







研究论文
● 期刊:Nature Microbiology (IF:20.5)
● DOI:https://doi.org/10.1038/s41564-024-01891-8
●原文链接: https://www.nature.com/articles/s41564-024-01891-8
● 第一作者:Lejla Daruka, Márton Simon Czikkely, Petra Szili, Zoltán Farkas
● 通讯作者:Csaba Pál (pal.csaba@brc.hu);Balázs Papp(papp.balazs@brc.hu);Bálint Kintses(kintses.balint@brc.hu)
● 发表日期:2025-1-13
● 主要单位:
匈牙利塞格德大学, 匈牙利塞格德国家生物技术实验室, 匈牙利塞格德HCEMM-BRC代谢系统生物学实验室, 匈牙利分子医学卓越中心, 匈牙利布达佩斯厄尔特厄特沃什·洛兰大学, 匈牙利分子医学卓越中心, Delta Bio 2000有限公司, 匈牙利塞格德HUN-REN生物研究中心, 英国埃克塞特大学等。
摘要Abstract
尽管抗生素研发不断推进,细菌耐药性的进化可能使候选抗生素失效。本研究对比了2017年后引入或正在开发的13种抗生素与现用抗生素的体外耐药性产生情况。实验室进化结果显示,大肠杆菌(Escherichia coli)、肺炎克雷伯菌(Klebsiella pneumoniae)、鲍曼不动杆菌(Acinetobacter baumannii)和铜绿假单胞菌(Pseudomonas aeruginosa)等优先关注的革兰氏阴性ESKAPE病原体在抗生素暴露后60天内即产生临床相关的耐药性。研究发现,病原体的自然种群中已存在耐药突变,这表明通过选择预存的细菌变体,耐药性可在自然环境中出现。功能性宏基因组学分析显示,针对候选抗生素的可移动耐药基因在临床细菌分离株、土壤和人体肠道微生物群中广泛存在。总体来看,候选抗生素对耐药性发展的易感性与现用抗生素相当,且相应的耐药机制高度重叠。然而,某些抗生素与细菌菌株的特定组合不易产生耐药性,揭示了可能长期有效的窄谱抗菌疗法。最后,我们提出了指导开发有效候选抗生素的标准。
结果Results
现用抗生素与在研抗生素的耐药性机制存在重叠
我们从四种革兰氏阴性细菌病原体(包括大肠杆菌、肺炎克雷伯菌、鲍曼不动杆菌和铜绿假单胞菌,详见补充表 4)中选择了40株具有代表性的菌株,并测定了它们对22种临床现用抗生素(对照组)和13种在研或2017年后引入的抗生素(新型抗生素,详见补充表 3)的体外敏感性。在这40株临床来源的菌株中,8株被确认具有广泛耐药性(extensively drug resistant, XDR),因为其对几乎所有临床推荐抗生素的最低抑菌浓度(minimum inhibitory concentrations, MIC)均超过了既定的临床断点(详见扩展数据图 2)。对于这40株菌株,与作用机制相似的对照抗生素相比,新型抗生素(如头孢吡肟、SPR-206、依拉环素和德拉沙星)平均具有显著更高的效力(即平均MIC更低,图 1a)。
实际上,基于抗生素敏感性谱的热图层次聚类分析显示,具有相似作用机制的对照抗生素和新型抗生素聚类在一起(图 1b)。此外,相较于同种细菌中的抗生素敏感菌株(antibiotic-sensitive, SEN),多重耐药(MDR)和广泛耐药(XDR)菌株对对照和新型抗生素的敏感性均普遍降低(详见扩展数据图 3a)。这些结果表明,临床使用的抗生素和在研抗生素在耐药性谱上存在重叠。然而,一些靶向细菌膜的抗生素(如 POL-7306 和 SPR-206)在对抗 MDR 和 XDR 菌株时,与对抗 SEN 菌株一样有效(详见扩展数据图 3b),突显了其抗菌潜力。
