







依靠锰的氧化实现细菌的化能自养生长
Bacterial chemolithoautotrophy via manganese oxidation
Nature [IF: 40.137]
DOI:https://doi.org/10.1038/s41586-020-2468-5
发表日期:2020-07-15
第一作者:Hang Yu1
通讯作者:Jared R. Leadbetter (jleadbetter@caltech.edu)1,2
主要单位:
1美国加利福尼亚理工学院地质与行星科学系(Division of Geological and Planetary Sciences, California Institute of Technology, Pasadena, CA, USA)
2美国加州理工学院工程与应用科学系(Division of Engineering and Applied Science, California Institute of Technology,Pasadena, CA, USA)
写在前面
分享标题:Nature:依靠锰的氧化实现细菌的化能自养生长
导读背景
锰氧化是一个研究时间较长的课题,最早在上世纪五六十年代就有相关文献研究,但是其核心问题锰氧化的生理学意义一直没有被解释,也就是说“细菌为什么要进行锰氧化”这个问题没有被回答。锰氧化在细菌的生理活动中起到了什么样的作用一直困扰着广大研究者,这次发表在《Nature》上的关于锰氧化的文章,发现了一种细菌锰氧化的生理学意义——化能自养。此外,作者分析这样的生化过程在厌氧-好氧交界的地方很可能广泛存在,比如卡斯特地貌中,具有相当的生物地理学意义,可以说为地学过程尤其是和Mn循环相关的过程提供了一些新的思路。此外,文章还发现了锰氧化过程中双菌的协作关系。恰巧,笔者所在的课题组曾经在2017年也发现了相同的双菌体系锰氧化现象(相关研究结果发表于当年ISME),虽然两者在生物学机理上不一致,但这更加印证了锰氧化的多态性与广泛性。
摘要
锰是地球上含量丰富的元素之一,有关锰氧化的过程一直是热点问题,原因在于其机理过程和生理学意义仍然需要更加详细的阐述和分子机理印证。本研究中,作者富集了一个锰氧化的环境,发现其中存在共培养的两株细菌并且共培养环境依靠锰(MnCO3)的氧化进行增殖。在共培养体系中,其中一株菌,所占比例较高,属于硝化螺旋菌门,但是和常见的硝化细菌(Nitrospira和Leptospirillum)具有较远的亲属关系,和目前数据库中的任何已知菌种均无完美匹配。