;S11)门的弯曲菌纲(n=2)编码。Calvin Benson-Benson-Bassham (CBB)循环由5个假单胞菌MAGs编码,包括γ变形杆菌纲(n=4)和ζ-变形菌纲(n=1),以及3个嗜盐菌MAGs包括甲烷八叠球菌纲(n=2)和甲烷微菌纲(n=1)(图2B;S12)。Wood-Ljungdahl (WL)途径由8个脱硫菌门 MAG编码,包括Desulfobacteria (n= 6)和Desulfobulbia (n=2),1个Myxococcota MAG编码,包括Polyangia (n=1),1个Nitrospirota MAG编码,包括Thermodesulfovibrionia (n=1),HalobacteriotaMAG编码,包括甲烷八叠球菌纲 (n=1) (图2B;S13)。这些发现表明,这些MAGs所代表的微生物可能是潜在的化能自养固氮菌。
对这36株固氮菌的Read mapping分析表明,它们广泛分布于不同的沉积物深度(图3A;表S4)。当单独考虑时,化能自养分类单元(Campylobacterota)在除18-20 cm层外的沉积物深度中最丰富(图3A),表明其在红树林沉积物中作为潜在固氮菌的重要作用。前期研究表明,弯曲菌在表层(0~15 cm)红树林沉积物中大量存在,耦合硫氧化和反硝化过程。此外,假单胞菌门(Pseudomonadota)和化能自养硝化螺旋菌门(Nitrospirota)在0-18 cm层中也表现出较高的丰度,但随着沉积物深度的增加,丰度持续下降(图3A)。而在18-20 cm层,脱硫菌门、Myxococcota门和Halobacteriota门的成员是最主要的固氮菌(图3A)。这些高丰度可能指示了这些门在深层沉积物固氮中的重要作用。最近研究表明,在30 cm深的红树林沉积物中,脱硫菌门和嗜盐菌门为优势菌。在纲水平上,弯曲菌在0-18 cm层显著占优势,其次是Desulfobacteria、Gammaproteobacteria、Dehalococcoidia和Thermodesulfovibrionia等纲,而在18-20 cm层,Desulfobacteria、YA12-FULL-58-9和甲烷微菌纲(Methanomicrobia)等纲的丰度最高(图S14A)。因此,在红树林表层0~20 cm沉积物中,化能自养生物始终作为优势固氮菌出现。
