






Environment-sensitivity functions for gross primary productivity in light use efficiency models
摘要
光合作用对环境变化的敏感性对于理解碳循环对全球气候变化的响应以及发展能够解释其时空可变性的建模方法至关重要。我们收集了大量已发表的总初级生产力(GPP)对不同强迫变量的敏感函数,以评估GPP对环境因素的响应。这些包括GPP对温度、蒸汽压缩差异,其中一些包括大气CO2浓度的响应;土壤水分可利用性(W);光照强度和多云程度等。这些函数被组合在一个 全因数 光能利用率(LUE)模型结构中,形成了5600个不同LUE模型的集合。每个模型都根据来自196个FLUXNET站点每日GPP和蒸散通量进行优化,并基于自举方法在站点之间进行排名。显示出每个环境因子(包括CO2施肥)对GPP敏感性显著,并且先前发表的任何模型结构都无法与最佳选择相媲美。从日尺度到周尺度再到月尺度,最佳模型在各站点上介于0.73、0.79和0.82之间,在年尺度上较差(0.23),强调了当前模型在描述GPP年际变异性方面的普遍局限性。尽管全球最佳模型与每个站点的最佳模型不完全匹配,但选择结果在生态系统类型之间是稳健的。光饱和度和多云程度对GPP在所有生物群落中均有贡献(从23%到43%)。温度和W主导着GPP和LUE,但GPP对温度和W的响应在寒冷和干旱生态系统中存在滞后。本研究的发现为更可靠地基于LUE估计全球GPP提供了基础,并可能为其他经验性GPP产品提供参考标准。
Introduction
总光合速率由总初级生产力 (GPP) 估算,是陆地生态系统碳吸收的衡量标准,由能源、水、大气二氧化碳和养分的可用性驱动(Beer 等,2010;Haverd 等).,2018;Ichii 等,2005;Zhang 等,2011)。由于对这些因素的互补和同时依赖,GPP 的响应和敏感性的量化通常具有挑战性(Anav 等人,2015;Ciais 等人,2014;Keenan 等人,2012;Zaele 等人,2014)。
目前,有多种为全球目的而开发的 GPP 模型。一些模型是根据局部测量开发的,用于机械地代表植物功能和生理学(Kaminski et al., 2013; Law et al., 2002; Lawrence et al., 2011; Vuichard et al., 2019),而其他模型则更多本质上是经验性的(Jung 等人,2009;Teubner 等人,2019)。机械模型通常在各种气候和环境条件下都很稳健(De Pury 和 Farquhar,1997)。然而,由于模型复杂性、参数鲜为人知以及由于不同的嵌入假设而导致它们之间的差异,它们在更大空间尺度上的应用通常受到限制(Anav et al., 2015)。相比之下,经验模型需要较少的驱动因素和参数,但受到数据质量、代表性和通用性的限制(Jung et al., 2020)。一般来说,它们缺乏与生产力根本原因的理论联系(Jung et al., 2011)。相反,光利用效率(LUE)模型(Monteith,1972)起源于纯粹的经验模型,并结合一些理论约束而发展起来,融合了两者的优点(Carvalhais 等,2010a;Carvalhais 等,2010a)。2008 年;Mäkelä 等人,2008 年;Running 等人,2004 年)。
这里处理的 LUE 模型将 GPP 定义为吸收的光合有效辐射 (APAR) 和 LUE (ε) 的乘积。 APAR 是光合有效辐射 (PAR) 和吸收 PAR 分数 (FAPAR) 的乘积。 ε 表示根据环境因素 (fX) 的灵敏度函数缩放的最大 LUE (εmax)。 LUE模型共同假设ε代表冠层尺度上特定光照下的所有光合作用过程,fX表示它们对环境因素的反应和敏感性。这一假设后来得到了 Wang 等人的验证。 (2017a),他从详细的叶尺度光合作用过程中推导出 P 模型的 ε 和 fX。
作为一个或多个环境因素的函数,fX 因模型而异。温度 (T) 是每个 LUE 模型中的驱动因素,因为它对酶活性和电子传输具有重要影响(Medlyn 等,2002)。 T 的部分敏感性函数 (fT) 可以表示为线性峰值函数或 S 形函数,表明光合速率不断增加,直至达到稳定水平(Mäkelä 等人,2008 年;Running 等人,2004 年)。或者,fT 可以是一个钟形函数,以在 T 超过某个最佳值时解释减少(Horn 和 Schulz,2011a;Potter 等人,1993;Xiao 等人,2004)。它还可以包括 T 的滞后函数(Horn 和 Schulz,2011a;Mäkelä 等,2008),以表示温度对北方和温带植被的延迟影响(Jarvis 等,2004;Mäkelä 等,2004)。
除了 T 之外,一些模型还包括蒸气压亏缺 (VPD)(Horn 和 Schulz,2011a;McCallum 等,2013;Running 等,2004;Wang 等,2017a;Wang 等,2015),土壤供水量(W;Stocker等,2018)、干旱指数(Yan等,2017)、降水指数(Horn和Schulz,2011a)、地表水指数(LSWI;Xiao等,2004)、或蒸发分数(Horn 和 Schulz,2011a;Yuan 等人,2007)作为植物需水量的指标。 VPD 的部分敏感性函数 fVPD 是一个递减函数,代表气孔导度的限制,可以是线性的(Running 等人,2004 年)或 S 形的(Horn 和 Schulz,2011a;Kalliokoski 等人,2018 年;Mäkelä 等人)等人,2008;王等人,2018)。相比之下,W (fW) 的部分灵敏度函数通常线性增加(Kalliokoski 等人,2018;Turner 等人,2006a;Xiao 等人,2004)或非线性增加(Horn 和 Schulz,2011a;Mäkelä 等人) ., 2008; Stocker et al., 2020),因为充足的供水可以保证气孔开放和理化反应。 fVPD 或 fW 都可以包括干旱地区的滞后效应(Horn 和 Schulz,2011a),与根系分布和干旱程度有关。
一些模型将 APAR 的部分灵敏度函数(称为 fL)嵌入到 LUE 模型中(Ibrom 等人,2008 年;Kalliokoski 等人,2018 年;Mäkelä 等人,2008 年;Propastin 等人,2012 年)来解释光饱和效应(Medlyn,1998),与传统的线性 LUE 模型不同。
研究发现,漫射辐射可以增加总冠层ε,因为更多的光可以扩散到更深的冠层(Cheng等人,2015年;Farquhar和Roderick,2003年;Gu等人,1999年;Huang等人,2014年;Knohl和Baldocchi,2008;Mercado 等人,2009)。因此,以云度指数(CI)为代表的漫辐射比的影响已被纳入一些LUE模型中(He et al., 2013; Turner et al., 2006a; Wang et al., 2015; Wang et al., 2015)。2018; Yan et al., 2017),并得到经典辐射传输理论(Ross, 1981)的支持,并被机械冠层模型采用(Ibrom et al., 2006)。 CI 的部分敏感性函数 (fCI) 可以是求和形式(Turner et al., 2006a)或阶乘形式(Wang et al., 2018),这两种形式都显示出对 CI 的单调正响应。或者,双叶 LUE 模型结构将阳光照射和阴影叶子的 εmax 和 FAPAR 分开,可以解释漫辐射比对植物不同部分的影响。在当前的 LUE 模型中,fL 尚未与 fCI 耦合,可能是由于它们之间的相关性(Gu 等人,2002 年;Knohl 和 Baldocchi,2008 年)。
