







 本文详细探讨了细胞内蛋白质的分选,包括信号假说、分选信号序列以及蛋白质向线粒体和叶绿体的分选路径。同时,介绍了细胞内膜泡运输的机制,如COP II、COP I包被膜泡的装配及运输,以及网格蛋白/接头蛋白包被膜泡的作用,揭示了蛋白质在细胞内的精确定位与转运过程。
本文详细探讨了细胞内蛋白质的分选,包括信号假说、分选信号序列以及蛋白质向线粒体和叶绿体的分选路径。同时,介绍了细胞内膜泡运输的机制,如COP II、COP I包被膜泡的装配及运输,以及网格蛋白/接头蛋白包被膜泡的作用,揭示了蛋白质在细胞内的精确定位与转运过程。
目录
第一节 细胞内蛋白质的分选
蛋白质的靶向转运(protein targeting)或蛋白质分选(protein sorting).
真核细胞中,除少量蛋白质在线粒体和叶绿体内合成外,绝大多数蛋白质都是有核基因编码,或在游离的核糖体上合成,或在糙面内质网结合的核糖体上合成,然后转运到细胞特定部位,组装成结构体与功能复合体,参与细胞生命活动的过程。
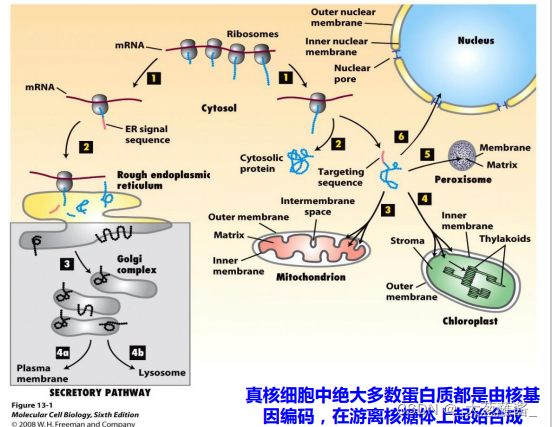
一、信号假说与蛋白质分选信号
信号假说
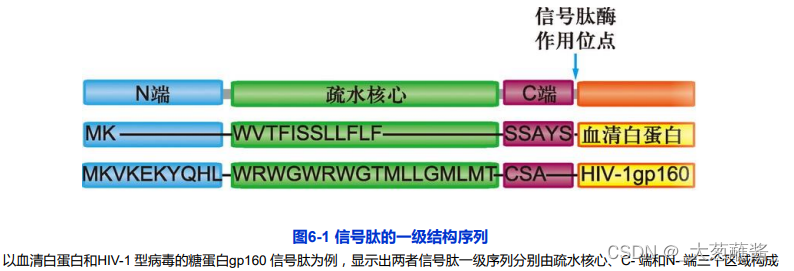
(1)信号肽(signal prptide)
位于蛋白质的N端,一般由16~26个残基组成;似乎没有严格的专一性。
包括疏水核心区、信号肽的C端和N-端三部分。
原核细胞某些分泌蛋白N端也具有信号序列。
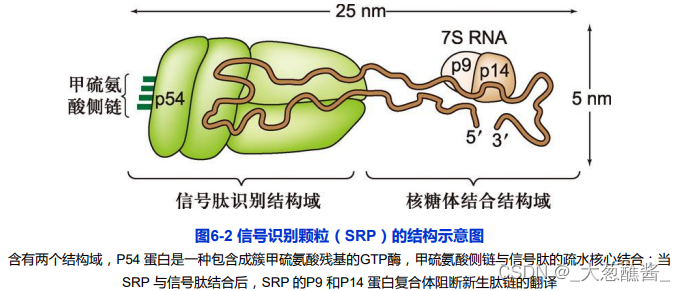
(2)信号识别颗粒(SPR)
由6种不同的蛋白质和一个由300个核苷酸组成的7SRNA组成的一种核糖核蛋白复合体。