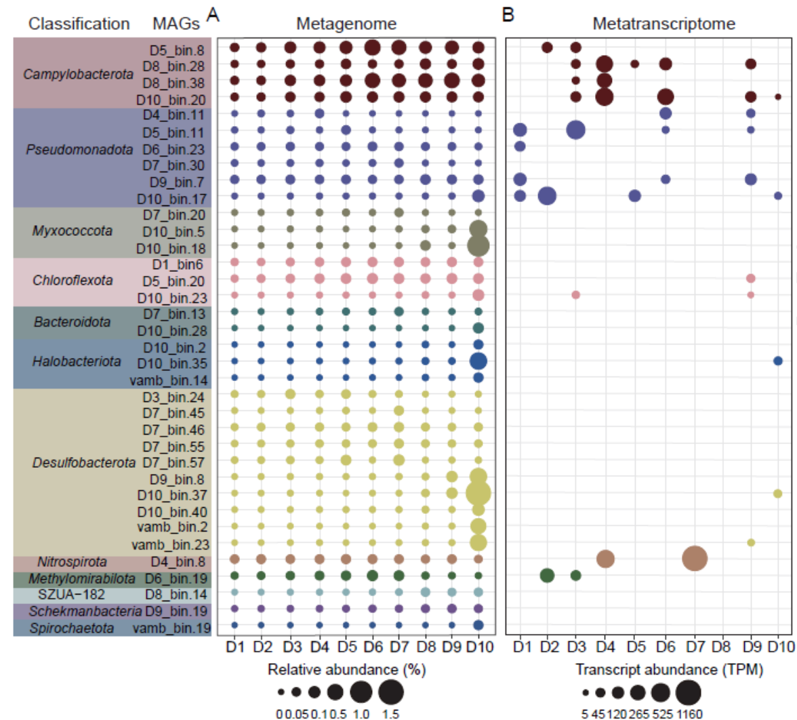 图3 36个固氮MAGs转录本在不同红树林沉积物深度的相对丰度和表达量。A,使用Cover M估算每个MAG的相对丰度。B,每个MAG的转录本表达量,单位为每百万个转录本(TPM)。详细信息为,见表S4和表S5。
图3 36个固氮MAGs转录本在不同红树林沉积物深度的相对丰度和表达量。A,使用Cover M估算每个MAG的相对丰度。B,每个MAG的转录本表达量,单位为每百万个转录本(TPM)。详细信息为,见表S4和表S5。
为了评估这些固氮菌的原位表达,来自不同沉积物深度的10个宏转录组与编码固氮酶的MAGs进行了比对(表S5)。结果表明,nifH基因在0-18 cm层的转录本,主要在Campylobacterota (n=4)、假单胞菌(n=5)和Nitrospirota (n=1)门的化能自养MAGs中表达(图3B),表明它们可能是红树林沉积物中的初级固氮菌。在纲水平上,弯曲菌的nifH基因在多个层从低到高转录水平,高达1456.38个转录本/百万reads (TPM),来自Thermodesulfovibrionia的转录本在6-8和12-14 cm层分别412.15和1122.25个TPM(图S14B)。此外,nifH转录本在多个沉积层的γ-变形菌纲中表达,在表层(0~6 cm)中高达,而在10-12 cm和16-18 cm层的ζ-变形菌中发现该转录本,分别为135.17和41.94 TPM(图S14B)。本研究是首次报道ζ-变形菌和Thermodesulfovibrionia具有活跃的固氮作用,表明他们可能在氮循环中发挥重要作用,除了他们先前报道的作用。在最深层的(18 ~20 cm)中,nifH转录本主要在脱硫菌门和Halobacteriota门中表达,但其值较低,分别为10.05 - 30.82和30.30 TPM (图3B),表明这些类群可能在更深层发挥较高的固氮作用。有趣的是,之前报道的异养固氮菌中没有表达nifH转录本,如粘球菌门(Myxococcota,n=2)、拟杆菌门(Bacteroidota,n=2)或螺旋体菌门(Spirochaetota,n=1)中的某些MAGs (图3B)。总体而言,本研究结果进一步证实了化能自养微生物主导了红树林沉积物中的暗固氮作用。
4 优势化能自养固氮菌的能量生产途径