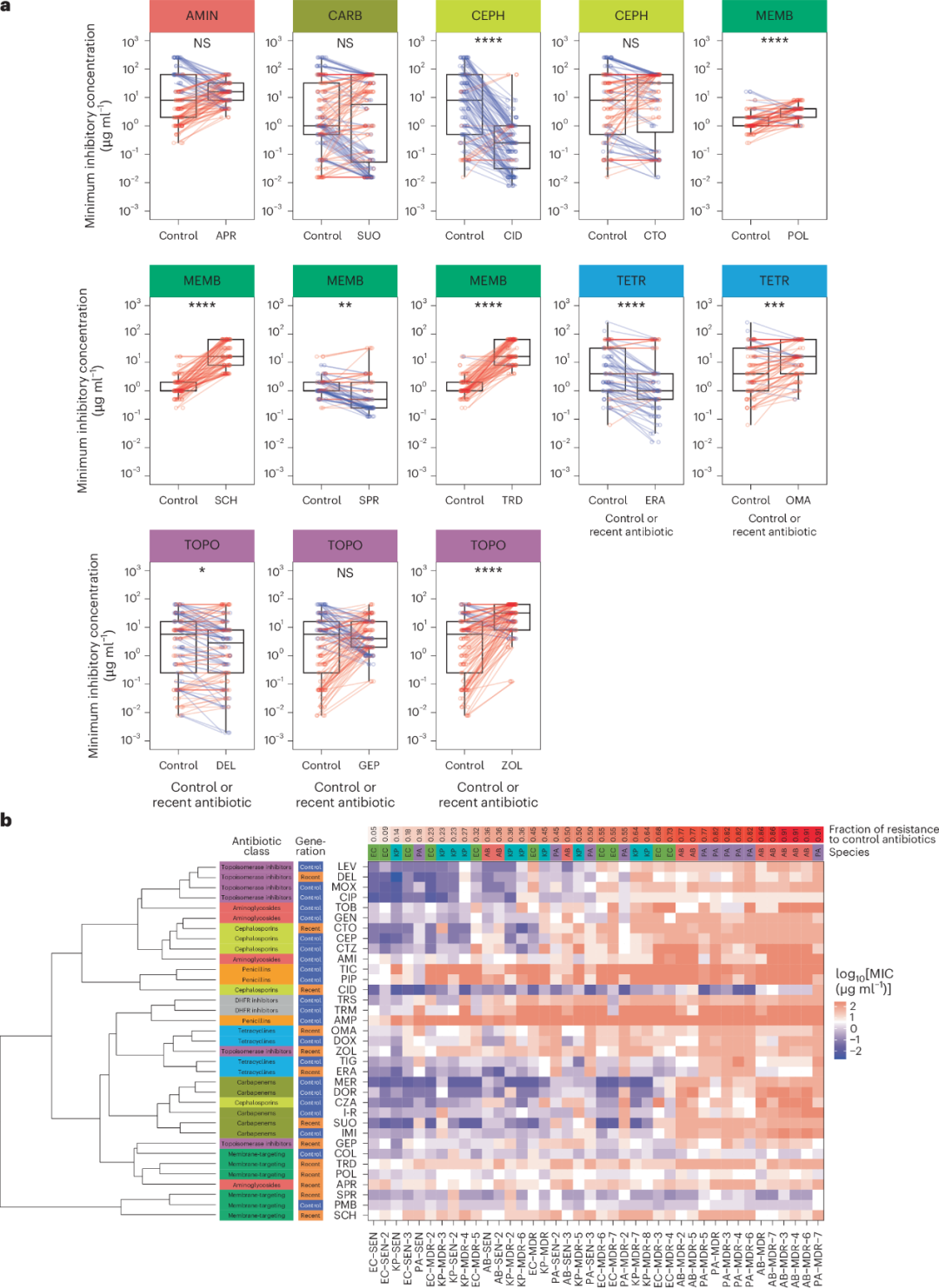
图1 | 分离菌株对对照抗生素和新型抗生素的敏感性分析
a. 对比对照抗生素和新型抗生素的MIC值。图中展示了所有测试菌株中对照抗生素和新型抗生素的MIC中位值(以log10刻度表示)。每个图代表一种特定的新型抗生素,并配有同类对照抗生素(在图顶部标注并用颜色编码)。各点表示菌株与抗生素组合的MIC中位值,连接线表示同一菌株对特定新型抗生素及同类对照抗生素的MIC值。蓝点和蓝线表示新型抗生素的MIC值低于对应对照抗生素的情况,红点和红线表示MIC值未降低的情况。MIC中位值基于每种菌株-抗生素组合的两次生物学重复和三次技术重复计算。箱线图显示中位值、第一和第三四分位数,须线表示5%和95%的分位数。通过配对Wilcoxon秩和检验(双侧检验)评估各类抗生素中对照和新型抗生素之间的显著差异。****P < 0.0001,***P < 0.001,**P < 0.01,*P < 0.05。抗生素缩写详见表1。抗生素类别包括:氨基糖苷类(AMIN)、碳青霉烯类(CARB)、头孢菌素类(CEPH)、靶向膜抗生素类(MEMB)、拓扑异构酶抑制剂(TOPO)和四环素类(TETR)。
b. 根据敏感性测试,新型抗生素与对照抗生素聚类在一起。热图展示了4种细菌(E. coli、K. pneumoniae、A. baumannii 和 P. aeruginosa)菌株的抗生素敏感性谱(列)。在x轴标签中,前两位字母表示物种(AB:鲍曼不动杆菌;EC:大肠杆菌;KP:肺炎克雷伯菌;PA:铜绿假单胞菌),接下来的三位字母指示根据敏感性分析的菌株分类(见方法),最后的数字为唯一标识符(注意,仅使用无数字标识的菌株进行进一步实验)。细菌菌株按照对照抗生素耐药比例(顶部红色渐变条)排列,耐药性依据相应物种特异的临床断点值定义。有关缩写的详细信息,抗生素见表1,物种和菌株见补充表4。抗生素面板包括22种对照抗生素(蓝色)和13种新型抗生素(橙色)(行)。抗生素的代次和类别标注在左侧。抗生素聚类基于计算中位MIC值的Spearman秩相关,并采用完全层次聚类法。细菌菌株按照对照抗生素耐药比例排序(顶部红色渐变条)。MIC中位值基于每种菌株-抗生素组合的两次生物学重复和三次技术重复。
抗生素缩写包括:AMI(阿米卡星)、AMP(氨苄西林)、APR(硫酸阿普拉霉素)、CEP(头孢吡肟)、CID(头孢吡罗)、CIP(环丙沙星)、COL(多粘菌素)、CTO(头孢他啶-阿维巴坦)、CTZ(头孢他啶)、CZA(头孢他啶-阿维巴坦)、DEL(德拉沙星)、DHFR(二氢叶酸还原酶抑制剂)、DOR(多尼培南)、DOX(多西环素)、ERA(依拉环素)、GEN(庆大霉素)、GEP(吉波霉素)、IMI(亚胺培南)、I-R(亚胺培南-雷立酶)、LEV(左氧氟沙星)、MER(美罗培南)、MOX(莫西沙星)、OMA(奥玛环素)、PIP(哌拉西林)、PMB(多粘菌素B)、POL(POL-7306)、SCH(SCH-79797)、SPR(SPR-206)、SUO(舒洛培南)、TIC(替卡西林)、TIG(替加环素)、TOB(妥布霉素)、TRD(三十三肽)、TRM(甲氧苄啶)、TRS(甲氧苄啶-磺胺甲噁唑)、ZOL(左氧氟沙星)。
实验室中耐药性物种特异性演变
接下来,我们研究了抗生素耐药在细菌病原体中的演化过程,这是导致两组抗生素在长期内疗效的下降的原因。我们选择了每种细菌(如 E. coli、K. pneumoniae、A. baumannii 和 P. aeruginosa)中的一个多重耐药(MDR)菌株和一个敏感菌株(SEN),详见补充表 4。在所有抗生素-菌株组合中,由于初始抗生素敏感性较低(即 MIC > 4 µg/ml),32% 的菌株被排除在分析之外。