光合作用的另一个驱动因素是大气中的二氧化碳 (Farquhar et al., 1980; Haverd et al., 2018; Kaminski et al., 2013),它可以通过气孔扩散到叶子中 (Leuning, 1995)。然而,大多数 LUE 模型都忽略了它,除了 CFix (Veroustraete et al., 2002)、P (Stocker et al., 2020)、修订后的 EC-LUE (Yuan et al., 2019) 和 PRELES 模型 (Kalliokoski) et al., 2018),这意味着传统的 LUE 模型对 CO2 浓度增加的影响不敏感。由于气孔导度依赖于 VPD 来控制气体交换,因此 CO2 变化的影响通常包含在 fVPD 中(Kalliokoski 等人,2018;Wang 等人,2017a)。 CO2 效应也受 T 影响,T 影响 Rubisco 对 CO2 分子的亲和力(Veroustraete 等,2002;Wang 等,2017a)。
fX 之间的所有上述差异导致了 LUE 模型和 GPP 估计之间的差异。尽管其中一些模型已经在全球范围内进行了比较(Yuan 等人,2014;Zhang 等人,2015),或在区域范围内进行了比较(Carvalhais 等人,2010b;Mäkelä 等人,2006;Ruimy 等人,1999),使用多个模型的贝叶斯平均值改进了 GPP 估计(Zhang 等人,2021),但尚未评估不同环境因素的必要性、其公式以及 GPP 动态的衍生敏感性。
我们的研究目标是评估环境因素和部分敏感性函数,为 FLUXNET 中的大多数涡流协方差 (EC) 站点找到一个总体稳健的 LUE 模型(Baldocchi 等,2001)。我们假设全球生态系统层面的 GPP 和 LUE 变化将对 T、VPD、W、光饱和度、CI 和 CO2 的独立变化做出响应,并且响应(即部分灵敏度函数)是非线性的。为了检验这些假设,我们的研究遵循图 1 中的工作流程。
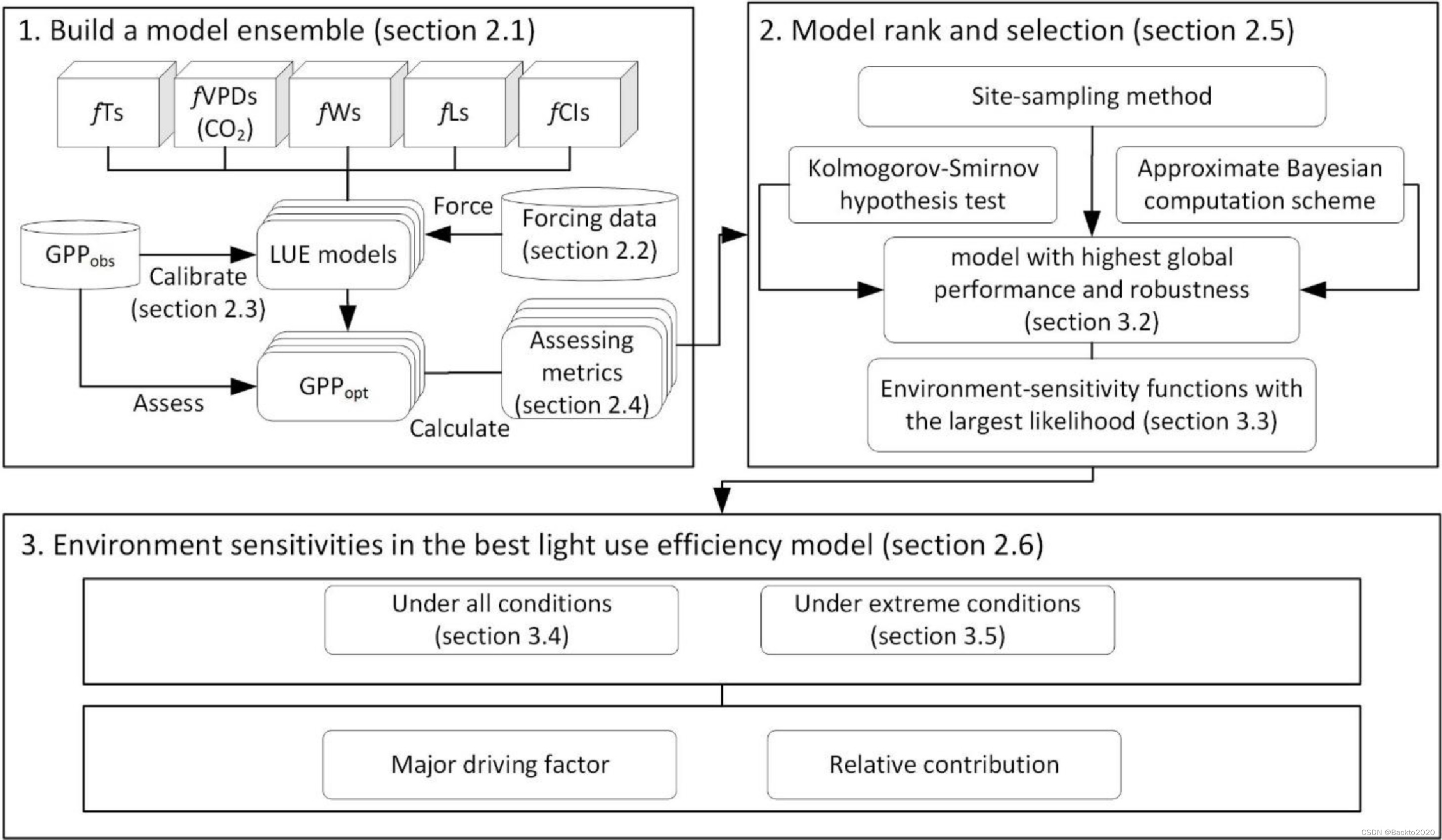 ## 材料和方法
## 材料和方法
Light use efficiency model ensemble
第一步,我们基于全阶乘结构(方程(1))构建了 5600 个 LUE 模型的集合,这使得可以最有效地测试单个环境因素的影响。
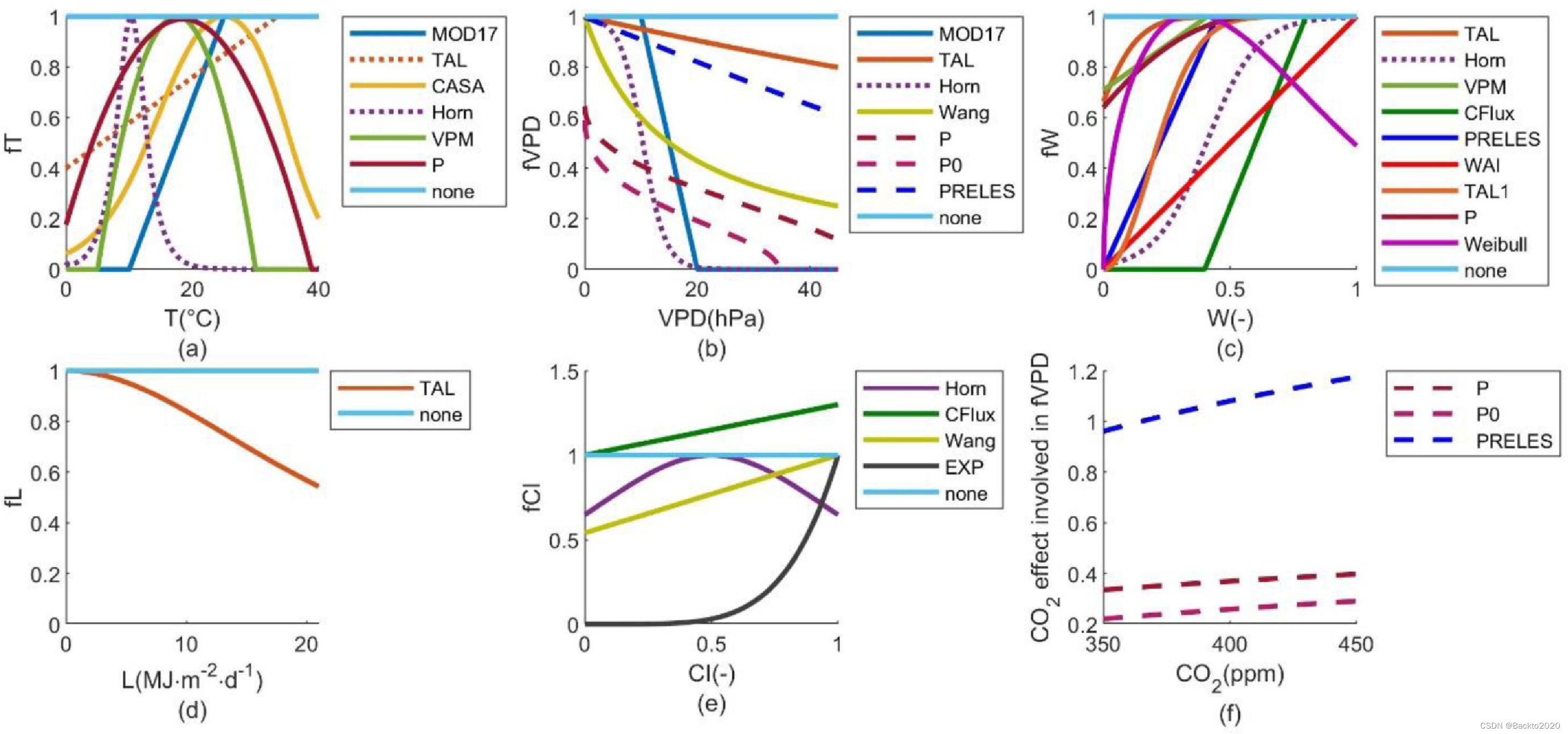
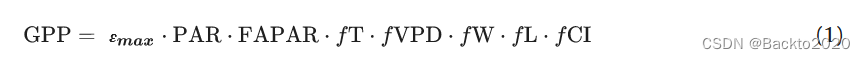 我们选择 T、VPD、W、光饱和度、CI 和 CO2 作为控制 GPP 的最主要因素。鉴于 CO2 扩散与 VPD 控制的气孔导度之间的关系,VPD 和 CO2 效应被整合为单一函数 fVPD。我们从已发布的 LUE 模型中收集了五种独立的部分敏感性函数(六种 fT、六种 fVPD,其中两种包括 CO2 效应、八种 fW、一种 fL 和两种 fCI,如表 1 所示)的不同数学表示(参见表 1)。参考表1)。 fVPDP0 是具有 CO2 效应的 fVPD 之一,它具有适用于所有气候和植被类型的固定参数(Stocker 等人,2020)。因此,我们用灵活的参数(fVPDP)补充了相同的功能。此外,我们还引入了土壤湿度的威布尔函数 (fWWeibull),以及 CI 的指数函数和钟形函数(fCIEXP 和 fCIHorn)。我们还为每个因素(fTnone、fVPDnone、fWnone、fLnone 和 fCInone,均等于 1)设置一个对照组,以测试 GPP 是否独立于这些环境因素。总之,我们根据式(1)中的结构组合了7个fT、8个fVPD、10个fW、2个fL和5个fCI(见图2中的函数曲线,表1中的方程和参考文献)。 (2) 制作 5600(=7 × 8 × 10 × 2 × 5) 个 LUE 模型的集合。
我们选择 T、VPD、W、光饱和度、CI 和 CO2 作为控制 GPP 的最主要因素。鉴于 CO2 扩散与 VPD 控制的气孔导度之间的关系,VPD 和 CO2 效应被整合为单一函数 fVPD。我们从已发布的 LUE 模型中收集了五种独立的部分敏感性函数(六种 fT、六种 fVPD,其中两种包括 CO2 效应、八种 fW、一种 fL 和两种 fCI,如表 1 所示)的不同数学表示(参见表 1)。参考表1)。 fVPDP0 是具有 CO2 效应的 fVPD 之一,它具有适用于所有气候和植被类型的固定参数(Stocker 等人,2020)。因此,我们用灵活的参数(fVPDP)补充了相同的功能。此外,我们还引入了土壤湿度的威布尔函数 (fWWeibull),以及 CI 的指数函数和钟形函数(fCIEXP 和 fCIHorn)。我们还为每个因素(fTnone、fVPDnone、fWnone、fLnone 和 fCInone,均等于 1)设置一个对照组,以测试 GPP 是否独立于这些环境因素。总之,我们根据式(1)中的结构组合了7个fT、8个fVPD、10个fW、2个fL和5个fCI(见图2中的函数曲线,表1中的方程和参考文献)。 (2) 制作 5600(=7 × 8 × 10 × 2 × 5) 个 LUE 模型的集合。

 在该集成中,有 11 个模型与已发布的模型具有相同的结构(参见补充信息表 S1 中的名称和结构)。
在该集成中,有 11 个模型与已发布的模型具有相同的结构(参见补充信息表 S1 中的名称和结构)。
强制数据和观察限制
我们强制 LUE 模型使用来自 196 个 FLUXNET EC 站点的生态系统规模观测结果 (fluxnet.fluxdata.org
;分布图见图S1,站点信息见表S2)。表 2 总结了强迫数据,包括 PAR、FAPAR、T、VPD、W、CI、CO2 和场地高程。我们还收集了 GPP (GPPobs)、蒸散量 (ET;ETobs) 及其不确定性(参见定义)表2)校准模型参数。
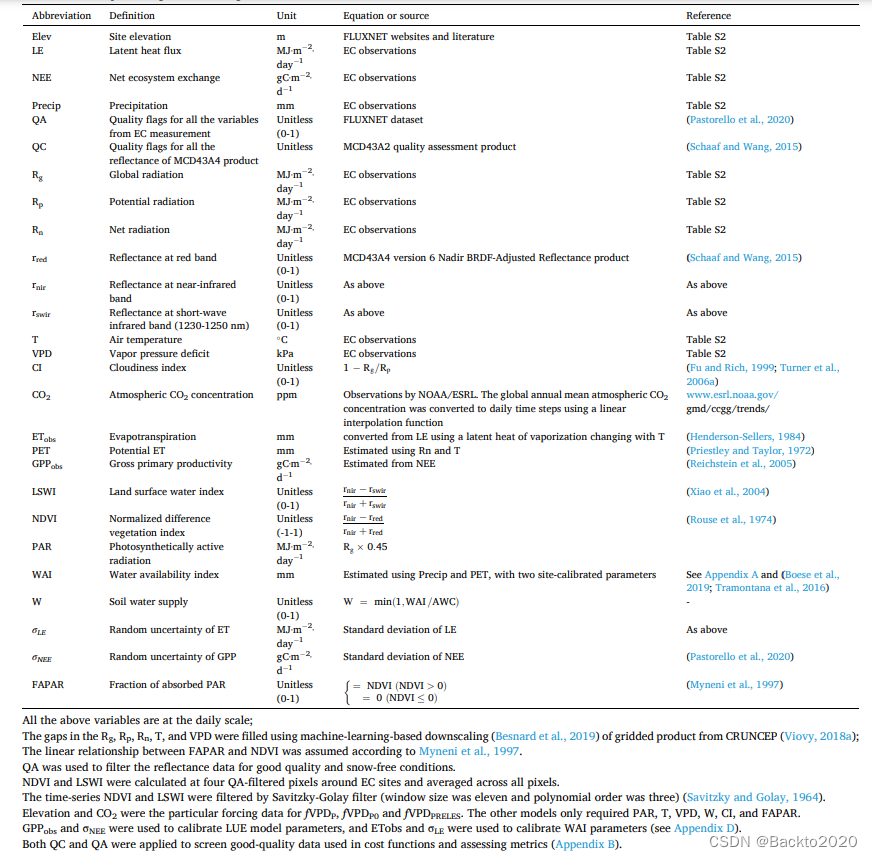
模型标定
模型标定的目的是为每个站点的每个LUE模型找到最佳参数向量,并减少与模型参数相关的模型不确定性。我们使用随机且无导数的模型校准了LUE模型参数、εmax和fX参数(表1中以粗体显示)和WAI参数(参见附录A中的定义以及附录C中联合校准WAI和LUE参数的原因)进化算法,CMAES(Hansen 和 Kern,2004)。 CMAES 是一种可靠的全局优化工具(Hansen 和 Kern,2004 年;Trautmann 等人,2018 年),它通过最小化成本在预定义范围(表 S4)内搜索最佳参数(本研究中最大迭代次数 = 105)功能。因此,在给定模型变量的某些约束的情况下,适当成本函数的定义对于找到最佳参数至关重要。我们将成本函数 (cf) 定义为四个部分的总和(请参阅附录 D 中的成本函数方程):按约束 LUE 参数的不确定性缩放的 GPP 误差 (cf1),按约束 WAI 参数的不确定性缩放的 ET 误差 (cf2)和 fX 约束(cf3 和 cf4)。
为了避免选择局部最优,我们在进化参数搜索过程的最后103次迭代中,使用cf的倒数作为权重,随机选择9个附加参数向量(即103个参数向量)来交叉检验最优参数向量由 CMAES 给出。十个参数向量均用于模型选择(第 2.5 节)。参数不确定性也使用参数协方差矩阵进行量化(附录 E)。
模型评估指标
我们使用附录 B 中定义的高质量观测数据在每日、每周、每月和每年的尺度上评估模型的适应性。首先,我们汇总了每日观测的 GPP (GPP) 和模拟的 GPP (
)到每周、每月和每年的规模。接下来,我们使用等式计算 Nash-Sutcliffe 效率(NSE)来衡量模型的全局适应度(NSEg)、空间适应度(NSEsp)和站点水平适应度(NSEi)。 (2)-(4)。
