






中国南海冷泉细菌和产甲烷菌协同作用代谢甜菜碱产甲烷
Bacteria and archaea synergistically convert glycine betaine to biogenic methane in the formosa cold seep of the South China Sea

Article,2021-9-7,msytems, [IF 7.32]
DOI:10.1128/mSystems.00703-21
原文链接:https://journals.asm.org/doi/10.1128/mSystems.00703-21
第一作者:Lingyan Li (李凌燕);Wenting Zhang(张文婷);Shengjie Zhang(张圣杰)
通讯作者:Xiuzhu Dong (东秀珠);Lingyan Li (李凌燕);
合作作者:Lei Song (宋磊);Qinglei Sun (孙庆磊);Huan Zhang (张峘);Hua Xiang (向华)
主要单位:中国科学院微生物研究所(State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing, China)
中国科学院大学(University of Chinese Academy of Sciences, Beijing, China)
中国科学院海洋研究所(Institute of Oceanology, Chinese Academy of Sciences, Qingdao, China)
- 摘要 -
冷泉是由深海深部沉积物中的甲烷及其他有机质流体向海底渗漏或喷发而形成的独特低温环境。冷泉周围沉积物中生活着大量的自养微生物。前期化学分析发现,冷泉甲烷为生物来源甲烷。但其优势产甲烷途径及功能菌群尚不清楚。我们以中国南海冷泉为研究对象,通过添加产甲烷前体物质,表明冷源甲烷主要有细菌和产甲烷菌协同代谢甜菜碱产生,16S rRNA扩增子及宏基因组分析发现,冷泉含有丰富的甜菜碱还原酶(grdH)及甜菜碱甲基转移酶(mtgB)功能基因簇。进一步的“微宇宙”富集分析表明,甜菜碱代谢富集物种优势的细菌和古菌分别为:Oceanirhabdus 和Methanococcoides。通过纯培养分离获得优势的细菌和古菌新种分别为:Oceanirhabdus seepicola 和 Methanococcoides seepicolus。Oceanirhabdus seepicola 含有甜菜碱还原酶(grdH)功能基因簇,Methanococcoides seepicolus 含有甜菜碱甲基转移酶(mtgB)功能基因簇,表明南海冷泉甲烷主要有细菌和产甲烷菌协同代谢产生。
- 引言 -