为了研究耐药的初步阶段,我们采用了标准实验方案进行自发耐药频率(Frequency of Resistance, FoR)分析,并使用多种抗生素浓度进行测试。约 1010个细菌细胞在培养基板上暴露于每种抗生素中,持续 2 天,浓度足够使给定菌株敏感。检测到耐药突变体(即 MIC 值至少增加 4 倍的菌株)在 49.8% 的菌株中出现。尽管大多数新型抗生素的临床断点尚未明确,但所有抗生素均提供了推荐的静脉用药剂量。因此,我们用最高可用血浆浓度(通过静脉给药时测量)作为代理,来估计耐药菌株中 MIC 变化的潜在临床意义(详见补充表 3)。在短短 48 小时内,MIC 值要么等于,要么高于峰值血浆浓度,其中最多 18.7% 的突变体株(扩展数据图 4a)。在 30% 的 FoR 适应株中,MIC 值超过了临床断点,其中数据可用(扩展数据图 4b)。在所有菌株中,新型抗生素和对照抗生素对细菌耐药同样容易,因为突变体每代出现的频率(Wilcoxon秩和检验,P = 0.9)和耐药性折合值的变化在统计上没有显著差异(配对 t 检验,P = 0.68)。
由于 FoR 检测无法单独或协同检测极为罕见的突变,并可能低估细菌耐药性的潜力,因此,我们使用了相同的八种原始菌株(详见补充表 4)来启动适应性实验室进化(adaptive laboratory evolution, ALE),目标有两个。首先,我们旨在最大化在固定时间内(最长可达 ∼120 代)群体中获得的抗生素耐药水平。其次,我们旨在明确与耐药性相关的机制。每种菌株的 10 个平行进化群体暴露于逐渐增加浓度的新型或对照抗生素中。通过比较进化株与其相应原始菌株的 MIC 值(图 2a),我们估计了耐药性的水平。
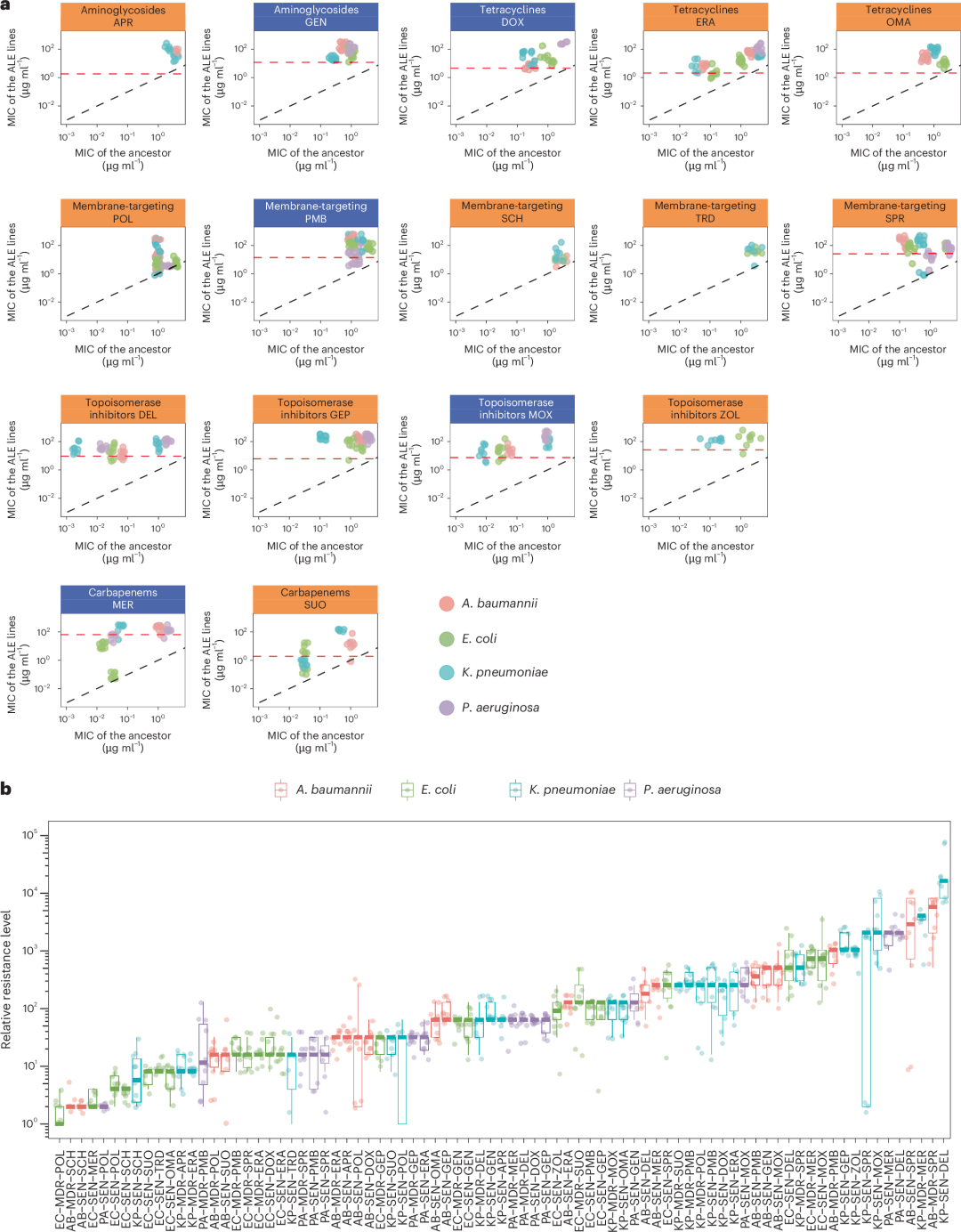
图2 | 通过ALE的抗生素适应
a、ALE后MIC值的变化。每个点代表一个实验演化菌系的中位MIC值和相应的祖先(以对数10为刻度)。中位MIC值基于每种细菌菌系–抗生素组合的两个生物学和三个技术重复。对照抗生素用蓝色绘制,新型抗生素用橙色绘制。每组图表示一个特定的抗生素类别(如图顶部所示)。数据点的颜色表示细菌种属。黑色虚线表示y = x(即实验演化过程中MIC值未变化),红色虚线表示特定抗生素的峰值血浆浓度。由于实验液体培养基的不稳定,头孢类抗生素未接受ALE处理。b、实验演化菌系在所有抗生素–祖先菌系组合中的相对MIC。x轴标签中的前两个字母代表种属,接下来的三个字母代表菌系,最后三个字母代表抗生素。有关缩写的详细信息,请参见表1(抗生素)和补充表4(种属和菌系)。相对MIC是演化菌系的中位MIC值除以相应祖先的中位MIC值。每个点是来自ALE的实验演化菌系,颜色表示细菌种属。每个点是菌系–抗生素组合的中位MIC值。中位MIC值基于每种细菌菌系–抗生素组合的两个生物学和三个技术重复。箱图显示了中位数、第一和第三四分位数, whiskers表示MIC值的5%和95%分位数。在抗生素–菌系组合中的相对MIC存在显著的异质性(Kruskal–Wallis χ2 = 630.43,d.f. = 80,P < 2.2 × 10−16)。
总体而言,经过120代(60天)的实验室进化,细菌菌株已足以产生抗药性;进化菌株的抗生素耐药性水平中位数比祖先菌株高约64倍。在所有研究的群体中,87%的最小抑菌浓度(MIC)等于或高于抗生素的血浆峰值浓度。此外,在有临床断点数据的情况下,88.3%的自适应实验室进化(ALE)菌株的MIC值超过了临床断点(扩展数据图4b)。平均来看,新型抗生素和对照抗生素在细菌抗药性方面同样易感(配对t检验,单侧,P = 0.37)。耐药性也出现在针对最近开发的抗生素上(图2a),这些抗生素对测试的多重耐药(MDR)和广泛耐药(XDR)临床分离株具有强效的抗菌活性(扩展数据图3b)。