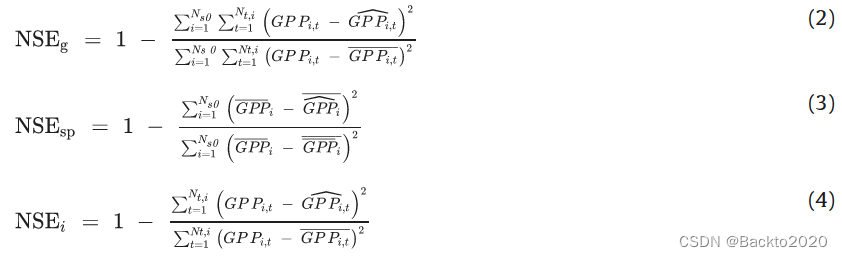 GPPi,t 代表在时间步 t 在地点 i 观测到的 GPP 和模拟的 GPP。 Ns0 和 Nt,i 是站点 i 的总站点数和总时间步长。和 是站点 i 的平均观测 GPP 和模拟 GPP。每个站点年和跨站点年(第 2.5 节中使用)的 NSE 与等式 1 相同。 (4)和(2),但只考虑了一年的GPP。同样,我们计算了决定系数 (R2) 和归一化均方根误差 (nRMSE)。
GPPi,t 代表在时间步 t 在地点 i 观测到的 GPP 和模拟的 GPP。 Ns0 和 Nt,i 是站点 i 的总站点数和总时间步长。和 是站点 i 的平均观测 GPP 和模拟 GPP。每个站点年和跨站点年(第 2.5 节中使用)的 NSE 与等式 1 相同。 (4)和(2),但只考虑了一年的GPP。同样,我们计算了决定系数 (R2) 和归一化均方根误差 (nRMSE)。
模型排名和选择
模型的选择通常取决于方法、数据信息和评估指标(Burnham 和 Anderson,2002;Burnham 等,2011;Efron,2014)。模型之间和模型参数之间的等价性限制了在观测有限的情况下复杂生态系统的模型选择(Beven,1993;Fisher 和 Koven,2020)。因此,在给定几个观测点的大型模型集合中选择鲁棒模型具有挑战性,这些观测点比模型总数少了近 28 倍。在这里,我们采取了以下策略来减轻模型选择方法、地点、评估指标和模型等效性的影响。
首先,为了减少使用独特的排名方法带来的不确定性,我们基于三种方法对模型进行排名:(1)站点抽样方法; (2) 成对 Kolmogorov-Smirnov (KS) 假设检验(附录 F;Lilliefors,1967); (3) 遵循近似贝叶斯计算 (ABC) 方案(附录 G;Toni 和 Stumpf,2010)。在这三种方法中,站点采样和ABC都考虑了模型和参数向量之间的等价性,使用似然值来衡量模型的鲁棒性。
为了避免选择局部适合特定站点组的模型,我们使用了站点采样方法,该方法使用在引导站点的多次迭代中成为最佳模型的可能性来定义模型的稳健性。表3总结了现场采样方法的详细工作流程。
 模型似然 P1 代表每个站点年的模型优度,可以视为模型跻身前 1% 最佳模型之列的站点年的分数。反过来,另一个模型可能性 P2 代表跨站点年份的模型优度,即所有站点年份的总体优度。通过根据 P1 和 P2 排序的模型排名 R1 和 R2,可以选择对于大多数站点年份而言局部最佳和总体最佳的模型。
模型似然 P1 代表每个站点年的模型优度,可以视为模型跻身前 1% 最佳模型之列的站点年的分数。反过来,另一个模型可能性 P2 代表跨站点年份的模型优度,即所有站点年份的总体优度。通过根据 P1 和 P2 排序的模型排名 R1 和 R2,可以选择对于大多数站点年份而言局部最佳和总体最佳的模型。
为了测试指标的选择是否影响模型选择,我们使用 RMSE 而不是 NSE 重复上述分析(表 3 中步骤 13 中的 mef,对应于各自场景中的 RMSE 或 NSE)。我们还测试了其他阈值(表 3 中的步骤 21)以确定最佳模型(θ0=2%、3%、4%、5% 和 10%)。结果如图S3和图S4所示。
我们对九种气候植被类型分别重复了上述模型选择过程:干旱森林、干旱草原、北方森林、北方草原、极地植被、温带森林、温带草原、热带森林和热带草原(定义见附录 H)。最后,我们选择全局最佳模型作为所有站点和各种气候植被类型站点R1和R2之和最小的模型;即 fX 的最佳组合。
为了评估全球最佳 LUE 模型的强度,我们将其与机器学习模型(附录 I)和站点最佳模型集合进行了比较;即每个站点每天具有最高 NSE 的模型集合。
此外,我们还计算了各种偏敏感函数及其组合的可能性 P1 和 P2。似然等于所有迭代的前 1% 最佳模型中出现的部分敏感性函数(例如 fTMOD17)或部分敏感性函数的组合(例如 fTMOD17 和 fVPDMOD17)与总数的 1% 的平均比率型号数量。我们将部分敏感性函数和具有最大似然之和的组合视为每个环境因素或环境因素组合的最佳敏感性函数。
最佳光利用效率模型中的环境敏感性
我们通过三种方法使用全局最佳 LUE 模型和站点最佳模型的部分敏感性函数分析了季节性 LUE 动态的主导控制。在第一种方法中,我们的目标是找到 LUE 最敏感的环境因素。为此,我们计算了平均偏敏感度,并采用最小值作为地点 i (Xi) 的主导环境因素,如下式: (5)
 然后,我们计算了每种气候植被类型的每个主导因子 (FracX) 的分数,如下式: (6) 式中,Ns0 表示相应气候植被类型的总站点数。我们将比例最大的因素视为相应类型的主要驱动因素。
然后,我们计算了每种气候植被类型的每个主导因子 (FracX) 的分数,如下式: (6) 式中,Ns0 表示相应气候植被类型的总站点数。我们将比例最大的因素视为相应类型的主要驱动因素。
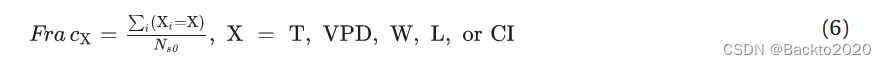 在第二种方法中,我们确定了每个环境因素 (X) 对每种气候植被类型的总 LUE 变化 (CX) 的相对贡献,如下式所示: (7)-(8)。
在第二种方法中,我们确定了每个环境因素 (X) 对每种气候植被类型的总 LUE 变化 (CX) 的相对贡献,如下式所示: (7)-(8)。
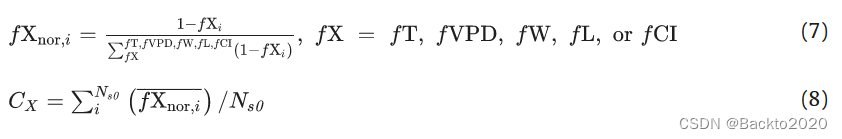 fXnor,i 指位点 i 处的归一化部分灵敏度函数,
fXnor,i 指位点 i 处的归一化部分灵敏度函数,
是位点 i 的平均偏敏感度函数。 CX 表示 X 从小到大(0 到 1)的相对贡献。
在最后一种方法中,我们评估了极端条件下的主导控制,极端条件定义为 T、VPD 或 APAR 大于第 90 个百分位值,或 T、W 或 CI 小于第 10 个百分位值的条件。我们的目的是确定当环境条件受到限制时,哪个因素对 GPP 影响最大。我们计算了极端条件下主要驱动因素的场地比例以及每种气候植被类型的每个环境因素的相对贡献。
为了区分 GPP 和 LUE 的敏感性并交叉验证主要控制,我们将上述变量与 GPPobs 和每个站点的每日强迫变量之间以及 LUEobs、APAR 的 GPPobs 和相同强迫之间的皮尔逊相关系数 ® 进行了比较。
结果
模型整体性能
在全球范围内,大多数校准模型在不同时间尺度上都具有良好的性能(图3)。然而,没有一个模型被选为每个站点或站点年份的最佳模型。站点最佳模型集合总共包括 196 个站点的 166 个不同模型。没有任何一个模型跻身所有站点的最佳 1% 模型之列(由 P1 反映)。同等地考虑每个站点和站点年份(由 P2 反映),无论考虑哪个站点,某些模型都排名前 1%。 P1和P2(图4)的最大值分别为0.82和0.83,这比已发布的模型(P1,P2=0)要好。因此,出现了新颖的 LUE 模型,即 fX 组合,它们对所有站点都同样有效,并且比已发布的模型更好。
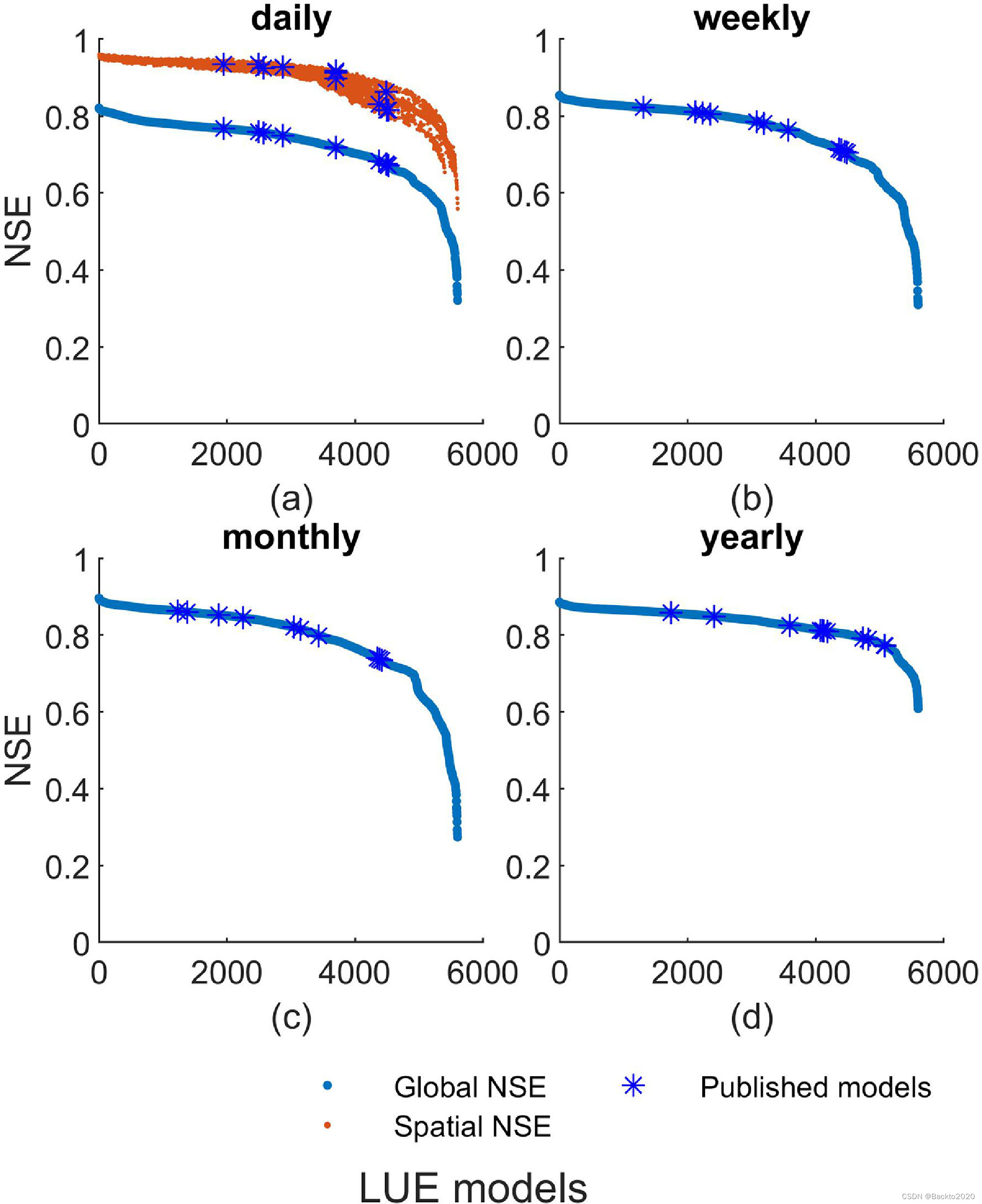 图 3. LUE 模型的 Nash-Sutcliffe 模型效率(NSE)在 a)每日、b)每周、c)每月和 d)每年尺度上的分布。 Global NSE 表示使用所有站点的 GPP 计算的 NSE,与图 6 中的相同。 Spatial NSE 表示使用站点平均 GPP 计算的 NSE。蓝色星号代表本研究中收集的已发表模型
图 3. LUE 模型的 Nash-Sutcliffe 模型效率(NSE)在 a)每日、b)每周、c)每月和 d)每年尺度上的分布。 Global NSE 表示使用所有站点的 GPP 计算的 NSE,与图 6 中的相同。 Spatial NSE 表示使用站点平均 GPP 计算的 NSE。蓝色星号代表本研究中收集的已发表模型