通常存在于细胞质基质,既可以于新生肽信号序列和核糖体大亚基结合。又可于内质网膜上SPR受体结合。
(3)SRP 受体(Signal Recognition Particle Receptor)
内质网膜上的整合蛋白,由α和β亚基组成,可特异地与SPR结合。
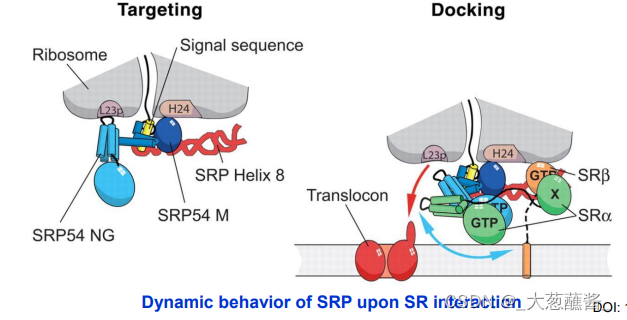
SPR的p54亚基和SPR受体的α亚基与GTP结合时,会增进SPR/新生肽/核糖复合体与SPR受体结合的强度。
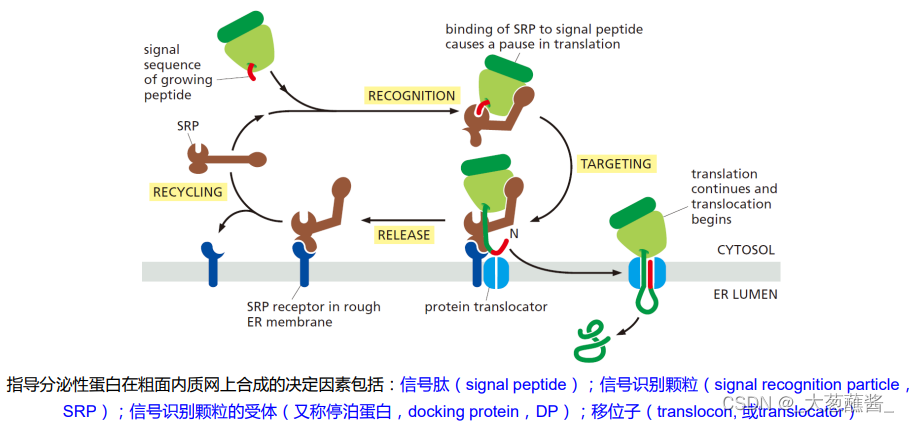
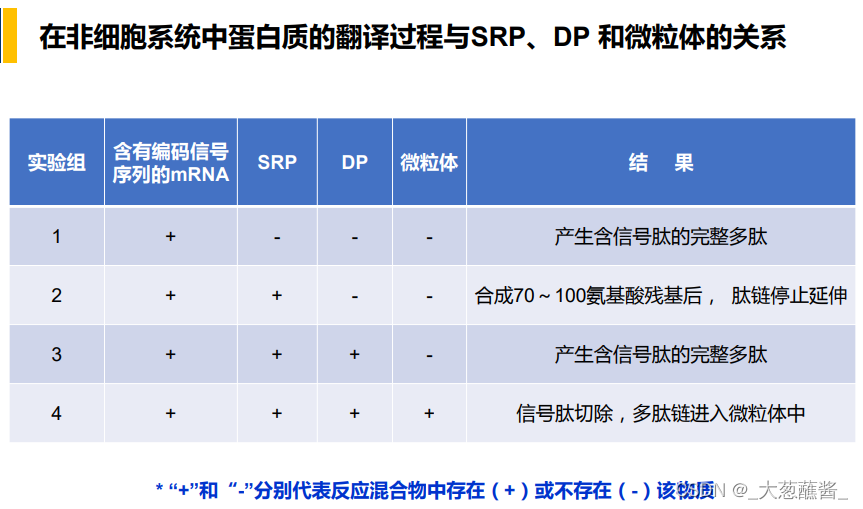
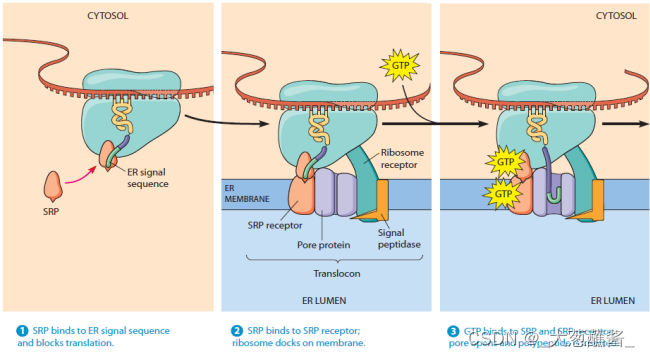
蛋白质翻译过程与SRP、DP和微粒体的关系
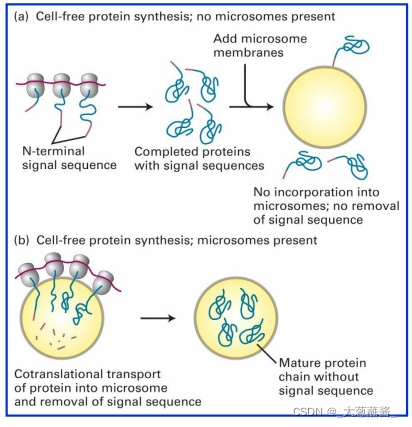
体外非细胞系统(Cell free system)进行蛋白质合成实验,证实分泌性但阿比向rER(microsome,微粒体)腔内的转运是同蛋白质翻译过程耦联进行的,这种分泌蛋白在信号肽的引导下边翻译边跨膜转运的过程称为共翻译转运 (cotranslational translocation)。
(4)分泌性蛋白的合成与共翻译转运过程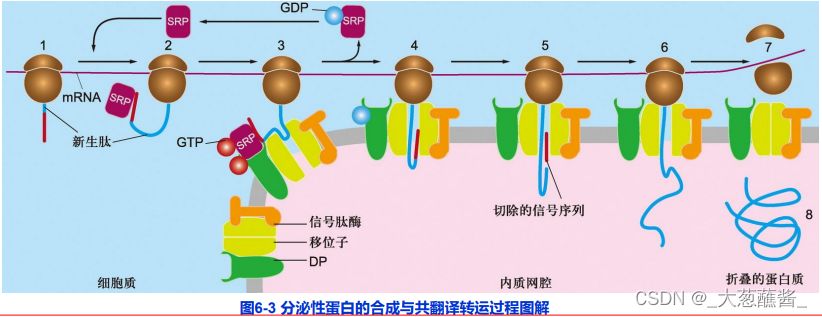
表示信号肽、SRP、SRP受体及移位子之间的相互作用。
- 步骤1、2:分泌蛋白合成起始,内质网信号序列合成并被游离的SRP识别;
- 步骤3:SRP与内质网上的SRP受体结合,引导核糖体——新生肽链复合体附着到内质网膜上,SRP/SRP受体的结合被其结合的GTP所强化;