鉴于抗生素与菌株组合在耐药性进化能力上表现出高度异质性,MIC倍增变化的最小值和最大值相差高达65,000倍(图2b),我们探讨了这一差异的可能原因。首先,我们分析了初始抗生素敏感性是否能够预测细菌对药物的长期反应。结果表明,在四个物种中的两个(扩展数据图5a)以及研究的八个菌株中的五个(扩展数据图5b)中,初始MIC与耐药水平的增加呈显著正相关。接着,我们逐一分析了每种抗生素的情况,评估初始MIC是否与菌株间耐药水平的增加有关。结果显示,在16种抗生素中,有5种表现出显著的正相关趋势(扩展数据图5c)。这些发现表明,初始MIC可以以菌株和抗生素特异性的方式预测抗生素的长期疗效。
先前的研究表明,某些抗生素在特定细菌菌株和物种中比其他抗生素更容易进化出耐药性。因此,我们使用多元线性回归分析了抗生素和菌株遗传背景对耐药性水平增加的整体影响,同时也考虑了祖先菌株的耐药水平(扩展数据图5d)。分别分析这些因素时,抗生素和菌株遗传背景分别解释了耐药性水平增加变异的24.4%和8.9%,初始抗生素敏感性水平(MIC)贡献了0.9%的变异。在加性模型中,将抗生素、初始MIC和遗传背景结合起来能够解释约33%的变异。值得注意的是,一个允许遗传背景与抗生素结合交互作用的模型相比简单加性模型,额外解释了约26%的耐药性水平增加变异(即58.6%对比32.6%;扩展数据图5d)。
综上所述,这些结果表明,细菌种群的初始遗传组成对耐药性进化有显著影响,但主要是以抗生素特异性的方式体现。对两种抗生素候选物SCH-79797和SPR-206的详细分析进一步支持了这一点(补充说明2)。
抗生素处理之间突变谱的重叠
为识别与耐药性相关的突变,我们对实验室进化获得的耐药菌株(n = 381)和FoR实验中的菌株(n = 135)进行了全基因组测序(补充数据1)。通过既定的计算流程,我们识别了相对于祖先基因组的突变。结果显示,有10株进化菌株积累了大量异常的突变(n > 18),其中许多可能在功能上无关。这些菌株表现出较高的基因组突变率;实际上,其中6株在甲基化依赖的错配修复基因(mutS、mutL或mutY)中存在突变。这类突变体常在抗生素应激下于临床或实验室环境中产生。
在其余的506株菌株中,我们共识别出1,817个独特的突变事件,包括1,212个单核苷酸多态性(SNP)和605个插入或缺失(扩展数据图6a)。非同义突变显著多于同义突变,表明蛋白编码区的SNP积累主要受选择压力驱动,从而提高耐药性(扩展数据图6b)。此外,观察到的突变中有19.7%生成了框内终止密码子、移码突变或起始密码子的破坏,这些可能是功能丧失型突变(扩展数据图6c)。这一结果与先前研究中关于失活突变在抗生素耐药性中作用的发现一致。
共检测到604个发生突变的蛋白编码基因,其中193个基因在每个物种中至少有两个独立进化的菌株中出现突变。在所有平行突变的基因中,69.4%在适应不同抗生素的菌株中携带突变。这些结果表明,尽管抗生素处理方式不同,突变基因集合之间仍存在显著的重叠(图3a,b)。进一步的结果显示,在抗生素存在下进行的实验室进化过程中,适应性变化与培养基的关系不大(补充说明3)。
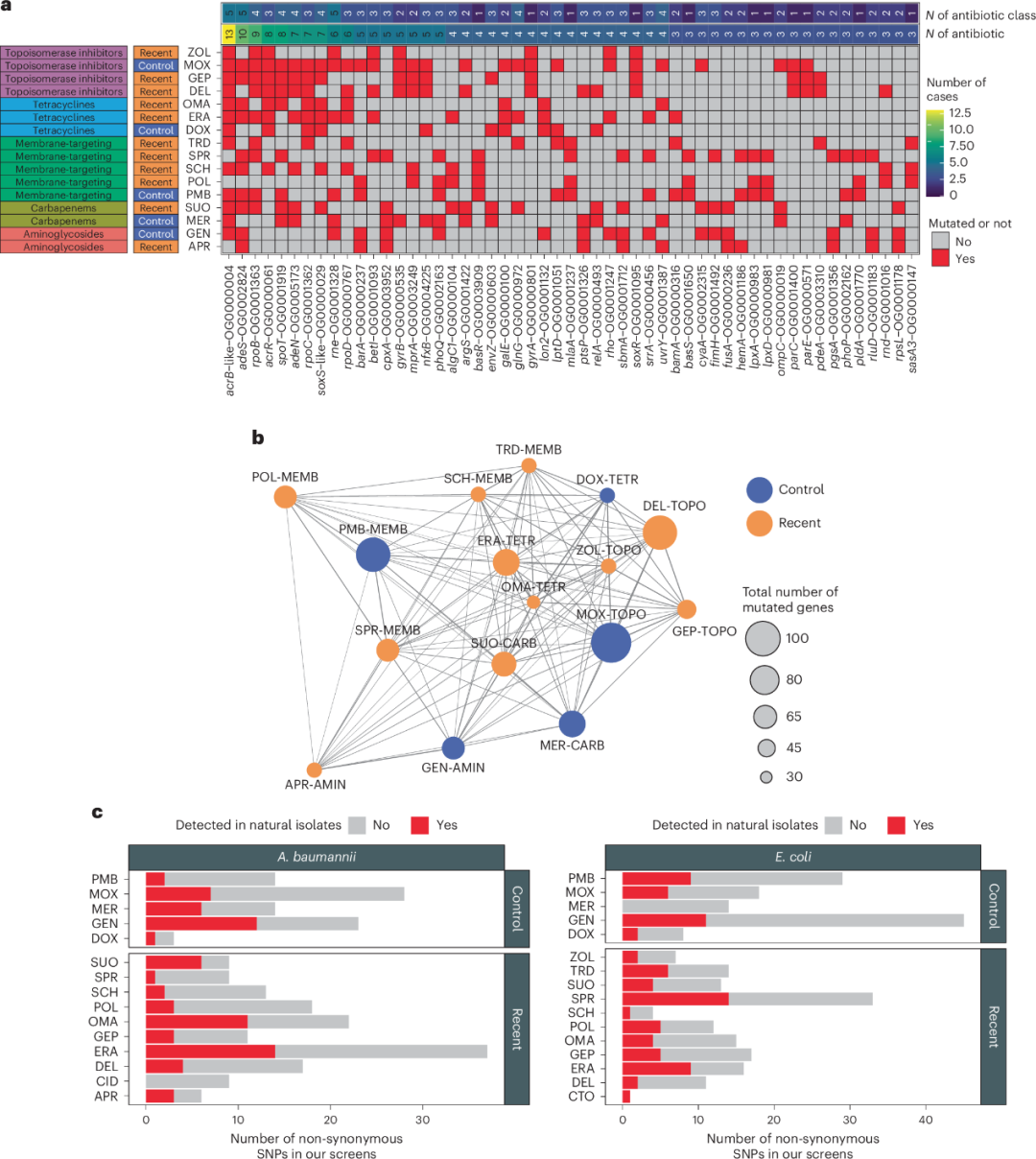
图3 | 利用全基因组测序对抗生素适应菌株的基因组进行分析