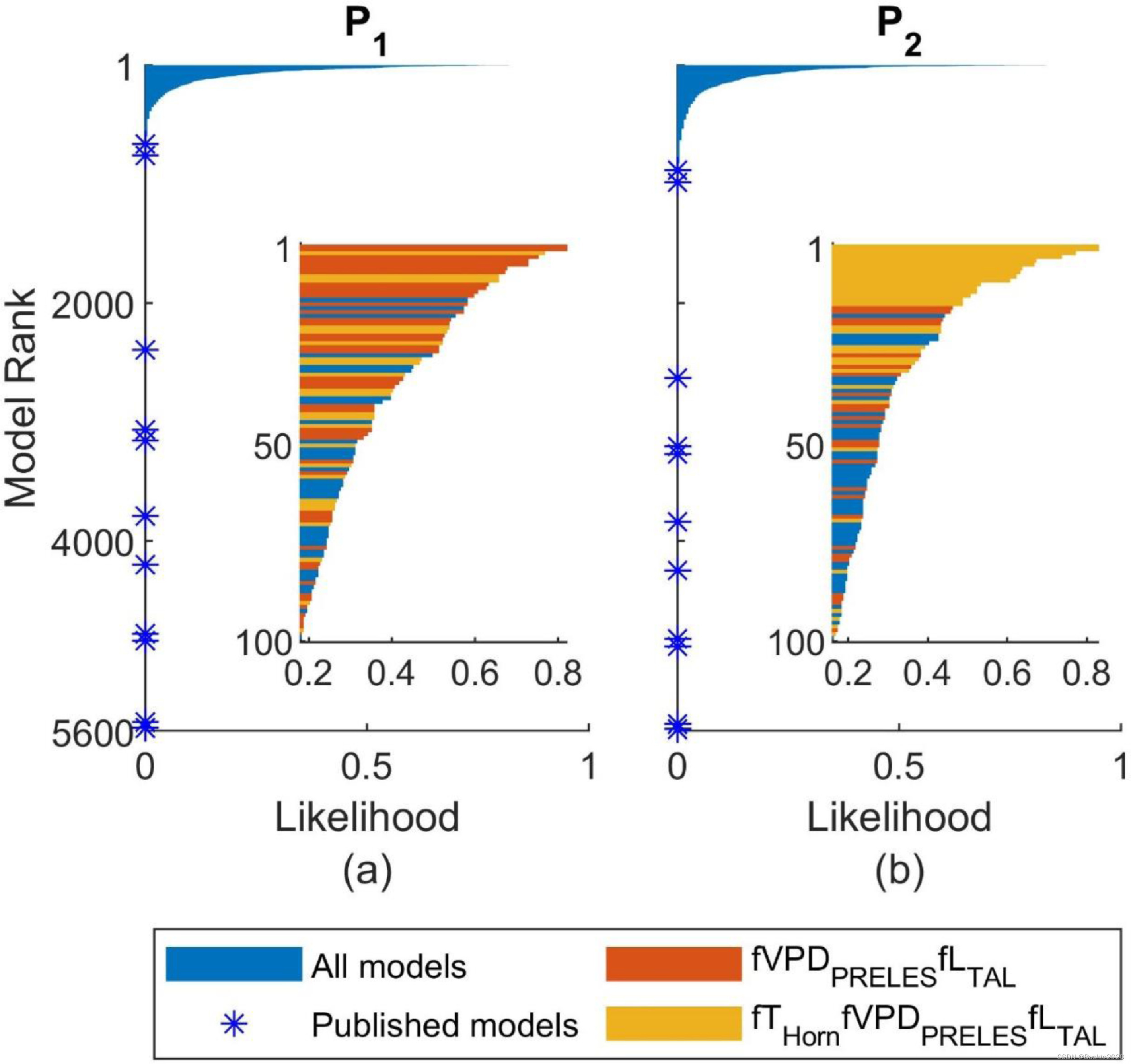 图 4. 基于站点抽样方法的所有 LUE 模型的似然: (a) P1 是每个站点年份中最好的 1% 模型之一的模型的似然,它代表局部优良性,可以视为作为模型跻身最佳 1% 模型之列的站点分数; (b) P2 是模型跻身跨站点年份最佳 1% 模型之列的可能性,代表整体优良性。两个放大窗口是按照P1和P2排序的最好的100个模型。具有 fVPDPRELES 和 fLTAL 结构的模型(红色和黄色)通常比其他模型(蓝色)具有更高的排名。在这些模型中,具有 fTHorn、fVPDPRELES 和 fLTAL(黄色)结构的模型根据 P2 排名较高。已发布模型(蓝色星号)的可能性均为零,这表示这些已发布模型从未跻身于最佳 1% 模型之列。
图 4. 基于站点抽样方法的所有 LUE 模型的似然: (a) P1 是每个站点年份中最好的 1% 模型之一的模型的似然,它代表局部优良性,可以视为作为模型跻身最佳 1% 模型之列的站点分数; (b) P2 是模型跻身跨站点年份最佳 1% 模型之列的可能性,代表整体优良性。两个放大窗口是按照P1和P2排序的最好的100个模型。具有 fVPDPRELES 和 fLTAL 结构的模型(红色和黄色)通常比其他模型(蓝色)具有更高的排名。在这些模型中,具有 fTHorn、fVPDPRELES 和 fLTAL(黄色)结构的模型根据 P2 排名较高。已发布模型(蓝色星号)的可能性均为零,这表示这些已发布模型从未跻身于最佳 1% 模型之列。
具有全球最高性能和鲁棒性的模型
根据站点采样方法,发现具有最高排名(即 R1 和 R2 的最小和)的全局最佳模型(模型#1,具有 fTHorn、fVPDPRELES、fWHorn、fLTAL 和 fCIEXP)为式(1)。 (9)。
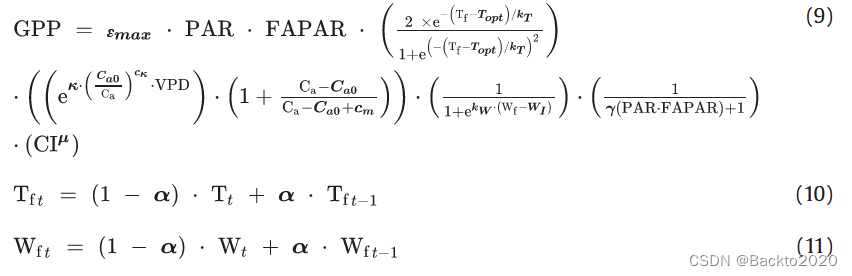 εmax、Topt、kT、κ、cκ、Ca0、cm、kW、WI、γ、μ、α为模型参数,其余为强迫数据。在极地、北方和温带地区,t 天的 T 包括按前一天 t-1 的 α 缩放的滞后效应(方程(10))。干旱地区的 W 也存在同样的滞后效应(方程(11))。在其他区域,α=0,Wf和Tf等于T和W。
εmax、Topt、kT、κ、cκ、Ca0、cm、kW、WI、γ、μ、α为模型参数,其余为强迫数据。在极地、北方和温带地区,t 天的 T 包括按前一天 t-1 的 α 缩放的滞后效应(方程(10))。干旱地区的 W 也存在同样的滞后效应(方程(11))。在其他区域,α=0,Wf和Tf等于T和W。
模型#1 的所有地点和气候植被类型的排名总和最高。即使地点是自举的并且不同的气候植被类型是分开的(图 5),它也是最好的 1% 模型中最常涉及的。干旱草原、干旱森林、极地植被和温带森林的可能性 P1 和 P2 低于其他气候植被类型,而模型 #1 与站点子集的最佳模型没有显着差异。根据显着性水平为 0.05 的 KS 检验,从日到年尺度的站点 NSE 值也没有统计学差异。模型#1也是使用RMSE作为评估指标(参考表3中步骤13的mef)而不是NSE(图S3)并使用其他阈值(θ0=2%、3%、4%, 5%,或表 3、图 S4 中步骤 21 的 10%。根据假设检验(图 S5)和 ABC(图 S6),它分别是第 13 和第二好的模型。基于这两种方法的最佳模型(与模型 #1 具有不同的 fW (fWCFlux) 和 fCI (fCIWang) 以及不同的 fCI (fCIWang))是现场采样方法的第 42 和第三最佳模型。然而,显着性水平为 0.05 的 KS 检验并未显示模型 #1 与这两个模型在位点级别之间存在差异,其中 fW(fWCFlux 和 fWHorn)和 fCI(fCIEXP 和 fCIWang)非常相似(第 3.3 节) )。根据 P2(图 4 中的放大窗口),具有 fTHorn、fVPDPRELES 和 fLTAL 以及各种 fW 和 fCI 的模型的排名高于其他模型。因此,模型#1 是跨气候植被类型的稳健模型。
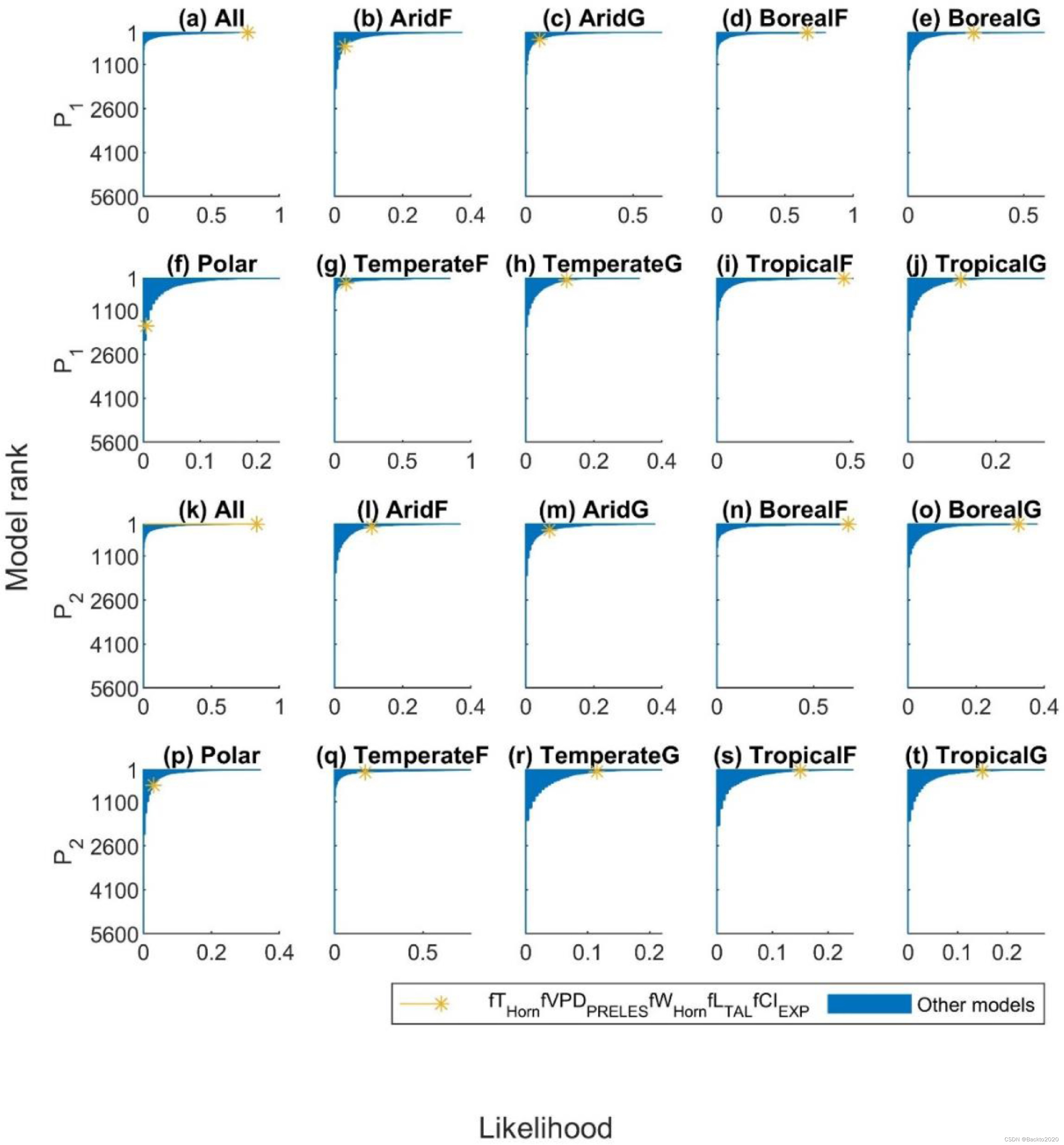 图 5. 根据所有站点(“全部”)和不同气候植被类型(干旱森林 = ‘AridF’、干旱草原 = ‘AridG’、北方森林 =‘)的站点采样方法排序的所有 LUE 模型的排名BorealF’,北方草原=BorealG,极地植被=‘Polar’,温带森林=‘TemperateF’,温带草原=‘TemperateG’,热带森林='TropicalF’和热带草原=‘TropicalG’)。 P1(a-j) 代表每个站点年的局部效益,P2 (k-t) 对应于属于相应气候植被类型的所有站点年的全局效益。带星号的黄线表示所有站点和不同气候植被类型的排名总和最高的模型,这是基于站点采样方法选择的全局最佳模型。其他型号为蓝色。
图 5. 根据所有站点(“全部”)和不同气候植被类型(干旱森林 = ‘AridF’、干旱草原 = ‘AridG’、北方森林 =‘)的站点采样方法排序的所有 LUE 模型的排名BorealF’,北方草原=BorealG,极地植被=‘Polar’,温带森林=‘TemperateF’,温带草原=‘TemperateG’,热带森林='TropicalF’和热带草原=‘TropicalG’)。 P1(a-j) 代表每个站点年的局部效益,P2 (k-t) 对应于属于相应气候植被类型的所有站点年的全局效益。带星号的黄线表示所有站点和不同气候植被类型的排名总和最高的模型,这是基于站点采样方法选择的全局最佳模型。其他型号为蓝色。
与已发布的模型和集成中的所有其他模型相比,全局最佳模型模型#1能够以良好的性能模拟全局GPP(全局NSE=0.82、0.85、0.89和0.88,全局nRMSE=40%,日、周、月、年尺度分别为 34%、29% 和 19%,图 6)。它还可以反映站点平均 GPP 的空间变异性(空间 NSE=0.96,空间 nRMSE=11%)。在站点级别,当使用大于 0.5 的 NSE 作为标准时,模型 #1 可以拟合每日和每周尺度上 85% 站点的时间 GPP 变化,以及每月尺度上 73% 站点的时间 GPP 变化。图 S7a-d 举例说明了日尺度下 NSE 最大、中值、第 25 个百分位和最小站点(= 0.96、0.73、0.58 和 -0.04)的模拟 GPP。模型#1的性能接近站点最佳模型(图7),这表明站点级别的全局最佳模型总体上具有良好的代表性。然而,模型#1 对于年际变化的表现很差。在年尺度上,只有 26% 的站点 NSE 和 41% 的站点 R2 大于 0.5。这在使用站点最佳模型和 MLbest 的 GPP 估计中也很明显(图 S8),这反映了 LUE 和机器学习模型在捕获给定当前强迫数据的 GPP 年际变化方面的常见局限性。虽然 MLbest 通常比 LUE 模型表现更好,但 MLbest 往往会过度拟合 GPP,并在没有高质量强迫数据的情况下导致相当大的误差(图 S9 a 和 b)。反过来,由于功能限制,LUE 模型不存在同样的问题。
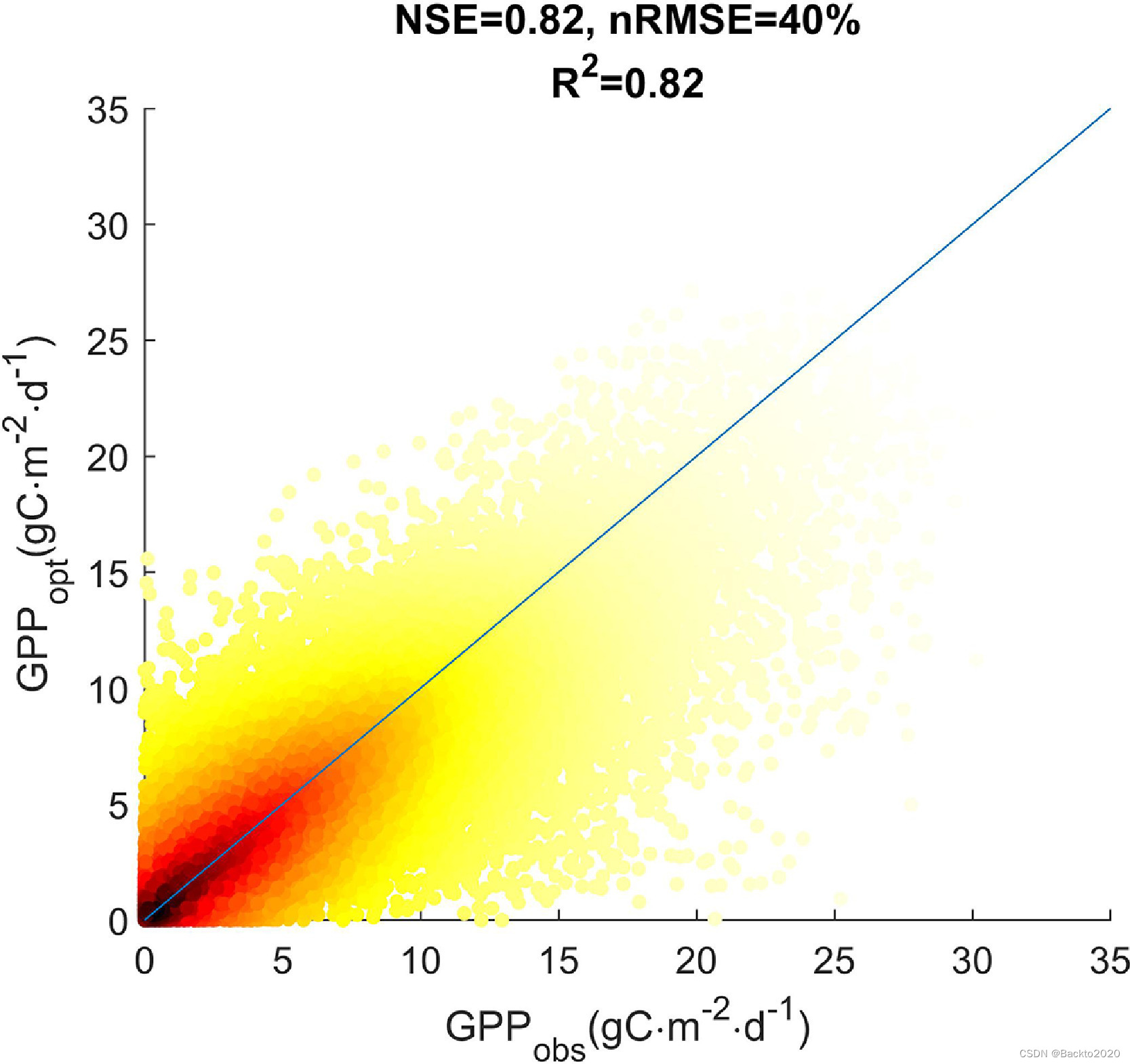 图 6. 使用 EC 数据 (GPPobs) 估计的 GPP 与通过全局最佳 LUE 模型 (GPPopt) 模拟的 GPP 的散布图。散射颜色从黑到黄代表散射密度(从大到小),与GPP的大小有关。 NSE、nRMSE 和 R2 表示 Nash-Sutcliffe 模型效率、通过平均 GPPobs 归一化的均方根误差以及使用由优质数据建模的所有 GPP 计算的决定系数。蓝线是 1:1 线。