另外一株菌(Ramlibacter lithotrophicus),所占比例较低,属于β变形菌门,但其单独存在时并不具有锰氧化能力。通过同位素探针标记CO2,发现碳固定过程依赖于锰氧化过程提供的能量。通过转录组学进一步揭示了代谢通路中胞外锰氧化过程在好氧条件下进行的能量储存与自养CO2固定过程之间的关系。这项研究拓展了目前已知的化能自养型细菌代谢过程,并且补充了生物地理学过程中锰循环过程(四价锰还原过程研究的较为广泛,机理较为清晰)。另外,这样的过程很可能会和其他的地理元素循环过程相互影响。
实验过程
锰氧化菌的培养
首先,作者根据计算推测锰氧化营养型微生物的存在是热力学可行的: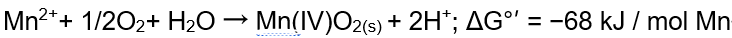
之后作者在瓶子中加入制备好的Mn(II)CO3和没有灭菌的自来水(接种源),并且随后发现体系中乳酪色的二价锰在几个月之后变成了黑色的锰氧化物。之后采用的培养体系中除了Mn(II)CO3,没有任何有机或无机的电子供体,比如说N源使用的是铵而不是硝酸盐,从而避免体系中引入硝化细菌。
接种来源:收集未经消毒的帕萨迪纳(美国加利福尼亚)市政饮用水(通常是来自地面和含水层的混合物),并在保持约21°C的空房间中开放、没有扰动地静置约10周(坐标:34.1367°,-118.1273°)
为了辨别生物氧化和非生物氧化过程,将之前的培养基质进行灭菌,然后在好氧条件下培养;另外考虑到锰氧化物本身也会氧化Mn(II),因此,又同时设置了灭菌体系中加入锰氧化物作为对照验证。以上两个体系均隔菌放置了一年,但是没有发现锰氧化现象。在接种体系中仅4周就能够发生明显的锰氧化现象(图 1a)。此外,体系中除了提供氧气还将温度保持在42℃。锰氧化的最佳发生温度是在34~40℃。而且还发现锰氧化过程会受到抗生素和50℃长时间加热环境的抑制。有意思的是,溶解态的二价锰当期浓度高于2mM后,会发生锰氧化的抑制作用。
图 1 MnCO3的生物氧化产生两个物种相关的氧化锰结核
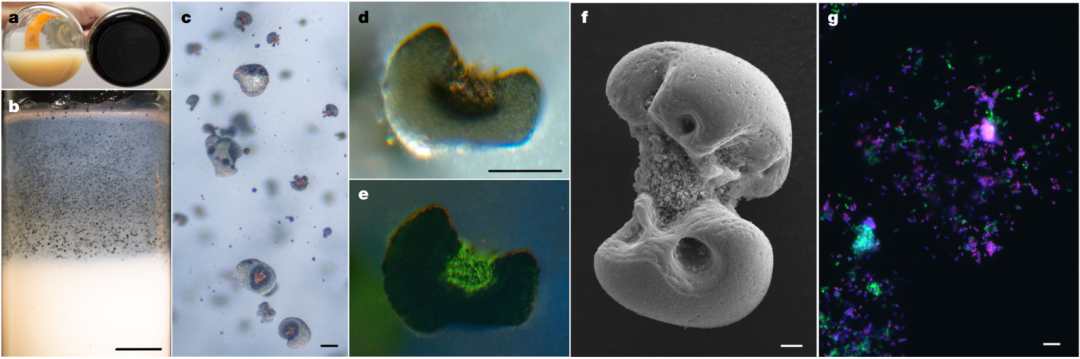
a 孵育后,将未接种的含有明亮,未反应的MnCO3的基础培养基对照瓶(左)与已接种有活物质的烧瓶中产生的附着的深色氧化物产物(右)进行比较。
b-e 在琼脂糖固化的MnCO3介质中产生的锰氧化物结核的显微镜观察。b 孵化接种有活物质的试管后,明亮的MnCO3颗粒云向暴露在空气中的弯月面澄清,同时产生较大的、离散的深色氧化物(在c中放大)。