为了预测优势化能自养固氮菌的功能,基于标记基因和代谢通路(图4A;表S6-8),测定了MAGs的代谢潜力。在硫代谢方面,所有弯曲菌MAGs (D5_bin.8、D8_bin.38、D8_bin.28、D10_bin.20)都含有多个编码硫化物的基因:醌氧化还原酶(Sqr)(图4A;S15),它催化硫化物氧化为单质硫。在D10_bin.20中鉴定到一个近乎完整的Sox系统(Sox ACDXYZ),表明其具有氧化硫代硫酸盐的遗传潜力。D5_bin.8和D8_bin.28包含部分Sox系统(图4A;S16),可能是由于MAG完整性较低。此外,D5_bin.8和D10_bin.20含有编码亚硫酸盐脱氢酶(SorAB)的基因,表明它们可以将亚硫酸盐氧化成硫酸盐。对于氢代谢,4个MAG都含有第1组[NiFe]-氢化酶(Hyd和Hya),只有D10_bin.20含有第2组[NiFe]-氢化酶(Hup) (图4A;S17),这可能使具有利用氢气作为能源的潜力。关于电子受体,D5_bin.8,D8_bin.38和D10_bin.20含有氧利用基因,编码cbb3型细胞色素c氧化酶(CcoNOPQ)和caa3型细胞色素c497氧化酶(CoxAB) (图4A)。与在有氧条件下诱导的低亲和力的caa3-type氧化酶相比,cbb3-type氧化酶是一种高亲和力的末端氧还原酶,能够在微氧到缺氧条件下发挥作用。细胞色素的存在也可以作为残余O2的清除剂,用于O2/活性氧(ROS)的解毒,以保护O2敏感的蛋白。此外,参与单质硫还原的多硫还原酶(Psr)由4个MAG编码,表明它们在缺氧条件下具有潜在的硫还原能力。这些结果表明,弯曲菌可以利用还原性含硫化合物和氢气作为电子供体,氧气和单质硫作为终端电子受体,生成ATP用于固氮作用。
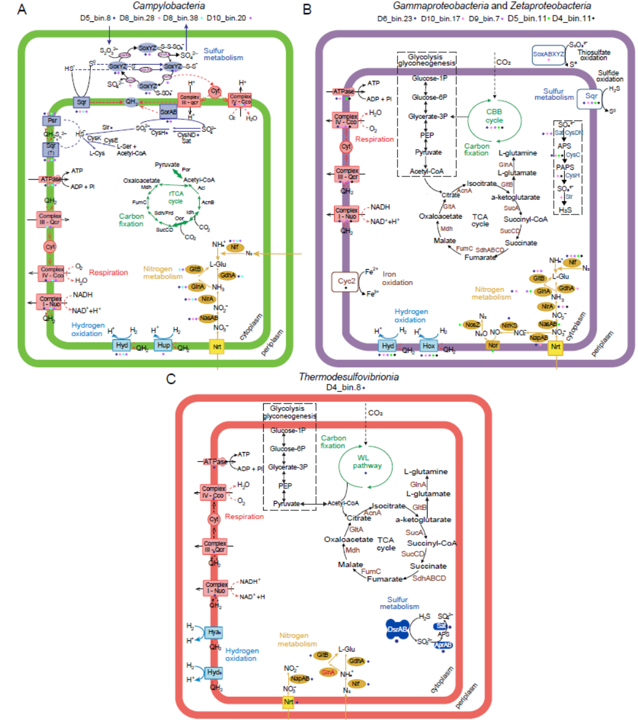
图4 优势固氮化能自养菌MAGs的代谢途径重建。推测代谢途径为弯曲菌属(Campylobacterota,A)、假单胞菌(Pseudomonadota,B)和硝化螺旋菌(Nitrospirota,C),固碳途径分别为rTCA、CBB和WL。具有多个箭头的步骤表明,操纵子编码不同的酶并催化该反应存在于基因组中。在这些重构中不存在的酶在red中被突出显示。全面的酶注释见表S6-S8。
1种MAG(D4_bin.8)属于Thermodesulfovibrionia,隶属于Nitrospirota(表S4)。对于硫代谢,D4_bin.8缺失了所有编码硫氧化的基因,如Sqr,Sox和Sor(图4C)。此外,D4_bin.8编码第1组[NiFe]-氢化酶(Hyd和Hya)和NAD-还原氢化酶(HoxHYU),用于氢氧化(图4C;S17)。