图 6. 使用 EC 数据 (GPPobs) 估计的 GPP 与通过全局最佳 LUE 模型 (GPPopt) 模拟的 GPP 的散布图。散射颜色从黑到黄代表散射密度(从大到小),与GPP的大小有关。 NSE、nRMSE 和 R2 表示 Nash-Sutcliffe 模型效率、通过平均 GPPobs 归一化的均方根误差以及使用由优质数据建模的所有 GPP 计算的决定系数。蓝线是 1:1 线。
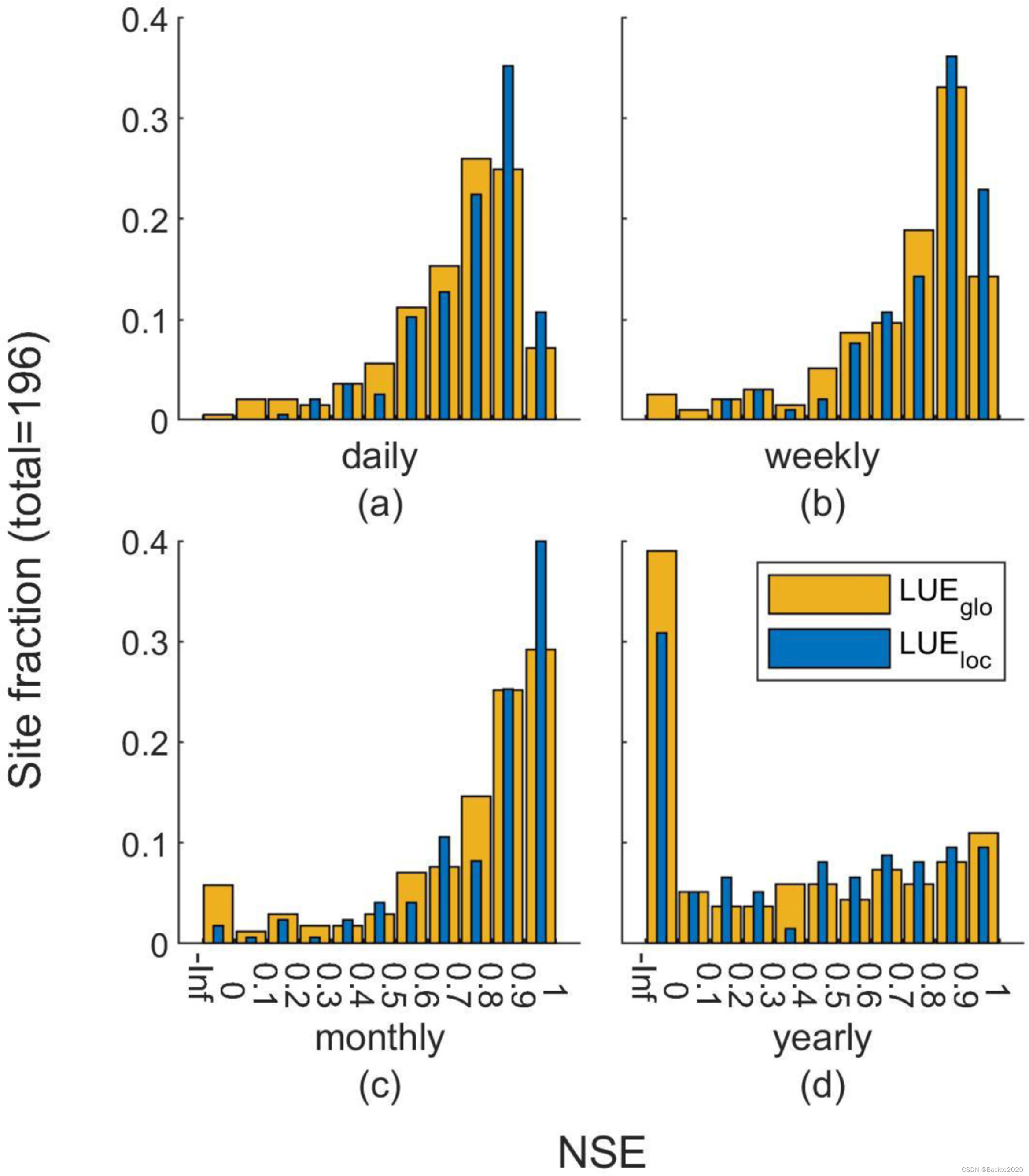 图 7 全球最佳 LUE 模型(LUEglo)和站点最佳 LUE 模型(LUEloc)的站点 NSE 在日(a)、周(b)、月(c)和年(d)尺度上的分布。
图 7 全球最佳 LUE 模型(LUEglo)和站点最佳 LUE 模型(LUEloc)的站点 NSE 在日(a)、周(b)、月(c)和年(d)尺度上的分布。
具有最大似然的环境敏感性函数
所有地点(图8)或各种气候植被类型(图S10)的最佳模型中都没有出现fTnone,并且很少出现fVPDnone、fWnone 和fCInone。尽管一些环境因素和部分敏感性函数是相关的(例如,APAR和CI、fL和fCI,见图S11),但当考虑每个因素的变异性时,模型改进是明显的(图S12)。在一些特定气候植被类型的最佳模型中,对饱和光和二氧化碳施肥(包括在 fVPDPRELES、fVPDP 和 fVPDP0 中)的独立响应可以被忽略,例如,它们在干旱中的可能性(图 S10b 和 g、d 和 i)和极地区域(图S10v和x)并不优越。然而,它们对于北方和温带森林很重要(图S10l和aa、n和ac),其可能性比其他部分敏感函数大得多。因此,考虑光饱和度和 CO2 施肥对于全局 LUE 模型是必要的。
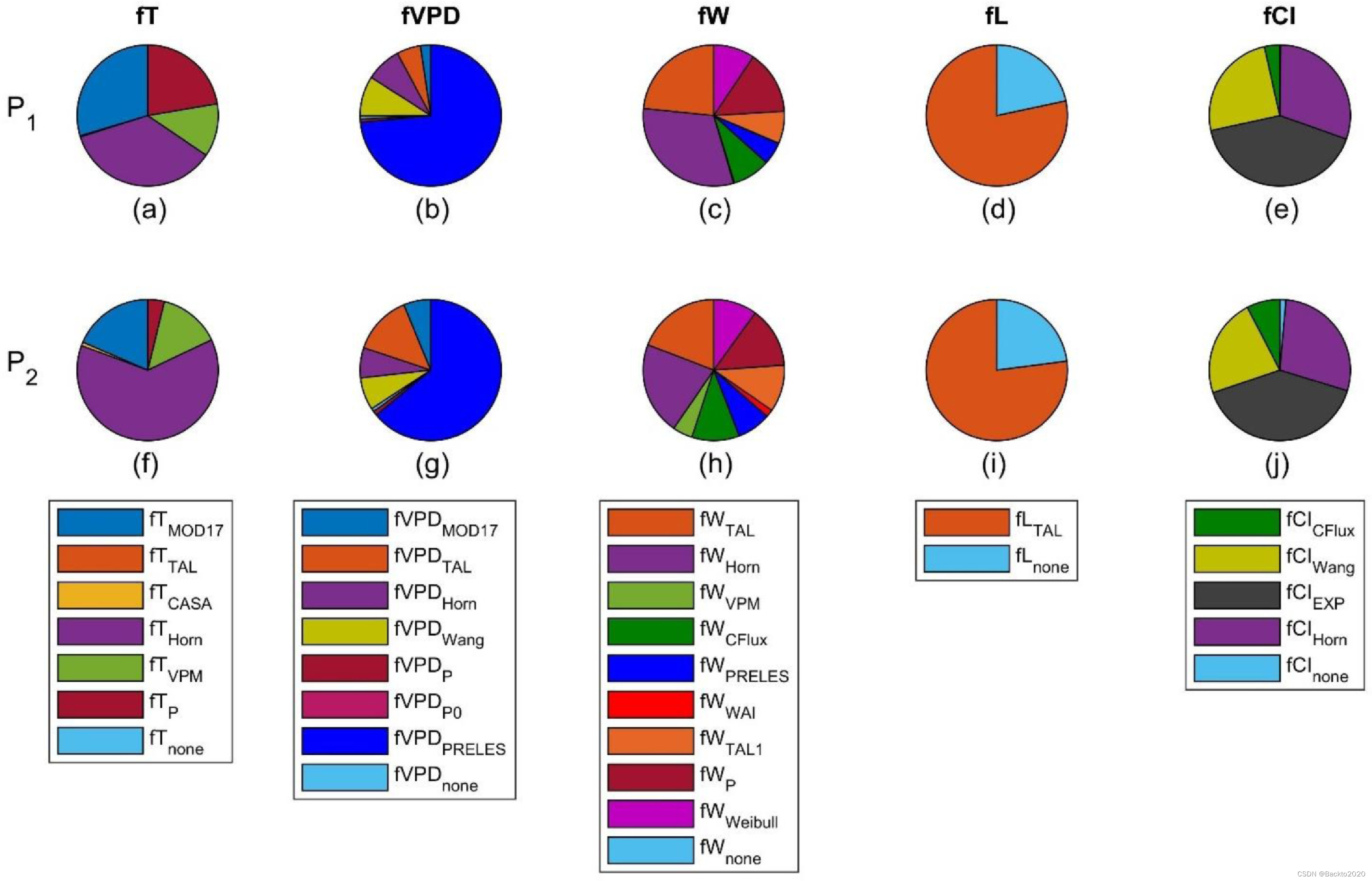 图 8. 部分敏感性函数的似然,等于从 P1 (a-e) 和 P2 (f-j) 导出的最佳 1% 模型中的比率。两个 fT(fTTAL 和 fTHorn)和一个 fW(fWHorn)包括北方和干旱气候的滞后效应,三个 fVPD(fVPDPRELES、fVPDP、fVPDP0)包括 CO2 效应。
图 8. 部分敏感性函数的似然,等于从 P1 (a-e) 和 P2 (f-j) 导出的最佳 1% 模型中的比率。两个 fT(fTTAL 和 fTHorn)和一个 fW(fWHorn)包括北方和干旱气候的滞后效应,三个 fVPD(fVPDPRELES、fVPDP、fVPDP0)包括 CO2 效应。
一些偏敏感函数在表示 LUE 对全球环境因素变化的响应方面明显优于其他函数。 fVPDPRELES(VPD 和 CO2 的联合效应)和 fLTAL(光饱和效应)在更多站点年份(由P1)和跨站点年份(由 P2 表示)。 fTHorn 是一种在寒冷气候(即极地、寒带和温带)中具有滞后效应的钟形函数,优于其他 fT(图 8a 和 f)。至于 fW 和 fCI,几个函数的表现类似(图 8c、e、h 和 j),但 fWWAI 的性能较差,其中响应 W 变化的斜率是固定的。因此,许多具有 fTHorn、fVPDPRELES 和 fLTAL 以及不同 fW 和 fCI 的模型都是最好的模型(图 4,黄色)。根据KS检验和ABC选择的最佳模型,其fT、fVPD和fL相同,但fW和fCI不同,也与模型#1相似。
除了其中两个之外,成对的部分灵敏度函数的组合没有表现出明显的通用特征。其中之一是 fVPDPRELES 和 fLTAL,其可能性比 fVPD 和 fL 的其他组合大得多(图 S13)。另一个是 fTHorn 和 fLTAL。根据 P1 和 P2,可能性分别为 0.32 和 0.50。其他部分敏感性函数的组合,例如 fVPD 和 fW 或 fT 和 fCI,在最佳模型中显示的可能性方面都非常接近。
不同气候植被类型之间的最佳部分敏感性函数有所不同(图S10)。例如,fTHorn 与极地植被、北方植被、温带草原和热带森林的相关性较高,但对于干旱气候和温带森林的植被不太重要。相反,线性峰值函数 fTMOD17 最适合干旱植被和温带森林。 fVPDPRELES(包括 CO2 效应)对于除干旱森林和干旱草原(其中具有延迟 VPD 功能的 fVPDHorn 更好)以及极地植被和热带森林以外的所有气候植被类型均较优。相比之下,涉及 W 滞后效应的 fWHorn 在干旱森林和草原上表现优于。 fWWeibull 为钟形或递减函数,更适合温带和热带草原。此外,对于每种气候植被类型,fCI、fCIEXP、fCIWang 和 fCIHorn 特别相似。 fCInone 出现的可能性非常低。 fLnone 出现在每种气候植被类型的最佳模型中,并且对于干旱森林、干旱草原和极地植被几乎与 fLTAL 一样好。一般来说,fVPDPRELES 和 fLTAL 对于大多数气候植被类型来说是必需的,而 fTHorn 对于寒冷地区来说是必需的,而 fVPDHorn 和 fWHorn 对于干旱地区来说是必需的。各种气候植被类型中的其他 fW 和 fCI 相似。
各种气候植被类型的主导控制
全球最佳模型(模型#1)显示了环境影响随气候植被类型变化的清晰模式。 T 是所有极地和北方地区的主要驱动因素(在第 2.6 节中定义),以及分别占温带森林和草原 54% 和 78% 的驱动因素(图 9a)。 T 的相对贡献(第 2.6 节中定义)在极地、北方和温带地点也是最大的,但很少高于 50%(图 9b)。 W 是 63% 干旱森林和 57% 干旱草原的主要驱动因素。同样,对于干旱地区,W 的相对贡献最大。 CI 和 VPD(包括 CO2)分别是 40% 热带森林和 100% 热带草原的主要驱动因素。 CI和VPD(包括CO2)分别对热带森林和热带草原的贡献最大。然而,与其他因素相比,大多数站点观察到的光饱和效应较少。特别是,它对干旱草原和极地地区的贡献很小。
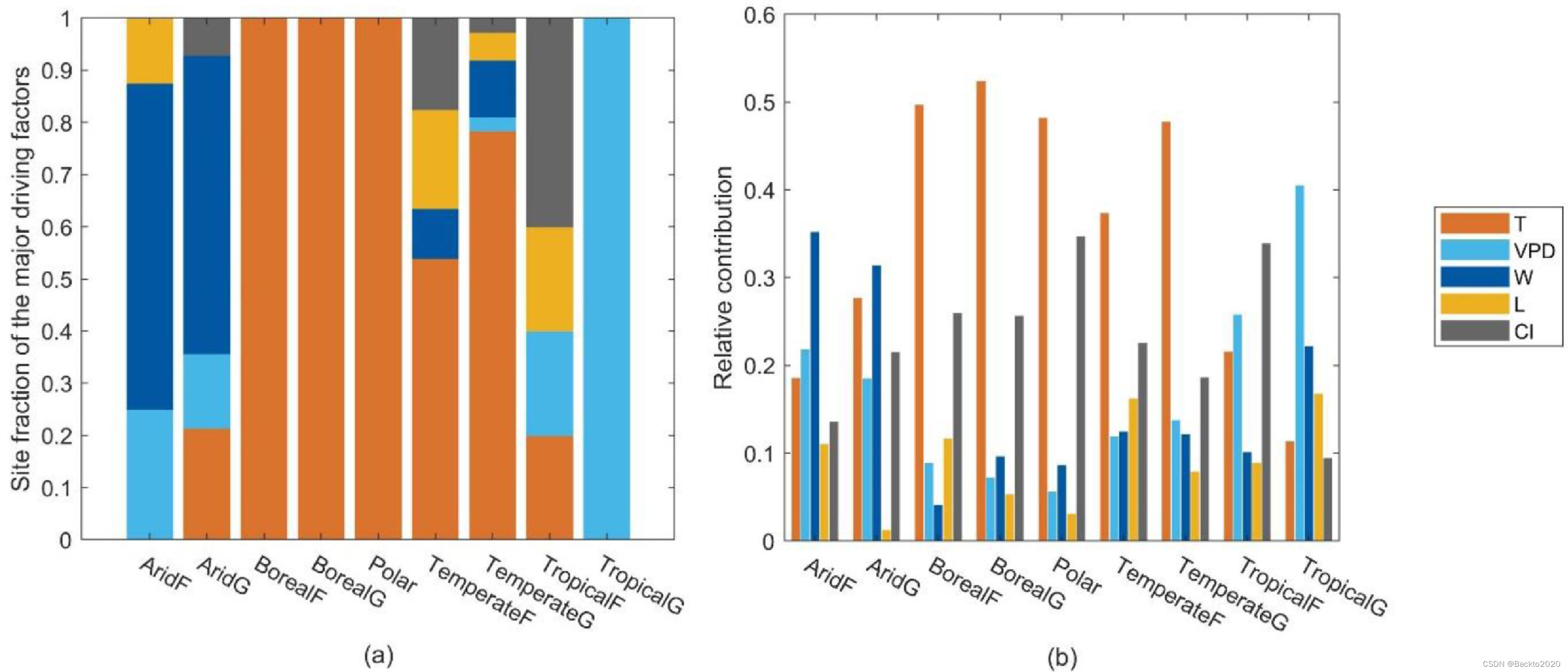 图 9. 不同气候植被类型的全球最佳模型中的环境敏感性:(a) 主要驱动因素的场地比例和 (b) 每个环境因素的相对贡献。两张图中,T(橙色)为温度,VPD(天蓝色)为水汽压亏缺,W(深蓝色)为土壤供水量,L(黄色)为光饱和度,CI(灰色)为云度指数。 “主要驱动因素”代表对站点层面的 LUE 影响最大的环境因素。 “相对贡献”是衡量环境因素对场地层面 LUE 总限制的贡献的指标,按气候植被类型进行平均。
图 9. 不同气候植被类型的全球最佳模型中的环境敏感性:(a) 主要驱动因素的场地比例和 (b) 每个环境因素的相对贡献。两张图中,T(橙色)为温度,VPD(天蓝色)为水汽压亏缺,W(深蓝色)为土壤供水量,L(黄色)为光饱和度,CI(灰色)为云度指数。 “主要驱动因素”代表对站点层面的 LUE 影响最大的环境因素。 “相对贡献”是衡量环境因素对场地层面 LUE 总限制的贡献的指标,按气候植被类型进行平均。
模型 #1 显示的环境对 LUE 的影响模式与站点最佳模型类似(图 S14)。两者最大的区别是在热带草原,CI是主要驱动因素,贡献最大,而不是VPD。此外,从全局最佳模型和站点最佳模型得出的 fT、fVPD、fW、fL 和 fCI 之间的 R2 在 73%、66%、56%、87% 和 93% 的站点上高于 0.7。因此,全局最佳模型一般可以反映LUE对环境因素的局部响应。
然而,环境对 LUE 建模的影响模式与环境因素与 GPPobs 或 LUEobs 之间的相关性并不一致。对于大多数气候类型,与 GPPobs 相关性最强的因素是 APAR,而不是 T 或 CI(图 S15a)。 W 和 T 也分别与干旱地区和热带地区、温带地区和寒冷地区的 GPPobs 密切相关。在环境因素和 GPPobs 之间的平均 R2 中也观察到了同样的情况(图 S15b)。相比之下,CI 与寒冷地点的 LUEobs 相关性最强(图 S15c)。 CI 和 LUEobs 之间的平均 R 在寒冷地点也是最大的(图 S15d)。然而,与 GPPobs 的相关性相比,T 与 LUEobs 的相关性极低。因此,GPP 与 APAR 更相关,而 LUE 与 CI 更相关。仅凭相关性并不能反映不同环境条件对 GPP 或 LUE 的主导控制。使用瞬时变量的相关性无法捕获寒冷和干旱地区温差和温差变化的延迟效应。