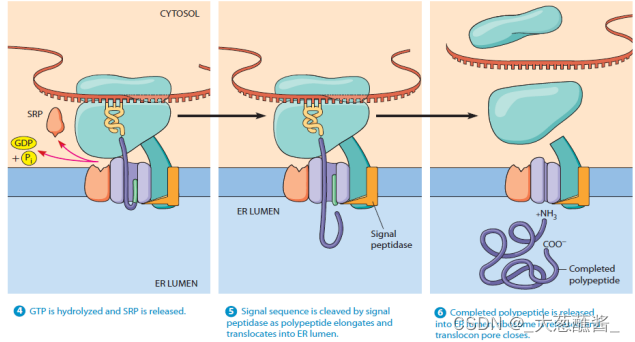
- 步骤4:新生肽——核糖体与内质网膜的位移子(translocon)结合,随着GTP的水解,信号识别颗粒与响应的受体解离,返回细胞质基质中重复使用,肽链又开始延伸,通过位移子以袢环的形式进入内质网网腔中;
- 步骤5:进入内质网腔的新生肽链其信号序列被信号肽酶切割降解;
- 步骤6:新生肽链继续延伸,直至多肽链合成结束;
- 步骤7、8:蛋白质进入腔内并折叠,核糖体释放,移位子关闭。
(5)内质网膜整合蛋白的信号序列
- 起始转移序列(start transfer sequence);
- 内在停止转移锚定序列(internal stop-transfer anchor sequence, STA);
- 内在信号锚定序列(internal signal anchor sequence, SA);
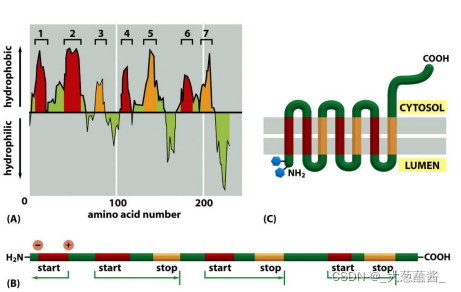
内质网膜整合蛋白的拓扑学类型
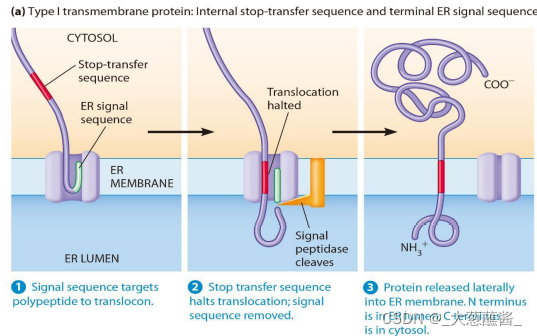
整合膜蛋白的合成——单次跨膜
- 单次跨膜蛋白插入转移机制:新生跨膜蛋白肽链中既有N端信号肽,又有停止转移信号(stop transfer signal)。
整合膜蛋白的合成——多次跨膜
 最低0.47元/天 解锁文章
最低0.47元/天 解锁文章


















 1万+
1万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