相对于电子受体,D4_bin.8包含所有编码硫酸盐还原的基因途径,包括异化亚硫酸盐还原酶(DsrABC)和腺苷酸硫酸酯还原酶(AprAB) (图4C)。D4_bin.8还含有氧呼吸酶CcoNOPQ和CoxAB以及硝酸盐还原为亚硝酸盐酶NapAB。D4_bin中不存在还原单质硫的Psr。上述结果表明,Thermodesulfovibrionia可以将氢氧化与硫酸盐还原、反硝化或好氧呼吸耦合以获得固氮所需的能量,这与该纲的其他研究一致。
5 化能自养菌弯曲菌中潜在优势固氮菌的分离
从红树林沉积物中成功分离出10个菌株,分别命名为HSL-C5、HSL1-2、HSL1-6、HSL3-1、HSL3-2、HSL3-7、HSL- 3221、HSL-1716、HSL-1656和HSL1-3(图5A)。通过宏基因组学16S rRNA基因和(图S18;S19)扩增子测序,它们与弯曲菌(Campylobacterota)中的硫单胞菌属(sulfurimonas)和硫氧化菌属(sulfurovum)的成员具有最高的16S rRNA基因序列相似性(表S9),是原位红树林沉积物中的两个优势菌属。此外,基于两个属的ASVs和我们分离株的16S rRNA基因序列的系统发育树表明,菌株HSL1-3与来自Sulfurovum属的两个最主要成员ASV7(相对丰度高达6.07%)和ASV18 (丰度高达2.07%)代表的亲缘关系最近,序列一致性分别为97.14%和97.03%(图5;表S10)。菌株HSL3-7与硫单胞菌属中最优势的ASV3(丰度高达2.46%)形成一个簇,序列占99.03%。菌株HSL-3221对应于硫单胞菌属中第二优势的ASV19(上调~1.04%丰度),序列一致性为98.51%(图5A;表S10)。因此,这些分离物代表了原位红树林沉积物中硫单胞菌属和硫氧化菌属的优势菌。此外,通过对平均氨基酸一致性(AAI)的评估和基于基因组的系统发育分析,这10株菌属于5个属,包括硫单胞菌(图S20;S21)中的3个潜在新属。
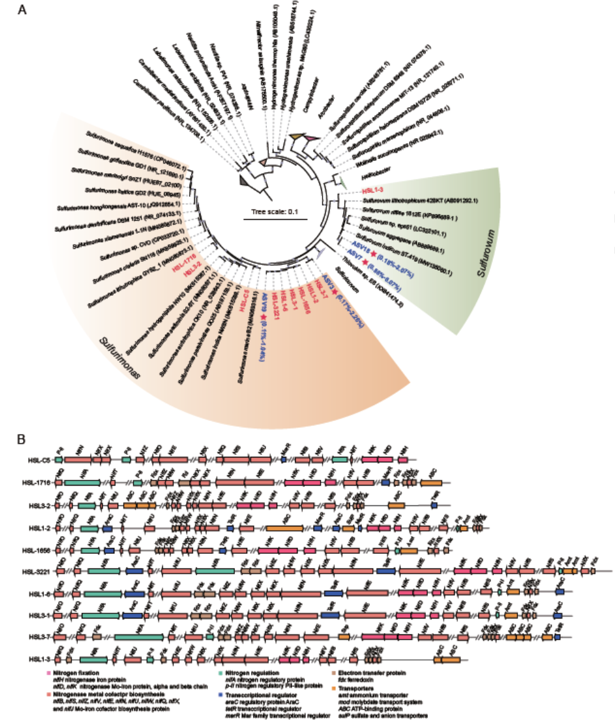
图5 弯曲菌(Campylobacterota)10个分离物的系统发育分析及固氮基因簇。A,基于16S rRNA基因序列的系统发育树来自Sulfurimonas和Sulfurovum属的代表ASVs和本研究中的分离物。在至少1个样本中,仅显示代表>1%群落的ASVs。刻度线代表每个位点的1.0个核苷酸替换。B,nifH DK的基因邻域包括固氮酶金属辅因子生物合成基因、调节固氮基因、转录调节基因、电子转移基因和转运蛋白基因。