极端条件下的主导控制
直观上,在给定环境因素的极端条件下,LUE应该由该因素主导。例如,当T低于第10个百分位数时,它是大多数地点的主要驱动因素,并且与除干旱森林和热带植被之外的其他因素相比具有最大的相对贡献(图10中的第六行)。同样,一旦W低于第10个百分位数,也可能直接影响干旱植被和热带草原(图10a和b中的第四行)。
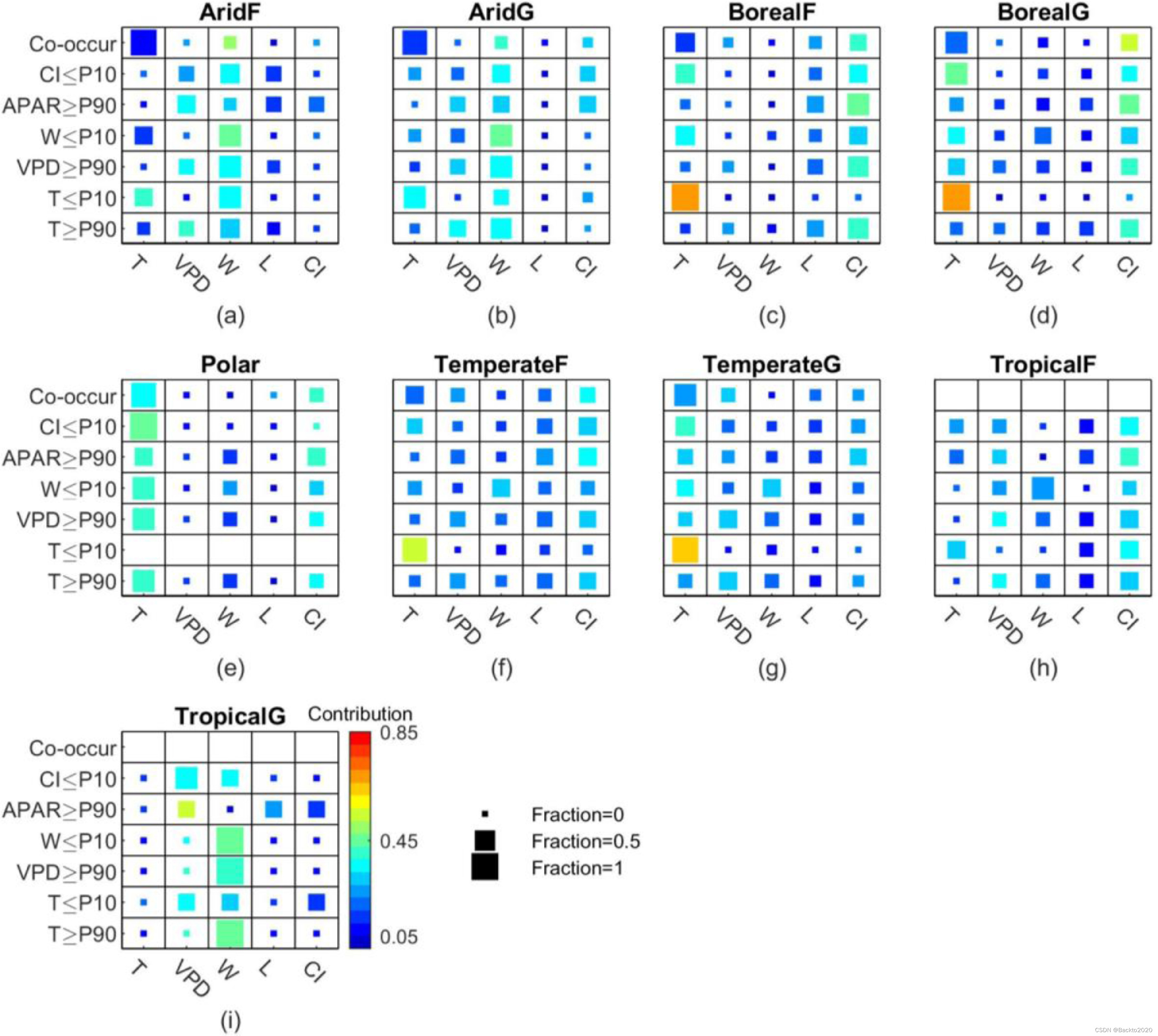 图 10. 不同气候植被类型极端条件下全球最佳模型中 LUE 的环境敏感性:每个“主要驱动因素”的场地分数(由方块大小表示)和每个环境因素的“相对贡献” (由方块的颜色表示)。 X轴代表环境因素:温度(T)、水汽压差(VPD)、土壤供水量(W)、光饱和度(L)、云量(CI)。 “同时发生”表示当 T≥90%、VPD≥90%、W≤10%、APAR≥90% 和 CI≤10% 时场地级别的环境条件。热带地区没有“同时发生”点(h 和 i)。 CI≤P10、APAR≥P90、W≤P10、VPD≥P90、T≤P10和T≥P90表示CI≤10%(晴空)、APAR≥90%(强光)、W≤10%的环境条件(土壤可用水量低)、VPD≥90%(大气需水量高)、T≤10%(低温)和T≥90%(高温)。正方形越大,环境因素对 LUE 影响最强的站点就越多。正方形越亮(黄色和红色),环境因素对 LUE 总限制的相对贡献就越大。例如,W是LUE的主要驱动因素,并且在干旱森林中CI≤P10时具有最大的相对贡献(a)。
图 10. 不同气候植被类型极端条件下全球最佳模型中 LUE 的环境敏感性:每个“主要驱动因素”的场地分数(由方块大小表示)和每个环境因素的“相对贡献” (由方块的颜色表示)。 X轴代表环境因素:温度(T)、水汽压差(VPD)、土壤供水量(W)、光饱和度(L)、云量(CI)。 “同时发生”表示当 T≥90%、VPD≥90%、W≤10%、APAR≥90% 和 CI≤10% 时场地级别的环境条件。热带地区没有“同时发生”点(h 和 i)。 CI≤P10、APAR≥P90、W≤P10、VPD≥P90、T≤P10和T≥P90表示CI≤10%(晴空)、APAR≥90%(强光)、W≤10%的环境条件(土壤可用水量低)、VPD≥90%(大气需水量高)、T≤10%(低温)和T≥90%(高温)。正方形越大,环境因素对 LUE 影响最强的站点就越多。正方形越亮(黄色和红色),环境因素对 LUE 总限制的相对贡献就越大。例如,W是LUE的主要驱动因素,并且在干旱森林中CI≤P10时具有最大的相对贡献(a)。
然而,我们发现 LUE 变异可能由不在极端条件下同时发生的环境因素主导,而当另一个因素处于极端条件下时,该环境因素也会同时发生。虽然 APAR 高于第 90 个百分位数(图 10a-i 中的第三行),但在一些干旱森林、北方植被、温带植被和热带植被地点,光饱和度是 LUE 的主要驱动因素。然而,T、VPD、W 或 CI 的相对贡献往往更大。相比之下,当其他因素之一处于极端条件下(T≥90%,VPD)时,CI 是主要驱动因素,对北方森林和温带森林地点(图 10c 和 f 中的第五列)贡献最大。 ≥90 个百分位,W≤10 个百分位,APAR≥90 个百分位,或 CI≤10 个百分位)。在干旱地区,W始终是主要驱动因素,贡献最大。
当环境因素同时处于极端条件下时,即T、VPD、APAR高于90%,CI低于10%时,大多数站点的主要驱动因素是T(图1中的“同时发生”)。 10)。然而,W 在干旱植被中的相对贡献和 CI 在其他气候植被类型中的相对贡献通常比 T 更大。此外,从站点最佳模型中可以观察到类似的模式(图 S16),这证明了上述环境对LUE影响的空间格局。
讨论
通过对各种环境因素组合及其敏感性函数的综合比较,我们的结果支持了最初的假设,即全球生态系统水平上GPP和LUE的变化会对T、VPD、W、L、CI和CO2的变化做出反应。这些环境敏感性函数反映了 GPP 对这些因素响应的潜在机制(第 4.1 节)。所选模型的鲁棒性和不确定性以及可能影响导出的环境效应的环境敏感性函数将在 4.2 节中讨论。第 4.3 节介绍了对所提出的方法及其对数据驱动的全球 GPP 估算的效用的总体评估。
总初级生产力对环境因素的响应
最初的假设是,GPP 对 T、VPD、W、L、CI 和 CO2 的响应都是显着且非线性的,尽管这些变量在时间和空间上都存在一定程度的相关性。此类响应的基本机制将在第 4.1.1-4.1.4 节中讨论。
温度
给定一定的光照,可以通过增加 T 来促进光合作用能力,但当 T 超过某个最佳值时会降低光合作用能力(Bernacchi 等,2003)。高温对GPP的限制与高VPD和低W有关(Fu et al., 2020; Sims and Bradford, 2001)。然而,钟形函数(例如,fTHorn)比 s 形函数或线性峰值函数更能代表 GPP 对 T 的全局响应,这反映出高温下 GPP 的降低不能通过考虑其他限制(例如, LUE 模型中的高 VPD 或低 W)(Bernacchi 等人,2003;Potter 等人,1993;Xiao 等人,2004)。仅在干旱地区,LUE 对 T 的响应可以通过线性峰值函数 fTMOD17 充分表示(图 S10f),因为其他限制因素(例如低 W)在炎热气候中占主导地位(图 10a 和 b;Guo等人,2015;张等人,2017a)。因此,即使结合其他因素,仅考虑 GPP 和 LUE 对 T 的正响应也会导致低估 GPP 对高温的敏感性(Horn 和 Schulz,2011a;Hwang 等,2008;Turner 等,2011)。 ,2006b)。
更灵活的 fT 可以是峰值函数或钟形函数(例如 fTCASA),但由于在北方和温带气候中缺乏 T 的滞后函数,因此无法代表 GPP 对 T 的全球响应。已经观察到 GPP 和 LUE 的滞后 T 效应(Pelkonen 和 Hari,1980),这归因于冬季低温造成的损害导致针叶光合作用能力降低的不完全恢复(Bergh 等,1998;Tanja 等) al.,2003;Yang 等,2020)、冻土解冻(Jarvis 和 Linder,2000)以及春季夜间霜冻的发生(Bergh 和 Linder,1999)。我们的结果表明,对于北方和温带气候(即寒冷生态系统;Horn 和 Schulz,2011a)的针叶林(Mäkelä 等,2004)和落叶林(Jarvis 等,2004)来说,滞后 T 效应是不可忽略的。 。全局最佳模型中 fTHorn 的滞后参数所表明的效果遵循 Mäkelä 等人 (2008) 的研究,而不是 Horn 等人 (2011) 的研究。极地和北方植被表现出明显的延迟响应,而温带气候的植被对温度变化的反应更快(图S17a),这可能是由于春季变暖和植物种类不同所致(Yang等人,2020)。虽然霜冻引起的光合作用能力下降也发生在秋季(Bergh et al., 1998; Hollinger et al., 1999),但本研究没有考虑到这一点,但严重的秋季霜冻造成的 GPP 损失并不那么严重。与春季一样(Bergh 等人,1998;Liu 等人,2018)。
蒸气压不足与土壤供水