a. 实验室进化过程中在不同抗生素条件下反复发生突变的基因。热图展示了自适应实验室进化(ALE)后在测试抗生素中常见的突变基因(及其对应的直系同源组)。若某基因对至少三种不同的抗生素处理产生非同义突变,则被认为是常见突变基因。左侧列出了所用抗生素的类别及其代际信息。热图顶部两行分别表示突变发生的抗生素类别数量和直系同源组在突变中涉及的抗生素数量。b. 不同抗生素间的突变谱相似性。每个节点代表一种新型(橙色)或对照(蓝色)抗生素。连接线表示ALE后检测到的突变基因(或相应的直系同源组)集合之间的重叠。仅考虑蛋白编码基因中的非同义突变。连接线的粗细表示抗生素处理间突变谱重叠的程度(用Jaccard相似性计算,参考先前研究)。c. 实验室进化菌株与自然分离菌株中共享的非同义突变。柱状图显示了在适应不同抗生素的实验室进化鲍曼不动杆菌(左)或大肠杆菌(右)中发现的非同义突变数量。在同一物种的自然分离株基因组中也检测到的突变以红色标注,而未检测到的突变以灰色标注。在对照抗生素和新型抗生素之间,自然菌株共享的非同义突变比例没有显著差异(双侧二项回归模型,P = 0.206)。有关缩写,请参见表1。
为了明确交叉耐药性,我们专注于拓扑异构酶抑制剂,因为该类药物包含了目前正在临床试验中的大量抗生素,且耐药性通常由耐药突变编码。我们在编码莫西沙星靶点的基因(gyrA和parC)中进行了深度突变扫描研究。导致莫西沙星耐药的突变组合显著降低了临床开发中的拓扑异构酶抑制剂(包括迪拉弗洛沙星(delafloxaci)和吉珀替嗪(gepotidacin))的敏感性(补充说明4)。与吉珀替嗪的结果令人意外,因为它是一种正在开发的新型拓扑异构酶抑制剂,具有创新的靶点和作用机制。先前的研究报道,氟喹诺酮耐药临床分离株对这种抗生素没有交叉耐药,但研究数据有限。
新型抗生素的突变在环境中存在
鉴于与临床使用的抗生素和开发中的抗生素相关的突变靶点重叠(图3a,b),我们假设,实验室进化过程中产生的突变可能已经存在于环境中以及临床分离的细菌中。为了检验这一假设,我们分析了实验室进化获得的大肠杆菌和鲍曼不动杆菌菌株在公开可用的基因组目录中的流行程度,该目录包含了来自大肠杆菌(n = 20,786)和鲍曼不动杆菌(n = 15,185)的自然分离株基因组(图3c)。我们集中分析了编码蛋白的非同义突变,并估算了这两种细菌环境分离株基因组中这些突变的频率。在大肠杆菌中,观察到的245个非同义突变中,最多31.4%出现在至少1个自然分离株的基因组中;对于鲍曼不动杆菌,216个突变中,27.3%也出现在至少1个自然分离株中。尽管大肠杆菌中的大多数突变相对稀少(通常在1%以下的分离株中找到),但它们在病原性分离株中的丰度高于其他自然分离株(Fisher’s检验,P < 2.2 × 10−16,OR = 3.16)。在临床分离株中,一些适应性突变的丰度与典型的抗药突变相当,甚至高于抗生素临床分离株中的抗药突变(扩展数据图7)。
靶向新型抗生素的可移动耐药基因种类繁多
ALE无法分析水平转移的耐药机制。因此,我们接下来分析了环境样本和临床样本中可移动耐药基因(ARGs)的丰度。我们之前从以下样本创建了宏基因组文库:(1)人类活动影响的土壤微生物群落,例如印度7个抗生素污染工业场地附近的河流沉积物和土壤样本;(2)人体肠道微生物群落,例如来自欧洲10位未使用抗生素超过1年的个体的粪便样本;(3)临床微生物群落样本,这些样本来自于68株耐多药(MDR)菌株,分别从医疗设施中分离或从菌株收集中获得(补充表5)。每个文库包含多达500万DNA片段(contig),总覆盖面积为25 Gb(即约5,000个细菌基因组的大小)。我们使用了已建立的功能宏基因组实验方案,检测这些文库中长约1.7 kb的小DNA片段,这些片段在本质上对临床大肠杆菌和肺炎克雷伯菌株的敏感性高,且可转移耐药性。具体来说,这些DNA片段在这两种菌株中异源表达。