冷泉是由深海深部沉积物中的甲烷及其他有机质流体向海底渗漏或喷发而形成的独特低温环境。冷泉周围沉积物中生活着大量的自养微生物。前期化学分析发现,冷泉甲烷为生物来源甲烷。目前已报道的海洋生境可以代谢三甲胺产甲烷的古菌包含拟球菌属(Methanococcoides)和八叠属 (Methanosarcina); 直接代谢甜菜碱产甲烷的古菌很少,包括Methanococcoides sp. strainQ3C和Methanolobus sp. strain B1d。这两株古菌可利用甜菜碱甲基转移酶代谢产甲烷。甜菜碱产甲烷主要通过细菌还原甜菜碱产生三甲胺,三甲胺被古菌代谢产生甲烷。目前仅有报道从盐沼泽沉积物富集到的Pelobacter 和Methanococcoides 协同代谢甜菜碱产甲烷。本研究通过宏基因组分析发现,南海冷泉沉积物及甜菜碱富集物中含甜菜碱还原酶编码基因(grdH)及甜菜碱甲基转移酶编码基因(mtgB)。进一步的“微宇宙”富集分析表明,甜菜碱代谢富集物种优势的细菌和古菌分别为:Oceanirhabdus 和Methanococcoides。 通过纯培养分离获得优势的细菌和古菌新种分别为:Oceanirhabdus seepicola 和 Methanococcoides seepicolus。两株菌在低温底物互配,发现可协同代谢甜菜碱产甲烷。并且发现甲烷菌代谢产物二甲基甘氨酸(DMG)具有冷保护作用。
- 结果 -
中国南海冷泉甲烷来源于甜菜碱
Glycine betaine supports biogenic methane production in the Formosa cold seep
通过“微宇宙”富集,添加产甲烷菌不同底物,发现甜菜碱及三甲胺来源产甲烷途径是南海冷泉优势产甲烷途径。通过添加抑制细菌生长的抗生素发现,甜菜碱产甲烷途径主要由细菌和古菌协同完成(图1a)。碳同位素分流实验表明,原位排放甲烷δ13C(-41.82 ∽-61.49‰)比甜菜碱富集的甲烷(-47∽-71.56‰)略重,但介于乙酸代谢产甲烷(-9∽-35‰)与氢还原二氧化碳产甲烷(-28∽-79‰)之间。推测甜菜碱来源产甲烷途径是中国南海冷泉的优势产甲烷途径(图1b)。
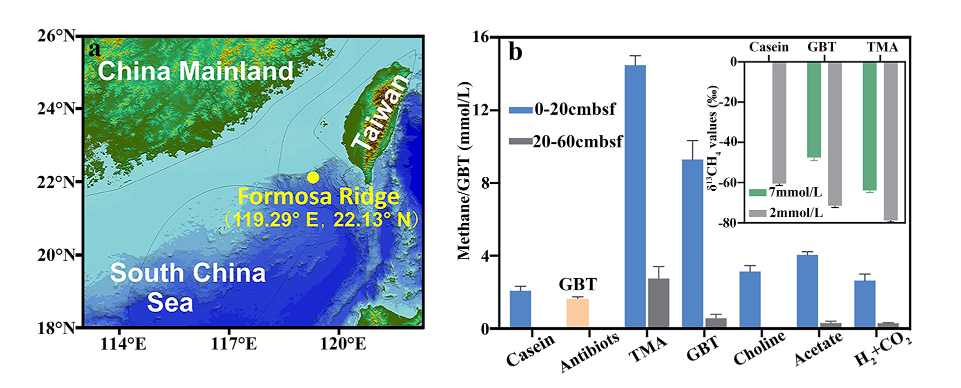
图1:南海冷泉底泥产甲烷前体物质测定
a: 南海冷泉底泥样品采集地理位置;b: 厌氧富集产甲烷前体物质,分别添加20mM 酪蛋白(Casein),三甲胺(TMA),甜菜碱(GBT),胆碱(Choline)及乙酸(Acetate),0.1MPa氢气:二氧化碳(1:4);氨苄青霉素和卡纳青霉素(1毫克/毫升);内插图为南海冷泉底泥添加酪蛋白、甜菜碱及三甲胺的甲烷同位素分馏。
甜菜碱厌氧富集物中检测到未培养梭菌及拟球菌
Uncultured Clostridiales spp., Methanococcoides spp., and the genes involving in GBT metabolism are enriched in GBT-amended cold seep microcosms