10个菌株均含有一个完整的固氮基因簇,编码固氮酶,命名为NifHDKENB(图5B)。对这菌的基因邻近分析发现,在nif基因簇中编码电子传递蛋白、调节蛋白和固氮酶辅因子生物合成所必需的蛋白(图5B),这在之前的报道中也有发现。此外,生理特征显示,所有10个分离物都是专性化能自养菌,测试的有机化合物都不支持它们作为碳源和能源的生长。这10株菌中参与硫和氢的氧化以及各种终端受体还原的基因,如表S11所示,这些基因可以为固氮提供能量。事实上,固氮作用在化能自养弯曲菌中并不常见,因此描述了分离自地下原油储库中的Sulfuricurvum kujiense。因此,本文鉴定的含有固氮酶基因的弯曲菌(Campylobacterota)菌株可能在氮限制的红树林沉积物中具有竞争优势。与异养固氮菌相比,化能自养型固氮菌迄今为止培养较少。在本研究之前,仅从淡水、矿山废物、盐沼和温泉中分离到几株属于Pseudomonadota门和Aquificota门的细菌,它们可以利用还原性含硫化合物、氢、As或Sb作为能源固定N2进行化能自养生长。
6 菌株和红树林沉积物中的固氮活性
选取生长旺盛的7株活性菌株,利用15N标记同位素分析法测定其在细胞内固定15N2的能力。结果表明,这7株菌在15N2:CO2(图6A,B)的混合气体下均能以N2为唯一氮源固定15N2。作为对比,缺乏固氮基因簇的嗜氢硫杆菌(Sulfurimonas hydrogeniphila) NW10T和铟硫杆菌(Sulfurovum indium) ST-419T不能固氮。在能源中,当菌株利用氧气、硫代硫酸盐(仅菌株HSL1-3)或单质硫作为唯一的电子受体(图6A,B)时,氢和硫化合物如硫代硫酸盐和单质硫可以被利用,其中氢可以作为首选能源。固氮作用与氢的形成有关。因此,通过氢氧化回收能量可以最大限度地减少固氮过程中的能量损失。将固氮和氢氧化相结合以节省能源已被描述为Cyanobacteriota,但也被提及到Pseudomonadota门的其他成员,如Thermochromatium和Rhodospirillum。此外,在低浓度和高浓度(1 mM和40 mM NH4Cl)条件下,氨气的加入完全抑制了15N2的固定。添加1 mM NaNO3和NaNO2对7株菌株均有一定的抑制作用(图S22)。相反,有机氮化合物如尿素和氨基酸不能抑制它们的固氮活性(图S22),这归因于这些菌株不能在这些化合物上作为氮源生长。此外,这7株菌在低氧和高氧浓度(4%和20% O2)下均能固定15N2,这可能是由于它们的抗氧化系统。