使用iTag 16S rRNA基因测序对共培养基质进行鉴定,发现了70个物种,然后在MnCO3筛选型培养基中进行逐级稀释,最终获得了两株菌的共存环境,分别为命名为A菌和B菌,其相对丰度比例大约为7:1。
作者尝试单独分离出来A菌,但是没有成功,但是在其他异养培养基中成功分离出B菌,但是分离出来的B菌独自并不能进行锰氧化。作者猜测可能存在如下四种类型的双菌协作完成锰氧化的形式。同时,为了开展进一步的研究,A菌和B菌都进行了完整的基因组测序。
(笔者看完全文以后发现作者并没有纠结于证明两株菌之间的关系到底是哪种,不过纵观所有的实验过程,笔者认为第一种形式的可能性较高,也就是左上b这种形式)
锰氧化物表征
体系中生成的锰氧化物尺寸大约在20~500μm,并且在静态培养基和摇床上均能够生成锰氧化物,通常情况下是附着在玻璃瓶壁,菌体相互之间发生团聚现象,也能在琼脂糖固态培养基中发生(图1b)。通过傅里叶变化红外光谱对其进行表征发现其大多为松散结构,和水钠锰矿结构类似(无定型结构)。在荧光显微镜下观察到微生物被包围在锰氧化物的内部(图1d, e)。目前针对这种现象还没有进一步的解释和说明,究竟是什么原因造成的“包围”结构。通过FISH(原位荧光杂交)发现A菌的丰度高于B菌(图1g,A菌洋红色,B菌绿色),这也和之前测定的16S的结果相符。
生物生长速率与生物量积累
作者假设:如果其中一株菌或者两株菌均为化能自养型微生物,那么体系中应该存在以下情况,锰氧化的速率应该和Mn(II)生长速率趋势相当。通过实验发现,锰氧化速率存在两个阶段:每10.8天增加一倍和每6.2天增加一倍(图2a)。与此相符的是,A菌和B菌在Mn氧化过程中均呈现出指数型生长,并且两株菌的生长动力学相似(图2b)。此外,将锰氧化的量和细菌的生长过程进行相关性分析发现其中存在线性相关关系。通过估算得到,锰氧化产生的生物量大约是每摩尔Mn(II)氧化产生100~200mg干物质(dry biomass)。这样的生物量和亚硝酸盐氧化驱动的化能自养菌相当。也就是说Mn氧化产生的生物量足以支持微生物进行生长,并且和其他类型的化能营养型微生物具有一定的可比性。
图2. Mn氧化偶联物种A和B的共培养生长
Mn(II) oxidation coupled to co-culture growth of species A and species B
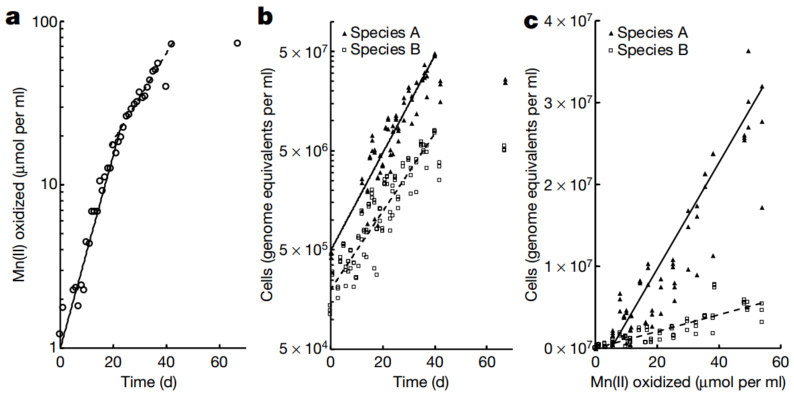
图3. 物种A的系统发育分析和代谢重建