甜菜碱(20mM)作为唯一碳源,温度为18℃来富集南海冷泉表层沉积物产甲烷古菌及互作细菌。16S rRNA扩增子测序表明,沉积物原位细菌主要由Pseudomonas, Sulfurovum,及未知梭菌组成;甜菜碱富集物培养至41天,细菌菌群发生改变,主要由Vallitalea,梭菌门的 Tyzzerella 和Oceanirhabdus,绝对厌氧的 Draconibacterium组成(图2a)。甜菜碱富集物中优势古菌菌群同样发生变迁。原位古菌主要由ANME-2a (28.91%),ANME-1b (31.25%) ,Thaumarchaeota (24.22%)组成,甜菜碱富集物中主要以Methanococcoides(22.50%),Methanobacterium (18.83%),uncultured Bathyarcheaia(30.49%)组成。甜菜碱富集物培养至81天后,uncultured Bathyarcheaia消失,古菌大部分由Methanococcoides组成(图2b)。
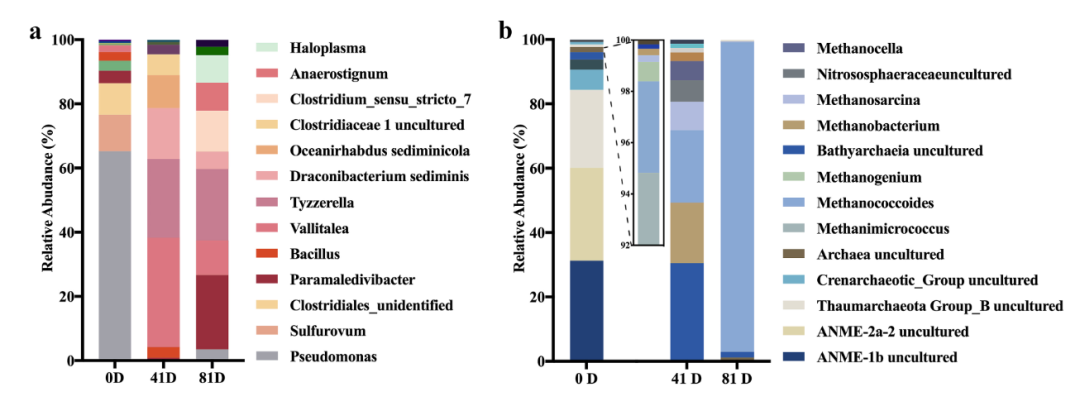
图2:南海冷泉沉积物原位及甜菜碱富集物中优势细菌古菌组成。
a: 优势细菌菌群组成;b: 优势古菌菌群组成。
甜菜碱的厌氧降解主要包括甜菜碱还原酶(grdHI)及甜菜碱甲基转移酶 (mtgB)。细菌利用甜菜碱还原酶代谢甜菜碱产生三甲胺和乙酸,三甲胺进一步被甲基类产甲烷菌代谢产甲烷。甜菜碱也可被甲基转移酶直接代谢产生二甲基甘氨酸(DMG)。运用宏基因组学方法,鉴定到南海冷泉沉积物富含甜菜碱还原酶(grdHI)功能基因簇(图3a)及甜菜碱甲基转移酶(mtgB)基因簇(图3b)。宏基因组学分析获得总计26个,6种类型含有grdHI功能基因簇的contig。其中类型Ⅰ,Ⅲ,Ⅴ在甜菜碱富集物中富集了约100倍(图3a)。同时获得总计503个,6种类型的甜菜碱甲基转移酶(mtgB)基因簇,其中类型Ⅰ,Ⅳ,Ⅴ,Ⅵ在甜菜碱富集物中富集了约10到100倍(图3b)。

图3 南海冷泉原位底泥及甜菜碱富集物功能基因 grdH 和 mtgB的类型及丰度
a: 功能基因 grdH的类型及丰度 b: 功能基因 mtgB 的类型及丰度。
甜菜碱富集分离获得梭菌及拟球菌新种
Oceanirhabdus seepicola sp.nov and Methanococcoides seepicolus sp.nov. were isolated from glycine betaine microcosm