总共690个独立的DNA片段赋予了细菌宿主对测试的新型和对照抗生素最高可达256倍的耐药性(补充数据2)。总体来看,在研发中的抗生素与其对应的内类对照之间,赋予耐药性的contig数量没有显著差异(配对Wilcoxon符号秩检验,P = 0.791;图4a)。然而,我们在任何宏基因组文库和宿主种属中均未检测到与三十烯M152-P3耐药相关的DNA片段(图4a)。临床微生物群体贡献了57.8%的抗生素耐药性DNA片段,显著高于土壤和肠道微生物群体的贡献(分别为25.5%和24.8%)。总共检测到642个非冗余开放阅读框(open reading frames, ORF),其中许多片段出现在多个DNA片段中(补充数据2)。在690个DNA片段中,77%显示与相关数据库中的已知耐药基因(即ARGs)具有相似的序列(补充数据2)。这些ARGs涉及抗生素失活、抗生素外排或保护抗生素靶点等,其系统发育起源十分多样(补充数据2)。
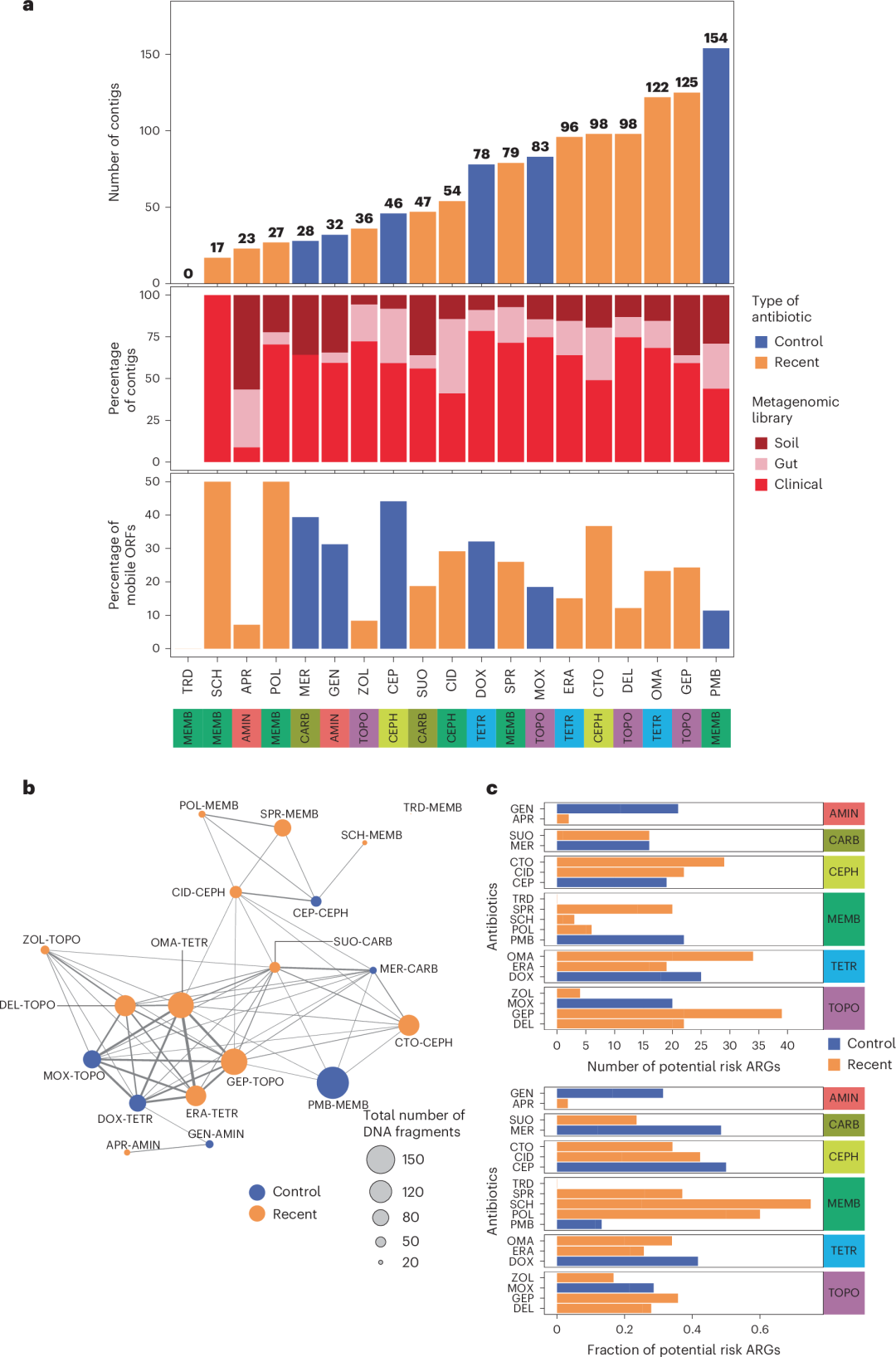
图4 | 利用功能宏基因组学探索可移动耐药基因
a. 概述功能宏基因组学生成的抗生素耐药菌株的测序结果。通过18种抗生素对宏基因组文库进行功能筛选,生成了许多不同的耐药性编码DNA片段,仅在三十烯M152-P3(tridecaptin, TRD)中未检测到。顶部条形图显示了赋予对照(蓝色)和新型(橙色)抗生素耐药的独立DNA片段(contig)的数量,而底部条形图展示了在宏基因组文库中检测到的耐药性编码contig的分布以及每种抗生素的移动开放阅读框(ORF)百分比。ORF的移动性由近期的水平基因转移事件或存在于可移动质粒上的证据定义(方法部分)。条形图下方的行标明了分析的抗生素类别。我们观察到在最近抗生素和其对应内类对照之间,contig数量无显著差异(配对Wilcoxon符号秩检验,双侧,P = 0.4973)。相同的模式也适用于移动ORF的百分比(配对Wilcoxon符号秩检验,P = 0.576)。关于抗生素的缩写,请参见表1。
b. 不同抗生素中耐药性编码DNA片段(contig)的重叠。每个节点代表最近(橙色)或对照(蓝色)抗生素,链接表示在功能宏基因组筛选中识别的耐药性编码DNA片段的重叠。链接的厚度表示抗生素处理之间重叠程度(由Jaccard相似性计算)。节点的大小对应于每种抗生素检测到的DNA片段总数。给定抗生素所属的类别也指示出来。
c. 潜在ARGs的风险分析。图表显示了在不同抗生素功能筛选中健康风险ARGs的总数(顶部)和比例(底部)。如果ARGs满足以下三个标准中的至少两个,它们被归为潜在风险:1)移动,2)存在于与人类相关的微生物群中,3)为人类病原菌。蓝色和橙色表示对照和最近的抗生素。分析揭示了在测试的抗生素中潜在风险ORF的比例存在显著差异(比例检验,双侧,P < 0.05),这表明某些抗生素更可能与潜在风险ORF相关。
与基因组突变和ARGs相关的潜在耐药机制在性质上存在显著差异(扩展数据图8)。特别是,抗生素外排和靶标改变是由基因组突变引发的最普遍的耐药机制(扩展数据图9a),而抗生素失活则在功能基因组筛选中的检出率更高(扩展数据图9b)。有关基因组突变和可移动耐药基因的详细比较,请参见补充说明5。此外,我们的分析还揭示了非典型耐药机制的贡献(补充说明6)。
编码多重耐药(MDR)DNA片段的富集程度
我们观察到,赋予对照和新型抗生素耐药的DNA片段之间存在重叠(图4b),包括具有不同作用机制的抗生素对。例如,检测到69个contig能够耐受拓扑异构酶抑制剂莫西沙星和四环素抗生素奥马沙星。在这些contig中,分析识别出了两个关键基因,baeR和ramA,它们分别由73.9%的contig携带。BaeR和RamA分别诱导MdtABC/AcrD28和AcrAB–TolC外排泵复合体的表达,RamA还下调了外膜蛋白OmpF的表达。这表明,一些DNA片段能够耐受多种抗生素,这可能反映了化学结构或作用机制的相似性。因此,我们通过使用SMILES(简化分子输入线条记录系统)标识符定量抗生素对的结构相似性,研究化学相似性对共耐药性的影响,并分析它们与基于检测DNA片段的轮廓相似性的相关性。