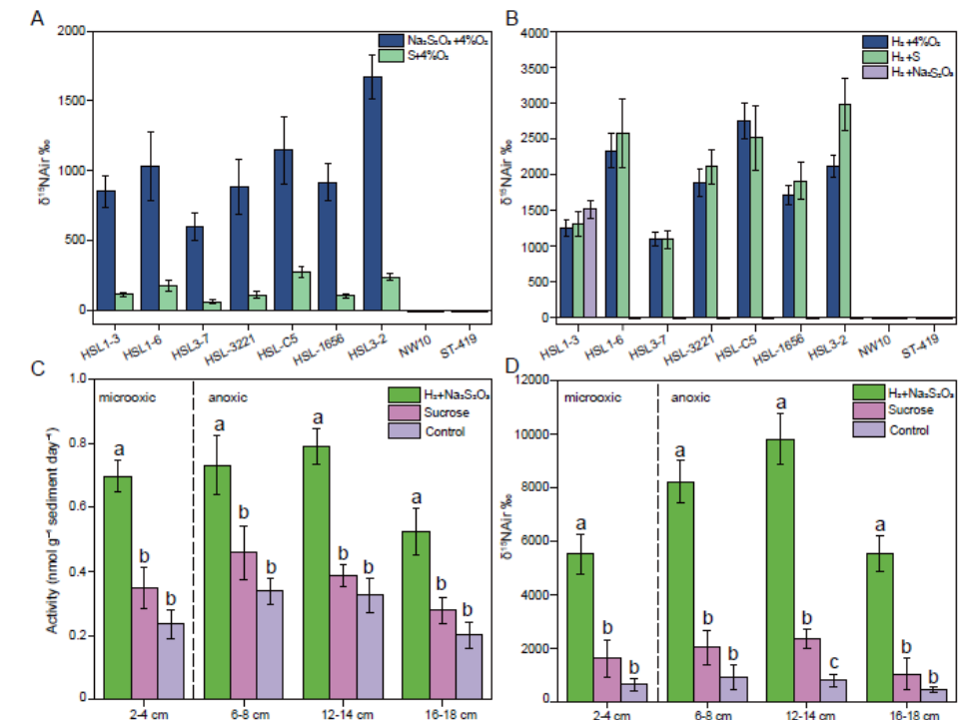
图6 弯曲菌代表菌株和红树林沉积物中的固氮活性。A,15N丰度基于15N2参入测定7株弯曲菌与不同还原性硫化合物孵育作为的能量来源。B,15N丰度基于7株弯曲菌的15N2掺入实验,以H2为唯一能量源与不同的电子受体进行耦合。以缺失nif基因簇的相关型菌株作为对照。C,乙炔还原法测定在化能自养和异养条件下培养的四个沉积物层中的固氮酶活性。D,15N丰度基于15N2掺入实验,在化能自养和异养条件下孵育四种沉积物。标准偏差用误差棒表示。通过t检验(P<0.05),列上的字母表示差异具有统计学意义。
为了验证化能自养菌对红树林沉积物固氮作用的贡献,本研究采用乙炔还原法和15N2掺入法(图6C、D)进行了活性测定。基于测定的氧化还原电位和本研究中检测到的最高转录本表达量,选择4个沉积物层(0~2、6~8、12~14、16~18 cm)来检测固氮活性。结果表明,在微氧和缺氧条件下,红树林沉积物样品的自养固氮活性范围为0.52±0.07~0.79±0.06 nmol g-1day-1,显著高于异养固氮活性(0.28±0.04至0.46±0.08 nmol g-1day-1) (图6C)。在不添加任何碳源或能源的情况下,原始沉积物的固氮酶活性仅为0.19±0.04~0.34±0.04 nmol g-1day-1(图6C)。此外,在培养10天后,添加无机碳源和能源的培养物的δ15N值显著高于添加有机物的处理或原始沉积物的δ15N值(图6D)。
7 红树林沉积物中化能自养型固氮生物型的意义
总之,本研究结果表明,在富含有机质和氮限制的红树林沉积物生境中,化能自养型固氮生物型而不是异养型固氮生物型占主导地位,这可能归因于这些环境中低的氧化还原电位和丰富的还原性无机化合物,如H2和H2S。总体而言,在海洋沉积物中,当Eh<0 mV时,沉积物为强还原性,当Eh值在0~200 mV范围内时,沉积物为弱还原性。在本研究中,4 cm以下的沉积物表现出Eh<0 mV(图S3,表J)的高度还原条件。除表层沉积物外,其余深层沉积物的氧化还原电位均为负值,表明红树林沉积物大部分处于水淹状态,没有周期性的曝气,这与微生物学数据一致。此外,好氧微生物在分解有机质的过程中消耗氧气,这也导致了随着深度的增加氧气耗竭。然而,考虑到红树林根系延伸的干扰以及多毛类和蟹类的掘穴活动,缺氧沉积物中的氧化还原梯度可能是动态的。除了较低的氧化还原电位外,该环境中丰富的还原性无机化合物如H2和H2S也是化能自养菌固氮的重要因素,它们分别来源于有机质的厌氧发酵和沉积物中硫酸盐的还原。