甜菜碱富集物分离获得Oceanirhabdus seepicola sp.nov(CGMCC1.17897,JCM34232),与Oceanirhabdus sediminicola NH-JN4 同源性为94% (图4a)。Oceanirhabdus seepicola sp.nov基因组含有甜菜碱还原酶基因簇 (grdHI)(图4b)。Oceanirhabdus seepicola sp.nov可以甜菜碱为唯一碳源生长,同时检测到三甲胺的产生(图4c)。生长温度范围为(4到30°C),最适生长温度为18°C。同时,在甜菜碱富集物中分离得到Methanococcoides seepicolus sp.nov.(CGMCC1.17896),与Methanococcoides alaskenses 和Methanococcoides burtonii的同源性为99%,与Methanococcoides burtonii基因组ANI同源性为91%,所以将Methanococcoides seepicolus定为新种(图5a)。菌株LLY基因组编码甜菜碱甲基转移酶功能基因簇(图5b)LLY可以直接代谢甜菜碱产甲烷(图5c)。

图4:Oceanirhabdus seepicola sp.nov基因组编码甜菜碱还原酶功能基因簇且可以代谢甜菜碱产三甲胺。
a: Oceanirhabdus seepicola sp.nov 系统发育树的构建;b: Oceanirhabdus seepicola sp.nov 基因组编码甜菜碱还原酶功能基因簇; c: Oceanirhabdus seepicola sp.nov 代谢甜菜碱产三甲胺。
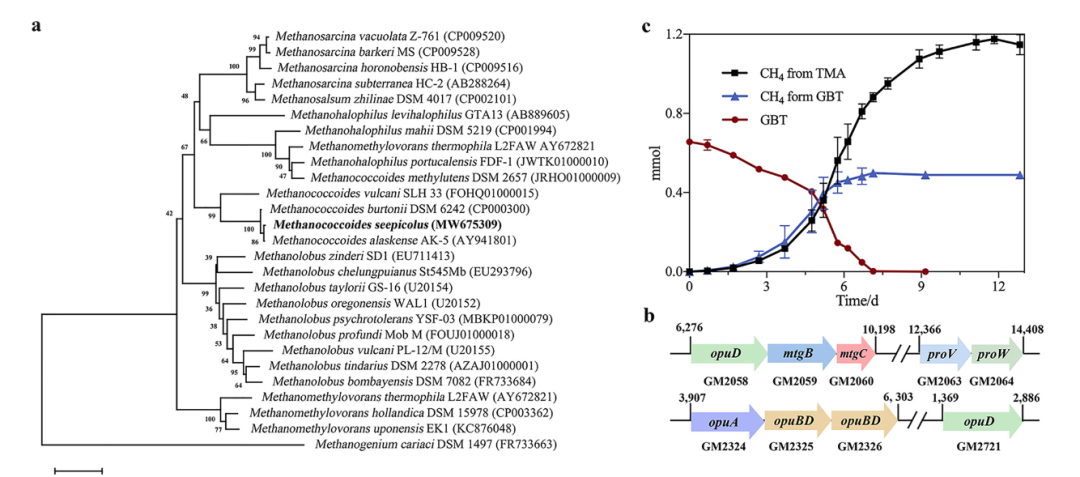
图5:Methanococcoides seepicolus sp.nov.基因组编码甜菜碱甲基转移酶功能基因簇且可代谢甜菜碱产甲烷。
a: Methanococcoides seepicolus sp.nov.系统发育树构建;b: Methanococcoides seepicolus sp.nov.基因组编码甜菜碱还原酶功能基因簇;c: Methanococcoides seepicolus sp.nov.代谢甜菜碱产甲烷。
梭菌新种与拟球菌新种协同代谢甜菜碱产甲烷
Oceanirhabdus seepicola and Methanococcoides seepicolus syntrophically convert GBT to methane
为了验证Oceanirhabdus seepicola 和 Methanococcoides seepicolus和能否协同代谢甜菜碱产甲烷,将两株菌互配,以甜菜碱为底物,18℃培养15天,发现甜菜碱全部代谢完,同时检测到了三甲胺的生成。由于互配体系前2天检测到大量的三甲胺,但甲烷量较低,推测甜菜碱优先被细菌 Oceanirhabdus seepicola代谢产生三甲胺,三甲胺后续被Methanococcoides seepicolus代谢产生甲烷(图6a)。