植被通过控制气孔关闭来响应 VPD 和 W 限制,以减少水分流失,并避免木质部水势严重负值和栓塞。一般来说,在所有模型中,它通常被认为是涵盖水资源可用性影响的单一因素(McCallum 等人,2013;Running 等人,2004;Wang 等人,2017a;Wang 等人,2015; Yan 等,2017;Yuan 等,2007)。然而,LUE或GPP对VPD和W的响应不是同时的(参见图S11中的相关矩阵)并且取决于植被的等水性或非等水性(Roman等,2015;Tardieu和Simonneau,1998)。土壤水供应也会影响GPP对VPD变化的敏感性;例如,浇水良好的植物在关闭气孔之前可以承受较高的 VPD,而干旱胁迫的植物则显示出更早关闭气孔(Tardieu 和 Simonneau,1998)。我们的结果表明,应该考虑 W 和 VPD 效应,这与其他一些结果一致(Bagnara 等人,2015;Mäkelä 等人,2006;Mäkelä 等人,2008;Stocker 等人,2020;Turner 等人) ., 2006a; Wang 等人, 2018)研究(Bagnara 等人, 2015; Mäkelä 等人, 2008; Stocker 等人, 2020; Turner 等人, 2006a; Wang 等人, 2018)。
随着灵敏度的变化,增加 VPD 会降低 LUE,因此非线性函数(例如 fVPDHorn、fVPDTAL 和 fVPDWang)在表示 LUE 对 VPD 的响应方面比线性函数(例如 fVPDMOD17)表现更好(图 8)。我们观察到,VPD 效应在干旱气候中滞后,尤其是草原(图 S17c),因为干旱期后树冠必须重新发育绿色组织(Horn 和 Schulz,2011a)。木本植物通常可以从比草更深的土壤中提取水分(Kulmatiski 和 Beard,2013),从而导致不同的滞后。尽管如此,VPD 滞后并不像 W 那样明显(图 S17b),两者都可以反映植物对水分胁迫的适应(Flexas 等人,2009),但也可能反映了建模的 WAI 的过快响应降水和蒸散,或模型限制不代表更深的水源(Dorigo 等,2017)。
LUE 单调响应 W 的增加,但不同站点的灵敏度不同。因此,具有固定斜率的fWWAI不能代表GPP在全球尺度上对W的响应。对于北方植被与植被相关的水分状态,LSWI 是比 WAI 更好的指标,这可能是由于它具有捕捉叶片含水量即时变化的优势(Chandrasekar 等,2010)。非单调或递减的 fW fWWeibull 的可能性较低,反映出高 W 对光合作用没有限制,这与预期相反(例如,Stocker 等人,2018)。这可能是由于当前 FLUXNET 数据集中缺乏连续几天的浸水站点年数。
光饱和度和漫射辐射
强光通常伴随着晴朗的天空、高温、大气需水量高(图S11)和土壤供水量低(Piao等,2020)。因此,这些因素中的任何一个或组合都可能掩盖模型中光饱和度的影响,表明模型过度参数化相对于数据中给出的信息可能产生的影响(Ibrom 等人,2006)。此外,fL 和 fCI 参数之间的高度相关性(平均 R=-0.68)表明这两种效应之间的时间和空间同时性。原因是当CI较大时,相同量的光可以分布在更多的叶子上,整个冠层的光饱和度可以得到缓解(Gu et al., 2002; Ibrom et al., 2006; Knohl and Baldocchi, 2008;罗德里克等人,2001)。由于这些因素之间的相互作用,光饱和度似乎不像 LUE 模型中的其他驱动因素那么重要。它的部分影响可能会映射到其他响应函数的参数,反之亦然,特别是在具有许多函数和参数的模型上。这种现象可能会降低模型的可预测性。
然而,在高 NDVI 条件下,强光对 GPP 的影响最强(图 S18),例如热带森林(Propastin 等,2012)、温带森林和温带草原的一些地点。在全球模型中包含更多气候植被类型的范围内,有必要同时考虑光饱和度的影响(Mäkelä等,2008;Medlyn,1998)和CI(Wang等,2018)的影响。为了代表 LUE 对饱和光的响应,APAR 的反函数是合适的,因为另一项研究已经导出了类似的形式(Ibrom 等人,2008)。
增加 CI 可以改善 LUE,因为当总辐射不变时,它代表更大的漫射辐射比例。然而,由于 CI 和 PAR 之间的负相关性,在 GPP 中无法识别 CI 的重要性,这可能会抵消 CI 的积极作用。然而,更灵活的函数,例如fCIHorn,可以是单调递增、递减或钟形函数,不能更好地代表GPP在全球范围内对CI的响应。这表明不同气候植被类型的 LUE 和 CI 之间的单调正相关关系是最合适的,这与 Alton 的研究(2008)相反,但与其他研究一致(Cheng 等,2015;Gu 等,2002) )。
二氧化碳
作为光合作用的燃料,提高CO2浓度会以缓慢的速度为GPP施肥(Smith et al., 2016; Wenzel et al., 2016)。 CO2 施肥在时间和空间上都可能是异质的(Liu et al., 2016)。尽管大多数 LUE 模型没有考虑这种效应,但我们的结果反映了在 LUE 模型中包含 CO2 效应的优势。考虑到过去几十年大气中二氧化碳的大幅增加,它将转化为更广泛的 LUE 应用。
由于CO2施肥程度与VPD有关,VPD影响气孔关闭和气体交换,因此LUE模型能否捕获CO2效应取决于fVPD的函数形式。如果函数不合适(例如,fVPDP 不如 fVPDPRELES),模型性能可能会受到限制,并且对 CO2 升高的模型响应可能会有所不同(Haverd 等,2018;Smith 等,2016)。
此外,在 LUE 模型中使用空间变化的 CO2 强迫(Sun et al., 2018;Worden et al., 2021)对于探索 CO2 施肥对 GPP 空间变异性的影响是值得的。
模型的稳健性和不确定性
在下一节中,我们将讨论使用各种模型选择方法、随机选择的站点组和模型评估指标测试的模型鲁棒性(第 4.2.1 节)、模型参数估计、模型结构(第 4.2.1 节)所导致的所选模型可能存在的不确定性4.2.2)、观测数据(第 4.2.3 节)和其他潜在驱动因素(第 4.2.4 节)。
模型选择方法、站点和模型评估指标
模型评估通常对模型选择方法、给定的数据信息和评估指标敏感(Burnham 和 Anderson,2002;Burnham 等,2011;Efron,2014)。例如,VPM模型在Zhang等人的研究中就有不同的表现。 (2017b) (全局 R2=0.74 和 RMSE= 2.08 gC‧m−2‧d−1, 113 个位点) 和 Yuan 等人。 (2014)(空间 R2=0.44,157 个站点)。在我们的研究中,最佳模型随选择方法和引导站点(以站点年份表示)而变化,但不随识别最佳模型的评估指标或阈值而变化。最好的 20 个模型的排名对站点和站点年份(排名方差 = 134)比模型选择方法(排名方差 = 96)更敏感,突出了引导方法在跨站点选择稳健模型方面的重要性,从而在全球范围内。这里使用两个评估指标、六个阈值和三个独立模型选择方法选出的最佳模型是 77% 不同站点年份组(总共 200 个随机组)的最佳 1% 模型之一,也是100% 不同站点年份组。此外,它与使用 KS 检验(显着性水平 = 0.05)在每日和每周尺度上在任何站点年组中排名第一的任何其他模型没有统计学差异。因此,所选模型和部分敏感性函数对于当前 FLUXNET 数据集来说是稳健的。
模型参数及结构
模型不确定性的另一个来源是其参数。为了避免在参数校准中选择局部最优,我们使用了随机且无导数的进化全局搜索算法(Hansen 和 Kern,2004)。在基于位点采样和基于 ABC 的模型选择中都考虑了多个优化参数向量,以便可以选择对不同参数向量不敏感的鲁棒模型(Toni 和 Stumpf,2010)。然而,由于函数形式、驱动因素之间的相关性(图S11)和参数之间的相关性(参见第4.1.3节),有可能找到不确定的参数。使用特定位点的生物和非生物特性进行参数预测的实验(S1 节)显示参数预测存在很大偏差。因此,在没有 EC 测量的区域中进行参数外推可能会出现问题。尽管如此,据报道,霍恩模型的参数可以使用特定地点的属性进行推断(Horn 和 Schulz,2011b)。还可以预测位点平均 fT 和 fW,这是所有其他因素中相关性最小的环境因素和敏感性函数(图 S11)(图 S19),表明 GPP 对这些环境因素的响应模式是由生物物理控制的。和环境特征,并强调减少参数相关性的重要性。