潜在的化能自养固氮菌可能填补了一个重要的生态生态位,有助于红树林沉积物中有机碳和氮的初始积累,并促进生态系统生产力,这与它们在尾矿或冰川前沿等贫营养生境中的作用相似。构建了不同能量源的深度和氧化还原相关的微生物固氮概念模型(图7)。在0~18 cm上层沉积物(Eh=~-117 mV)中,化能自养菌是最主要和最活跃的固氮菌,利用氢气、还原性硫物种和铁作为电子供体,氧气、硝酸盐和硫作为终端电子受体。此外,这些化能自养菌利用二氧化碳形成新的有机碳,为微生物群落中的异养菌提供食物。在18~20 cm (Eh=-120 mV)附近的深层沉积物中,参与异化硫酸盐还原和产甲烷过程的固氮菌,富集程度较强。硫酸盐还原菌通过有机质矿化或H2氧化产生大量硫化物,然后向上扩散,进一步被上层沉积物中的化能自养硫氧化菌利用,形成硫代硫酸盐甚至硫酸盐。小分子化合物如甲酸盐、乙酸盐、丙酸盐、丁酸盐、H2、CO2等来源于红树林深层大分子的厌氧发酵。
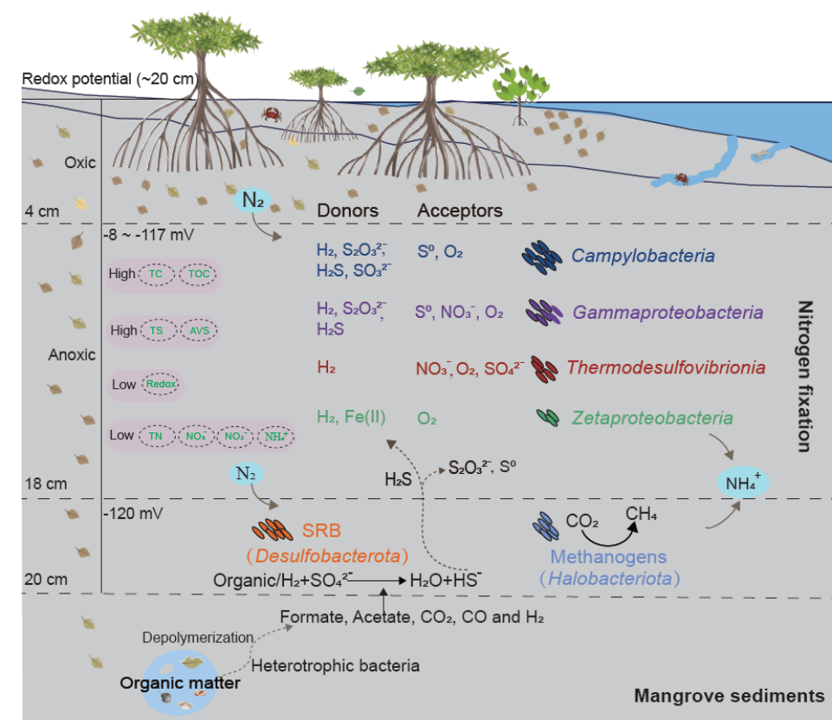
图7 红树林沉积物中与深度相关的微生物固氮概念模型。上层沉积物(0~18 cm):该模型突出了化能无机自养的意义,化能无机自养是先前被忽视的驱动暗固氮的能量来源。这些化能自养型固氮菌以氢、还原硫和铁等无机化合物为能源,以氧、硝酸盐和硫为终端电子受体。H2和H2S分别由降解有机物和硫酸盐还原产生。更深层的沉积物(18~20 cm):参与异化硫酸盐还原和产甲烷的固氮菌强烈富集。硫酸盐还原是利用从深层沉积物中产生的小分子化合物进行的,产生了丰富的硫化物。
原文链接:https://doi.org/10.1093/ismejo/wrae119
宏基因组推荐
本公众号现全面开放投稿,希望文章作者讲出自己的科研故事,分享论文的精华与亮点。投稿请联系小编(微信号:yongxinliu 或 meta-genomics)
猜你喜欢
iMeta高引文章 fastp 复杂热图 ggtree 绘图imageGP 网络iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读

















 1527
1527

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