进一步的RT-PCR实验证明,菌株Oceanirhabdus seepicola的编码甜菜碱还原酶的基因grdH在互配体系中表达量显著高于纯培养(图6b,c)。Methanococcoides seepicolus的编码甜菜碱甲基转移酶的基因mtgB在互配体系及纯培养物中表达没有显著差异,但编码三甲胺甲基转移酶的功能基因(mttB)在互配体系表达显著上调(图6b,d)。
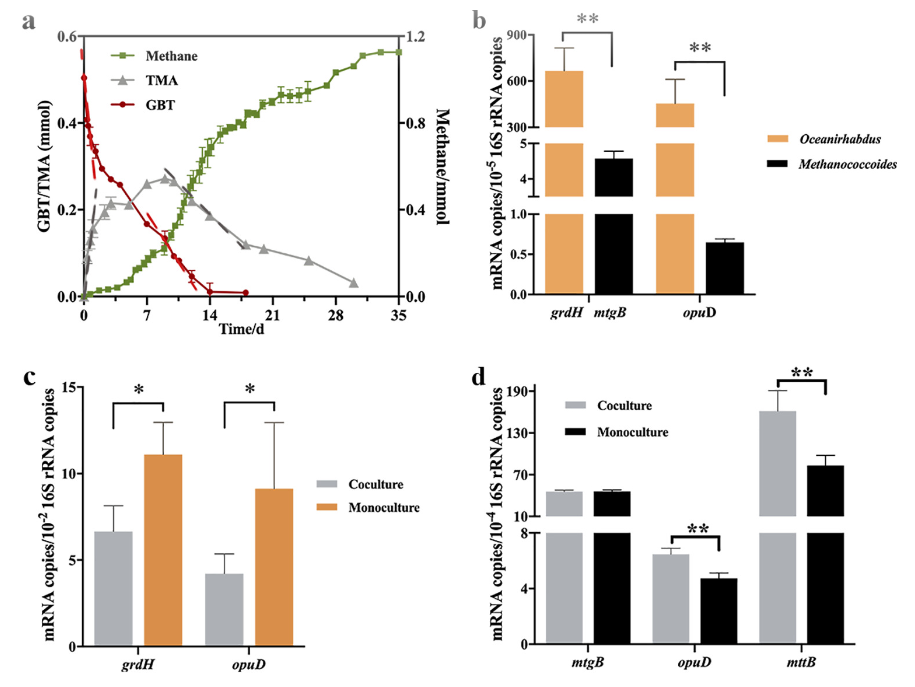
图6:Oceanirhabdus seepicola 和 Methanococcoides seepicolus 18℃协同代谢甜菜碱产甲烷。
a: Oceanirhabdus seepicola 和 Methanococcoides seepicolus协同代谢甜菜碱产甲烷;b,c,d: 两株菌在互配体系及纯培养物中功能基因grdH, mtgB,opuD的mRNA定量。
从甜菜碱产甲烷的富集物中获得含有grdH功能基因的细菌新种-居冷泉海洋杆菌(Oceanirhabdus seepicola sp. nov.),及含有mtgB功能基因簇的古菌新种--居冷泉甲烷拟球菌(Methanococcoides seepicolus sp. nov.)。两个冷泉新种的共培养可在低温下将甜菜碱转化为甲烷。进一步实验发现,甜菜碱脱甲基的产物-二甲基甘氨酸(DMG)对甲烷古菌发挥冷保护作用,促进其在冷泉原位温度4℃ 产甲烷。因此,冷泉甜菜碱发挥产甲烷底物及冷保护剂双重功能。解释了在温度很低的冷泉中微生物通过代谢偶联获得能量及嗜冷机制。根据该工作的发现,我们提出冷泉生物源甲烷的代谢模型(图7)。
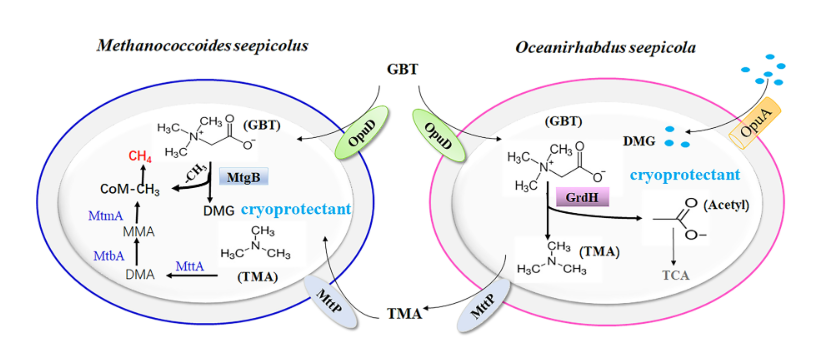
图7: 细菌新种Oceanirhabdus seepicola 和 古菌新种Methanococcoides seepicolus协同代谢甜菜碱产甲烷模型
- 讨论 -
甜菜碱作为渗透压保护剂广泛分布于海洋及盐碱生境。甜菜碱和二甲基甘氨酸均在嗜盐生境产甲烷古菌中发现。二甲基甘氨酸被报道抵抗芽孢杆菌热胁迫。此研究我们首次报道甜菜碱被甲基转移酶代谢产生二甲基甘氨酸,二甲基甘氨酸和甜菜碱可作为冷泉微生物的冷保护剂。而且,包括墨西哥湾在内的其它冷泉中均检测到了甜菜碱还原酶及甲基转移酶编码基因簇,暗示甜菜碱作为冷保护剂及产甲烷前体物质广泛分布于全球冷泉生境。
参考文献
Li, Lingyan*#, Zhang, Wenting#, Zhang, Shengjie#, Song, Lei, Sun, Qinglei, Zhang, Huan, Xiang, Hua, Dong, Xiuzhu*. (2021) Bacteria and archaea synergistically convert glycine betaine to biogenic methane in the formosa cold seep of the South China Sea. mSystems.