如果忽略参数的不确定性,本研究选择的全局最佳模型在全球范围内的应用中具有最低的不确定性。在局部尺度或特定地点尺度上,生物群落最佳模型或地点最佳模型可能比全局最佳模型具有更低的不确定性。此外,模型的不确定性可以来自于结合部分环境敏感函数的模型结构。例如,根据李比希定律,部分灵敏度函数可以以最小形式组合(Minunno等,2015;Tian等,2020;Yuan等,2007)。它们还可以以求和形式(Horn 和 Schulz,2011a)或更复杂的数学形式(McCallum 等人,2013;Oliphant 等人,2011;Wang 等人,2017a)进行组合。此外,将吸收光分成阳光部分和阴影部分的两叶模型结构(He et al., 2013; Yan et al., 2017)是否能够比大叶模型结构更好地表示漫辐射对 GPP 的影响仍然未知。
涡度协方差与遥感数据不确定性
在本研究中,环境敏感性的模型选择和分析依赖于使用EC数据的评估。尽管质量控制变量可以过滤出良好的数据质量,但由于过滤方法导致的系统误差和随机误差仍然存在于 EC 测量中(Aubinet et al., 2012),从而影响模型的训练和评估(Jung et al., 2020)。此外,通量的划分方法会导致数据的不确定性。一方面,NEE的GPP估计算法仍需改进。例如,生态系统呼吸的夜间估计忽略了白天植物呼吸的抑制(Keenan et al., 2019),从而在 GPP 中产生了误差和不确定性(Reichstein et al., 2005)。另一方面,在干旱地区光降解的影响下,标准划分方法可能会产生物理上不切实际的 GPP 估计(Cleverly 等,2016)。我们注意到,一般来说,LUE 模型可以在缺乏无偏 EC 数据的情况下对 GPP 进行合理的估计。然而,即使在一些每日尺度上这些偏差很小的站点,误差也可能传播到年度尺度,尽管只是在少数站点。因此,对存在这些问题的地点的年际变异、趋势和年度 GPP 大小进行建模评估可能会存在问题。此外,全球 EC 塔的稀疏分布造成了某些气候和植被类型的差距(Alton,2020),从而影响了我们的模型在极端环境条件地区的适用性。
本研究采用与叶片叶绿素含量相关的遥感植被指数 NDVI(Rouse et al., 1974;Zhang et al., 2018)来表示 FAPAR,与 NDVI 呈线性关系(Myneni et al., 2018)。等,1997)。因此,校准的 εmax 值可能小于使用 FAPAR 的值,因为 FAPAR 通常被高估(Kanniah 等人,2009 年),但应该与使用 FAPAR 校准的 εmax 具有相似的空间模式(Horn 和 Schulz,2011a)。然而,遥感数据足迹和 EC 测量之间的不匹配可能会导致像素数据(即遥感图像像素)相对于点数据(即 EC 塔)缺乏代表性,特别是在破碎的景观(Cescatti et al., 2012)。因此,景观异质性可能会影响一些地点模拟环境敏感性的可靠性(Migliavacca 等,2015)。
初级总生产力的其他驱动因素
众所周知,林龄和缓慢变化的环境因素(例如氮沉降、二氧化碳施肥、气候变化和管理)会影响 GPP 对当地环境的响应(He 等,2012;Ma 等,2019;Medlyn,1998) )。给定较长时间序列的数据,可以通过校准每年的模型参数并分析其值的时间序列来证明这种效应(Wu et al.,2013;Wu et al.,2012)。在 FLUXNET 数据集中,有几个具有长时间序列的站点表现出长期趋势(Fernández-Martínez 等人,2017 年;Pilegaard 和 Ibrom,2020 年)。其中一些变异性可以通过部分灵敏度函数来捕获,包括 CO2 效应(第 4.1.4 节)。随着长期通量数据集的可用性不断增加(Baldocchi,2020),类似的研究可能揭示将这些长期动态纳入 LUE 模型的方法。
该方法的总体评价和未来应用前景
我们表明目前正在使用多种 LUE 模型。其中许多是从前人的基础上发展而来的(例如,He et al., 2013; Wang et al., 2015)。据我们所知,这是第一项评估所有可能的 LUE 建模方法潜力的研究,考虑了环境因素及其部分敏感性函数的每种组合,从而测试了 5600 个不同的 LUE 模型的全局和特定站点性能,并且在模型评估中考虑各种站点组。
结果证实,没有任何一个模型能够以最高的性能来模拟所有地点和年份,但是,全球最佳模型,即模型#1,其鲁棒性表现得令人惊讶。所选模型构成了尚未发布的模型结构,它证实了我们的初始假设以及本研究中提出的系统模型开发和基准测试方法的有用性。尽管 LUE 模型没有提供 GPP 响应的完整生理或生态描述,但模型 #1 反映了解释当前 FLUXNET 数据集中的变异性所需的表示。在局部尺度或特定地点尺度上,具有特定特征(即必要的环境因素和适当的敏感性函数)的地点最佳模型可能更有效,但在其他地点表现不佳,因为这些特征可能是多余的,而其他特征可能是多余的。丢失的。在全球范围内,模型#1 捕获了各个生态系统中必不可少的特征,这表明模型#1 可以作为未来模型选择的参考,以实现稳健的全球 GPP 估计。
与任何其他半经验或机械建模方法一样,模型#1 的全球或区域应用必然需要更大规模的强迫数据可用性和对模型参数变化的理解。全球最佳 LUE 模型具有在全球和区域范围内应用的潜力,因为在不同空间尺度上都有强迫数据(例如 CRUNCEP;Viovy,2018a)。然而,对于没有观测数据的地点,模型参数是未知的,这需要对控制其变异性的因素进行假设,例如植物功能类型(Running et al., 2004),或者另外的植被特性和气候条件(Carvalhais et al., 2010b) ),或假设在所有站点上优化单个参数集(Yuan 等人,2007)。参数外推是 LUE 模型以及所有其他类型参数方法(从经验模型到机械模型)的潜在挑战(Luo 和 Schuur,2020)。因此,要在全球范围内应用最佳模型或本地开发的任何其他模型,需要进一步研究参数外推方法。
结论
为了确定 GPP 的环境驱动因素和适当的部分敏感性函数,我们的研究重点是对先前报告的整体(n = 5600)和新的 LUE 模型进行全面和详尽的评估,该模型基于使用 196 个涡流协方差 CO2 通量站点的引导方法。选择了一个在全球范围内和各种气候植被类型上具有良好性能的稳健模型,该模型以前从未发表过。我们的结果表明,GPP 对温度、VPD、土壤水分供应、光饱和度、云量和 CO2 的响应都很显着,而且大多是非线性的。在寒冷地区和干旱地区,由于光合能力的恢复,温度和土壤水分供应对GPP的影响分别滞后。根据所选模型,温度和土壤湿度分别是寒冷和干旱环境中的主导因素。在不同的生态系统中观察到云量指数和光饱和度的影响。
我们的研究证实,在全球经验模型中通常被忽略的光饱和度、漫射分数和二氧化碳施肥应该在全球范围内的 GPP 估计中得到考虑。此外,非干旱地区的GPP对温度的影响呈非线性和钟形响应,而寒冷和干旱的生态系统则表现出温度和土壤湿度对GPP的滞后影响。未来在全球范围内使用最佳模型,必然需要对模型参数的外推进行进一步研究。为了对机械模型和数据驱动模型进行合理的评估和校准,本研究的建议是考虑不同生态系统观测数据的代表性以及模型结构和参数之间的等效性。















 3386
3386











 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