我们发现,赋予耐药性DNA片段的重叠更可能发生在化学结构相似的抗生素对之间(Spearman’s秩相关,双侧检验,R = 0.43,P < 0.01)。当仅考虑属于不同抗生素类的抗生素对时,这一模式依然成立(Spearman’s秩相关,双侧检验,R = 0.31,P < 0.01)。然而,这般模式也存在显著偏离。例如,尽管SPR-206是多粘菌素B的衍生物,但没有DNA片段能够赋予两者耐药。同样,尽管cefiderocol与其他研究的头孢菌素结构相似,但氯钴香豆素的加入使其成为一个络合物,赋予耐药性的DNA片段大多特异于这一抗生素。总之,这些分析表明,化学结构的某些改变影响了抗生素的作用机制或摄取方式,从而导致关联耐药机制的重大变化。
耐药基因的健康风险分析
接下来,我们使用了一个此前开发的基于组学的方法框架来评估ARGs的健康风险。我们考虑了三个主要的ARG标准:(1) 基因的迁移性, (2) 存在于与人体相关的微生物群落中,和 (3) 病原性细菌的宿主。ARGs被定义为“潜在风险”,如果它们至少满足三项标准中的两项。
通过使用已建立的方法,基因的迁移性是通过在自然界中最近的水平基因转移事件的证据以及来自多种环境的自然质粒上的ARGs来定义的(方法部分)。在潜在ARG中,20.7%的基因被发现是通过质粒携带的,或经历了最近的水平基因转移事件(图4a,扩展数据图10a和补充数据2)。接下来,我们分析了人体微生物群中潜在ARG的丰度。我们在非冗余全球微生物基因目录(GMGCv1)中识别出了我们功能性组学筛选检测到的ARG的近亲。该目录总结了来自14个主要栖息地,包括人体微生物群、家养动物、废水、淡水和人工环境的超过13,000个可公开获取的组学数据结果。
大多数微生物基因在目录中是特定于单一栖息地的。相反,27.6%的潜在ARGs在耐药性筛选中检测到的抗微生物剂中出现在多个栖息地,这进一步表明它们可能具有迁移性。此外,当仅考虑与人体相关的栖息地(人体肠道、口腔、皮肤、鼻腔、血浆或阴道微生物群)时,这一比例上升至32.7%,表明这些微生物群落可能是新型抗生素耐药基因的丰富来源(扩展数据图10b)。令人欣慰的是,与人体相关的ARGs在与人体相关的非生物栖息地(如废水或人工环境)中也比其他ARGs更为普遍(Fisher’s检验,OR = 125,P < 0.0001)。在ARGs中,36.6%已经存在于临床重要的细菌病原体基因组中(扩展数据图10b,方法部分和补充数据2)。检测到的ARGs在从三个主要栖息地(农业、人体或野生动物宿主)分离出的大肠杆菌菌株基因组中也很常见(补充说明7)。
在642个ARGs中,24.5%被指定为潜在风险(补充数据2)。这些ARGs预计通过结合其危险特征,包括迁移性导致的广泛宿主兼容性,以及在人体微生物群落和细菌病原体中的富集,具有显著催化多重耐药性的潜力。然而,不同抗生素中潜在风险ARGs的频率存在显著差异(图4c)。一个典型的例子是阿帕霉素硫酸盐,这是一种在兽医学中广泛使用的抗生素,目前正处于用于人体应用的临床试验阶段。与该抗生素相关的63个潜在ARGs中,仅2个(3.2%)被指定为潜在风险,原因是缺乏其在细菌病原体中的迁移性和存在证据。
相反,对于一些新型抗生素,如磺苯类(N = 16)、头孢地尔(cefiderocol)(N = 22)和广谱头孢菌素(ceftobiprole)(N = 26),我们检测到了多种潜在风险ARGs。这些潜在风险ARGs包括多种β-内酰胺酶,如新德里金属酶(NDM)和维罗纳整合子金属β-内酰胺酶(扩展数据图10b)。尽管此前对头孢地尔耐药性发展较低的预期,这种抗生素中潜在风险ARGs的数量仍然显著。
整合对新型抗生素候选药物耐药性的证据
