






摘要
工作记忆是一个涉及大脑中多个功能解剖节点的关键认知过程。尽管有大量与工作记忆结构相关的神经影像学证据,但我们对控制整体表现的关键中枢的理解并不完整。因果解释需要在对特定功能解剖节点进行安全、暂时和可控的神经调节后进行认知测试。随着经颅交流电刺激(tACS)技术的进步,此类实验可以在健康人群中进行。本研究结合了28项安慰剂对照研究(共1057名参与者)的结果,这些研究应用了特定频率的非侵入性神经振荡刺激,并检查了神经正常成年人的工作记忆表现。本研究使用计算元建模方法来模拟虚拟现实大脑中的每次干预,并将观察到的行为结果与不同大脑节点中的刺激诱发电场进行比较测试。本研究结果表明,刺激前额叶和内侧颞叶θ振荡以及枕顶叶γ节律可以显著改善工作记忆任务表现。相反,前额叶γ调制不利于表现。此外,本研究发现了θ亚频段的空间表达存在差异,其中工作记忆变化遵循眶额高θ调制和内侧颞叶低θ调制。最后,所有这些结果都是由工作记忆准确性的变化而不是处理时间测量驱动的。这些发现为工作记忆机制提供了新的视角,与神经影像学研究相辅相成,并提出了临床治疗工作记忆缺陷的假设驱动目标。
引言
工作记忆(WM)是一种主动保持和操纵目标相关信息的复杂认知机制。数十年的脑成像研究已将人类WM与多个神经解剖中枢(包括额叶、顶叶、颞叶和枕叶区域)关联起来。通常情况下,前额叶皮层执行自上而下的控制以及记忆痕迹的短期保持和操作。内侧颞叶皮层在WM绑定、关联以及工作记忆和长期记忆之间的动态关系中发挥作用。顶上小叶和顶下小叶负责WM任务中记忆痕迹的时间操作,并与枕叶皮层合作,枕叶皮层也负责编码视觉记忆项目。从神经功能的角度来看,大脑区域表现出多种引起大脑振荡的宏观电活动模式。振荡或节律源于神经元群的突触通讯和相互协调。它们作为功能合作的载体存在于局部环路和远程网络中。研究通常将额叶和颞叶θ节律(3-8Hz)、后部α节律(8-13Hz)和各种γ节律(30-100Hz)与领域一般性WM过程联系起来。此外,给定的WM范式可以促进刺激或任务特异性节律,例如感觉运动β振荡(13-30Hz)。
经典的WM研究包括将计算机任务操作与人类或非人类灵长类动物的神经成像或电生理记录相关联。尽管这些研究非常重要,但它们往往难以确定导致WM处理出现问题的确切原因。对脑病变患者进行的认知研究为神经解剖学提供了更具因果关系的视角。然而,病变影响很少局限于单个完整的解剖单元内,并且几乎不能提供有关特定脑振荡的功能信息。
认知研究中非侵入性神经调节(尤其是经颅交流电刺激(tACS))技术的进步为健康人类的功能神经解剖学提供了独特的视角。在tACS期间,通过头皮向大脑施加特定频率的振荡电流,产生空间受限的电场,该电场可以使特定频率的内源性脑振荡同步。与神经影像学研究不同的是,tACS实验可以揭示一个选定的功能区域,其调节能够引起测量结果发生变化。因此,神经调节与神经影像学相辅相成,以实现对大脑的全面了解。大量研究测试了刺激大脑振荡对WM任务的因果影响,结果显示由于功能相关区域和节律的同步而导致WM任务的表现得到改善。另外,不兼容参数下的tACS调节可能会破坏行为效率。
最近的几项元分析总结了tACS对工作记忆或更广义认知表现的影响,并发现了中等大小的显著刺激效应。此外,之前的元分析显示tACS在正常成年人、老年人和有认知功能缺陷的临床人群中具有类似的效果。然而,传统的元分析根据声明的刺激参数(例如电极位置或总电流强度)考虑实验研究,而没有考虑tACS在靶区和非靶区位置上的表现。计算建模可以考虑tACS生物物理学并提供新的见解。也就是说,我们可以模拟任何tACS干预,并精确估计大脑中任何点的感应电场,从而揭示每个解剖体素所施加的“剂量”。因此,我们可以评估区域功能变化与行为结果之间的因果关系。
在这里,本研究使用tACS研究的元建模综合了领域一般性WM的因果功能图。元建模利用已发表实验的大规模计算模拟来建立WM表现与脑区振荡调制之间的关系。本研究确定了28项在解剖学正常成人中评估WM的安慰剂对照tACS研究,其中报告了64种干预类型,包括各种解剖靶区、刺激强度和振荡频率。本研究在100个神经正常成人大脑中对每次干预进行了模拟,共计6400个计算模型,将它们投影到每个频段的共同大脑空间中,并根据报告的行为结果(如WM 准确性和加工时间)分析诱导电场。这些功能图确定了每种振荡节律下调节领域一般性WM最敏感的神经解剖区域。
材料与方法
研究样本。两位专家独立地对PubMed和Web of Science核心合集进行广泛检索后,确定并收集了28项研究(总共1057名参与者)的数据,这些研究应用tACS和安慰剂/伪刺激(安慰剂对照设计)调节WM表现。这28项研究采用了64种主动干预类型,使用不同的刺激强度、刺激电极位置和刺激频率。每项研究调查一个或多个结果(反应时和/或准确性、刺激时间、任务难度水平),总共产生了298个结果测量值。