- 作者简介 -
第一作者

中国科学院微生物研究所
李凌燕
助理研究员
李凌燕,中国科学院微生物研究所助理研究员,主要关注厌氧环境基因组学、厌氧菌的分离培养、产甲烷古菌与细菌互作等,目前发表SCI 12篇,以第一作者发表了4篇,mSystems,Environmental Microbiology,Applied Microbiology and Biotechnology 等。

中国科学院微生物研究所
张文婷
直博生
张文婷, 中国科学院微生物研究所直博生。主要关注厌氧生境产甲烷古菌与细菌互作,目前发表SCI 4篇,以第一作者发表1篇。

中国科学院微生物研究所
张圣杰
博士研究生
张圣杰,中国科学院微生物研究所博士研究生,感兴趣的研究方向:极端环境微生物组与病毒组,难培养微生物的富集培养。
通讯作者

中国科学院微生物研究所
东秀珠
博士,研究员
东秀珠,博士,中国科学院微生物研究所研究员,博士生导师。2000年获得国家杰出青年基金,2004年中国科学院“百人计划”入选者。目前担任《微生物学报》副主编、 Frontiers: terrestrial microbiology、ISME journal、《微生物学通报》、《中国农业生物技术》和《生物多样性》编委。证明了甲酸盐在互营细菌和甲烷古菌之间的传递是有机物厌氧降解的重要机制;发表了25个细菌的新种并建立了7个新属;建立了以热激蛋白HSP60基因为基础的双歧杆菌发育系统;证明了碳酸氢盐是触发严格厌氧的互营细菌产芽孢的物质;发现口腔链球菌新的抗氧胁迫途径;首次发现古菌的群感效应系统及其调控的生理过程、在促进颗粒污泥形成和提高污水处理效率的作用;证明转录后调控在甲烷古菌糖体合成中的新机制,发现首个古菌冷适应蛋白及其RNA伴侣功能。在Nucleic Acids Res, ISME J,Msystems, PLoS Genetics, Mol Microbol, Environ Microbiol, J Bio Chem, J Bacteriol, Appl Environ Microbiol, Appl Microbiol Biotech, FEMS Microbiol, Int J Syst Evol Microbiol等刊物发表SCI论文100多篇,并主编专著3本。
猜你喜欢
iMeta简介 高引文章 高颜值绘图imageGP 网络分析iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读

















 345
345

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