理想的抗生素候选药物应具备以下几个关键特性:(1) 广泛的抗菌活性,确保对多种病原体具有显著的效果;(2) 在基因组突变导致耐药性方面,具有较低的风险;(3) 内源性和水平转移的可移动抗药基因(ARGs)稀少;(4) 与人体微生物群和细菌病原体中的耐药机制相关的现象较少。然而,研究中涉及的所有化合物都未能同时满足这些标准(图5)。通过综合多项收集的数据,我们得出了一个平均指标值,用于根据其耐药特征对新型抗生素候选药物进行排名,这在不同抗生素类之间显著存在异质性(Kruskal–Wallis检验,P < 0.05;图5)。根据这一排名,与四环素和拓扑异构酶抑制剂相比,针对细菌膜的最新抗生素在自然环境中的耐药性开发可能会显著减弱(Dunn后测检验,经过Benjamini–Hochberg校正的多重比较,P < 0.05)。然而,这些药物在疗效方面仍有很大的提升空间。
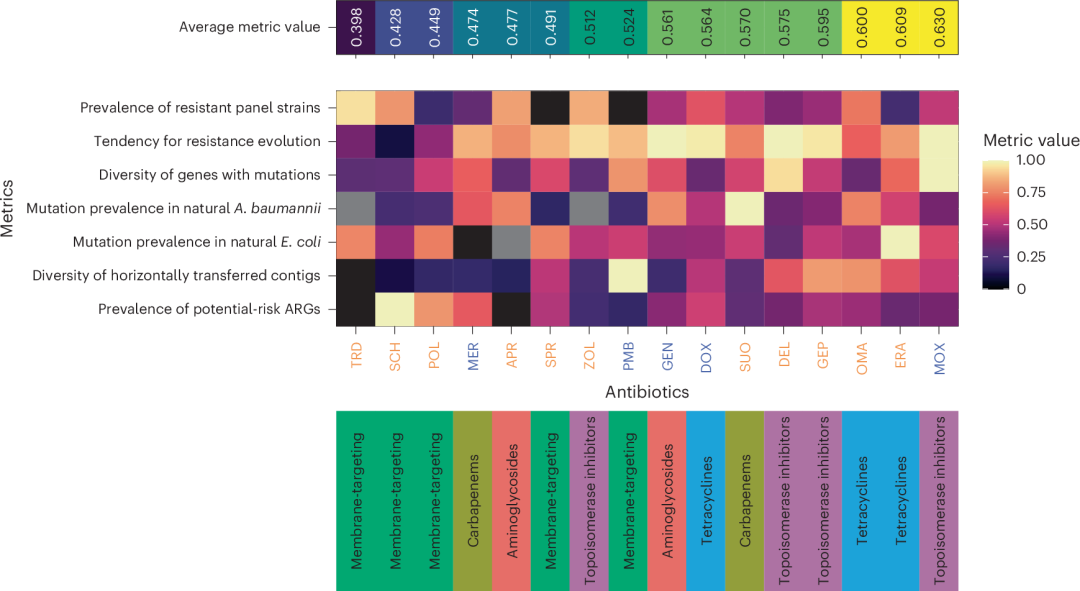
图5 | 每种抗生素的耐药性概览
该热图展示了该研究中每种抗生素的各种指标。指标包括:(1)降低敏感性菌株的流行程度——来自选定病原菌面板中具有高初始MIC值的细菌菌株的比例(方法);(2)耐药性的趋势——适应菌株中MIC相对值超过16的比例,代表所有相对MIC值的25%分位数;(3)突变基因的多样性——在ALE或FoR实验中,每种抗生素期间突变的Orthogroups的比例,经调整后为总的突变Orthogroups数量;(4)和(5)天然大肠杆菌或A. baumannii菌株突变的流行程度——实验观察到的适应性突变在天然大肠杆菌和A. baumannii菌株中已经存在的比例;(6)水平转移contig的多样性——每种抗生素的唯一DNA片段数,标准化为功能性宏基因组研究中的总contig数量;以及(7)潜在风险ARGs的流行程度——根据符合至少两项健康风险标准(方法)的ARGs比例,作为每种抗生素检测到的所有ARGs中潜在风险ARGs的比率。灰色表示由于研究种属的初始耐药性而缺失的值。抗生素按平均指标值排序。蓝色和橙色分别表示对照和新近使用的抗生素。热图下方的行表示所分析抗生素的类别。
作者简介
匈牙利塞格德国家生物技术实验室 Lejla Daruka, Márton Simon Czikkely, Petra Szili 和 Zoltán Farkas 为本文的共同第一作者,匈牙利塞格德生物研究中心(BRC)生物学家 Csaba Pál 为本文的通讯作者。
Csaba Pál (通讯作者)

Csaba Pál, 欧洲科学院院士, 匈牙利科学院通讯院士,匈牙利塞格德生物研究中心(BRC)生物学家。他的实验室是BRC合成和系统生物学部门的一部分。他的研究处于进化、抗生素耐药性和基因组工程的界面,在这些领域发表了多篇科学论文。Csaba Pal 致力于基因组网络进化和抗生素耐药性的基础和应用问题。为了实现这些目标,他开发了系统生物学、计算代谢建模和基因组工程的方法。
翻译:荀佳妮,中国农科院基因组所,生物信息学硕士在读
审核:朱志豪,广东医科大学,基因组所联合博士后
终审:刘永鑫,中国农科院基因组所,研究员/博导
排版:杨海飞,青岛农业大学,基因组所联培硕士在读
宏基因组推荐
本公众号现全面开放投稿,希望文章作者讲出自己的科研故事,分享论文的精华与亮点。投稿请联系小编(微信号:yongxinliu 或 meta-genomics)
猜你喜欢
iMeta高引文章 fastp 复杂热图 ggtree 绘图imageGP 网络iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文

















 693
693

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