Phylogenetic analysis and metabolic reconstruction of species A (‘Candidatus Manganitrophus noduliformans’)
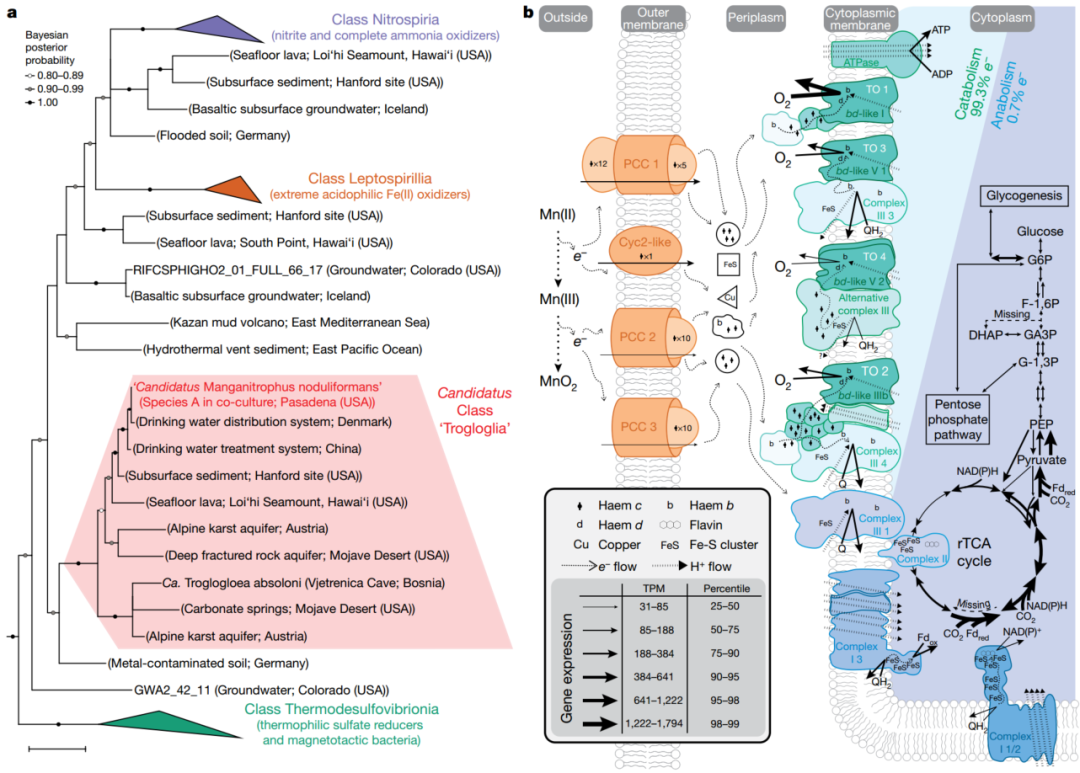
系统发育分析
从系统发育的角度对A菌和B菌进行鉴定和分类,见图3a。
A菌和Nitrospira, Leptospirillum具有较远的亲缘性,但和目前已知的Nitrospira菌门中其他菌属的核酸相似性低于84%。并且A菌的基因组中并不含有已知的化能(铵,硫,亚硝酸)自养相关的基因。从序列同源性角度出发,A菌和饮用水,地下水以及海洋的好氧-厌氧区的相关微生物亲缘关系较近,另外,A菌和一株通过宏基因组组装(MAGs)的菌Candidatus ‘Trogloglia’ 存在88%的序列相似度,而后者是在地下水环境中发现的。笔者认为以上的系统发育分析表明A菌的生活环境应该是寡营养条件,当然这也符合其自养微生物的生存环境特征。
相比较之下,B菌的系统发育路径较为清晰,属于β变形菌门的Ramlibacter属,并且B菌生长依靠H2和O2,并编码氧化脱氢酶基因,还原性硫氧化基因(Sox和DsrMKJOP),卡尔文循环基因以及某些厌氧呼吸通路基因。
转录组分析依靠锰氧化作用的细菌生长过程
生信分析使用Sleuth v.0.30.0对不同的RNA样品进行最终分析和标准化,同时重建两个物种或仅物种A的基因组。
结果显示从整体上看,有关细菌活动的相关基因(如鞭毛合成)并没有显著表达,相比之下,生物合成相关基因大量表达。
接下来,分辨哪些基因可能和锰氧化功能相关。A菌中存在四个基因簇,它们和铁氧化以及金属还原过程必需的胞外复合物合成基因相关。推测这些基因簇可能和锰氧化产生的电子传递相关,它们将电子从细胞外转移到到细胞周质空间,因而锰氧化物是在胞外产生的,而不是在细胞内部。笔者发现,这一点和以往研究锰氧化中发现的锰氧化依靠外泌的多铜氧化酶以及超氧化物的分子机理不一样。
接下来对电子传递通道相关基因进行分析,发现在铁氧化细菌中,胞外细胞色素C起到了初始氧化剂以及电子载体(electron carrier)的作用,功能基因为Cyc2或Cyt572,而A菌中该基因的同族基因进行了表达;同样的,在一些光营养型铁氧化菌和其他的化能铁氧化菌中,通常孔蛋白-细胞色素C结合的模型在电子传递过程中至关重要,菌A也能够表达相关基因,PCC1,PCC2和PCC3.
菌B在共培养的环境中大量表达了MtrA-MtrB-MtrC的孔蛋白-细胞色素电子传递通道,这一点和常见的金属还原型细菌Shewanella十分相像。
当电子传递到细胞周质中以后,他们如何在这个区域中发挥作用并产生能量(ATP)就显得尤为重要了。A菌中编码了很多氧化酶类基因,均是链接醌库和氧气的相关反应酶类。在转录组测序中,这些基因大量表达。其中最高表达的是‘TO 1’,它和氨氧化菌以及亚硝酸盐氧化菌中的相关过程中的氧化酶类似。TO 2~4也进行了表达,虽然表达量不及TO 1,但是它们和FeS的氧化过程相关,这一点和作者之前的推测相符。此外,作者提出本研究中的不足在于,这些酶类是否亲氧还需要进一步实验验证;而且由于锰氧化是“单电子两步”反应,通常来说,产生三价锰的阶段是一个限速步骤,因而未来研究中仍然值得考虑这两步中每一步的生理过程和意义。
锰氧化结合CO2固定
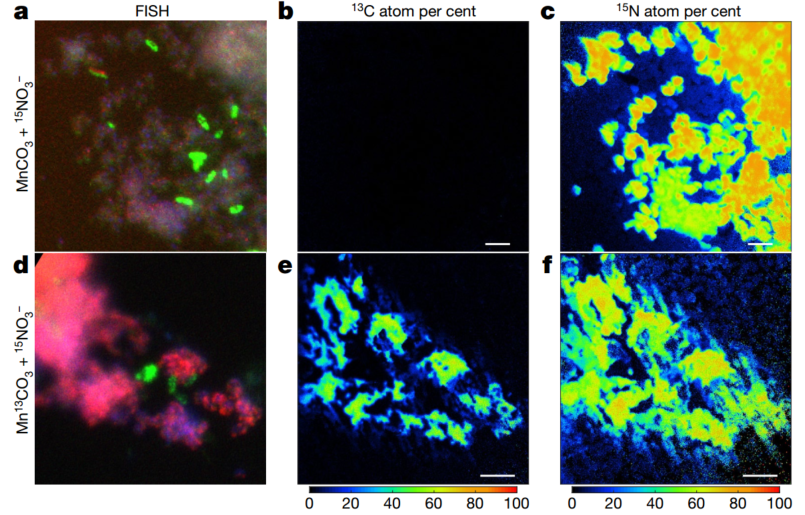
由于证实了共培养体系中是化能自养过程,那么生物量的产生离不开碳固定,作者推测其来源是CO2,因而考察锰氧化过程是否结合了CO2固定。通过同位素标记13C-MnCO3,同时也用15N标记硝酸盐,目的是追踪新的生物量合成过程,在FISH和nanoSIMS下进行观察。A菌仍然生长情况要好于B菌,并且利用13C进行了生物量的合成过程(图4)。这也和转录组结果相符,A菌中进行TCA循环的相关基因低表达,也就是说其对有机碳的固定缺乏,而B菌却可以。
图4. 自养二氧化碳固定的稳定同位素标记
Stable isotope probing of autotrophic CO2 fixation
讨论
关于化能锰氧化微生物是否存在,在过去的一个世纪中一直争论不止,这篇研究证实了他们的存在,并且从细胞、系统发育、转录组以及同位素示踪等不同的角度对其进行了详细的考察。在对A菌进行了物理外观,系统发育,基因组等多方面的考察之后,将其命名为‘Candidatus Manganitrophus noduliformans’。
锰氧化过程与生物生长的关联性使得这个过程具有重要的环境学意义。以目前实验观测到的生长速率和锰氧化速率来推算,AB两株菌在不受生长限制的环境中,两年内产生的锰氧化物的量与全球的锰储备相当,相关计算公式见原文。通过系统发育推断,A菌的相近功能的微生物可能存在于地下或喀斯特地貌环境中,包括厌氧-好氧交界处,如汉德福沉积物中。在这样的环境中,Mn的循环过程被建立:好氧锰氧化与锰氧化物呼吸,也因此会刺激了本质上发生的电子转移过程。这也意味着,C,N,S,Fe,H和氧等元素的循环过程会受到锰循环过程的影响,并且相互之间产生作用。
编译:王东麟 中科院生态环境研究中心
责编:马腾飞 南京农业大学
审核:刘永鑫 中科院遗传发育所
Reference
Hang Yu,Jared R. Leadbetter. Bacterial chemolithoautotrophy via
manganese oxidation.Nature volume 583, pages453–458(2020) https://doi.org/10.1038/s41586-020-2468-5
猜你喜欢
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
文献阅读 热心肠 SemanticScholar Geenmedical
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份,另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。

学习16S扩增子、宏基因组科研思路和分析实战,关注“宏基因组”
 点击阅读原文,跳转最新文章目录阅读
点击阅读原文,跳转最新文章目录阅读

















 2479
2479

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