效应量估计。本研究计算了以Hedges’g表示的安慰剂对照标准化效应大小。为了获得该测量值,首先计算伪刺激(安慰剂)和主动tACS之间的平均差异。也就是说,每个结果测量值(准确性、RT等)都从主动tACS期间或之后的分数中减去伪tACS期间或之后的分数(scoreactive-scoresham)。如果研究报告了基线测量值,则通过从伪tACS或主动tACS期间或之后的表现中减去该值来进行基线控制[(scoreactive-scoreactive_baseline)-(scoresham-scoresham_baseline)]。随后,将安慰剂对照和基线安慰剂对照的平均分除以总SD,得到Cohen’s d标准化效应量估计。Hedges’g是Cohen’s d的一种变体,但具有样本量偏差校正,适用于小样本量(n<20)研究。在本研究的分析中,Hedges’g大于0表示行为表现得到改善,例如更高的准确性或更低的反应时间,而小于0则表明行为表现较差。这些效应量估计用于元建模。
头部模型和电场建模。本研究在100名健康成年人(22-35岁,50名女性)的大脑中模拟每种干预类型的诱导电场,以代表典型的tACS研究人群。这些成年人的大脑图像(T1加权和T2加权结构MRI)来自人类连接组计划(HCP)数据库。使用SimNIBS 3.2框架(pipeline headreco)生成个体真实头部模型以进行计算模拟。该框架使用SPM12和CAT12执行自动组织分割和表面重建。随后由人体解剖学专家对这些重建进行手动检查和校正。使用Gmsh将所得数据自适应地划分为由6个不同组织类型组成的有限元(FEM)模型。这些组织类型包括皮肤、颅骨、脑脊液、眼睛、灰质和白质。由于组织电导率因个体而异,本研究通过从代表健康人体组织特性的分布中为每个头部模型分配不同的电导率值来模拟这种变异性。然后在SimNIBS中模拟给定强度的tACS以估计大脑中诱导电场的大小。最后,将每个头部模型转换到归一化的FreeSurfer(fsavg)空间,以便进行被试间分析。结果使用Gmsh和MATLAB 2023a进行分析。
使用HCP-MMP图谱展示结果。本研究使用人类连接组计划的多模态分区图谱(HCP-MMP)来获取不同脑区的平均结果。HCP-MMP将大脑分为360个区域(每个半球180个)。在讨论这些区域时,将使用术语“子分区”或“子区域”。此外,本研究还使用HCP-MMP的皮层下区域(每个半球22个)。本研究将这些区域称为“皮层分区”或“区域”。电场值和性能指数按子区域进行平均,并按节点面积进行加权。
各研究的平均电场。本研究确定了64种针对前额叶、顶叶和颞叶新皮质区域的干预类型。为了探索可能的区域偏差,本研究计算了每个区域节点上的平均电场值,并按节点面积进行加权,以考虑有限元模型中的可变单元尺寸。
WM PEI。为了确定tACS电场与WM表现改变有因果关系的脑区,本研究估计了PEI(性能-电场指数)。在头部模型的每个表面节点(或三角形区域),电场值与Hedges’g值进行相关。在主要分析中,节点数对应于FreeSurfer平均(fsavg)大脑空间(327684个节点)。在有298个结果测量值的情况下,每个头模型会产生一个327648×298的电场值矩阵。然后,估计上述矩阵中与特定条件(例如,θ刺激)相关的列,选择与相应结果(Hedges’g)值之间的线性Pearson相关性。此过程会在fsavg空间中生成一个包含327648个PEI值的向量。对于一部分分析,本研究观察了HCP-MMP分区(每半球180个分区)和皮层分区(每半球22个区域)。为每个头部模型(100名被试)生成PEI图,并通过对模型之间的PEI值进行平均来将其合并到单个大脑空间中。在这里,将所有WM结果测量视为彼此独立,无论它们是在同一实证研究中估计的还是在不同研究中估计的,因为在大多数实证研究中缺乏足够的个体数据,无法评估结果之间的假定关联程度。PEI值低于0表示电场与Hedges’g之间存在负相关,即该区域上的tACS与WM表现下降相关。PEI值高于0表示电场和Hedges’g之间存在正相关,即该区域上的tACS与WM表现改善相关。
结果
各研究中最大的平均诱导电场表明,额叶皮层是最常见的靶点(图1A)。最小的平均电场位于枕叶区域(平均最大值的46.3%)。研究总共报告了64种干预类型或子研究,通常每种干预类型都有若干结果指标。这些结果指标包括WM准确性和一个或多个WM任务的处理时间以及难度水平。在总共298个结果测量值中,有58.4%表征了θ振荡(3-8Hz)的调制,35.2%表征了γ振荡(30-100Hz)(图1B)调制,而在其余6.4%的研究中,则表征了其他频率。结果测量涵盖了多种WM模式。WM刺激表现为视觉刺激(82.8%)、听觉刺激(14.3%)或两者结合(2.9%)。认知任务测量了语音-言语(字母或单词)、语音-数字、空间、物体和颜色记忆域。总的来说,现有数据很好地覆盖了大脑解剖空间,从而使我们能够进行因果映射。本研究将后续的频率分析重点放在迄今为止研究最多的θ和γ频段上,并通过对不同任务领域进行平均,将WM分析重点放在领域一般性特征上。
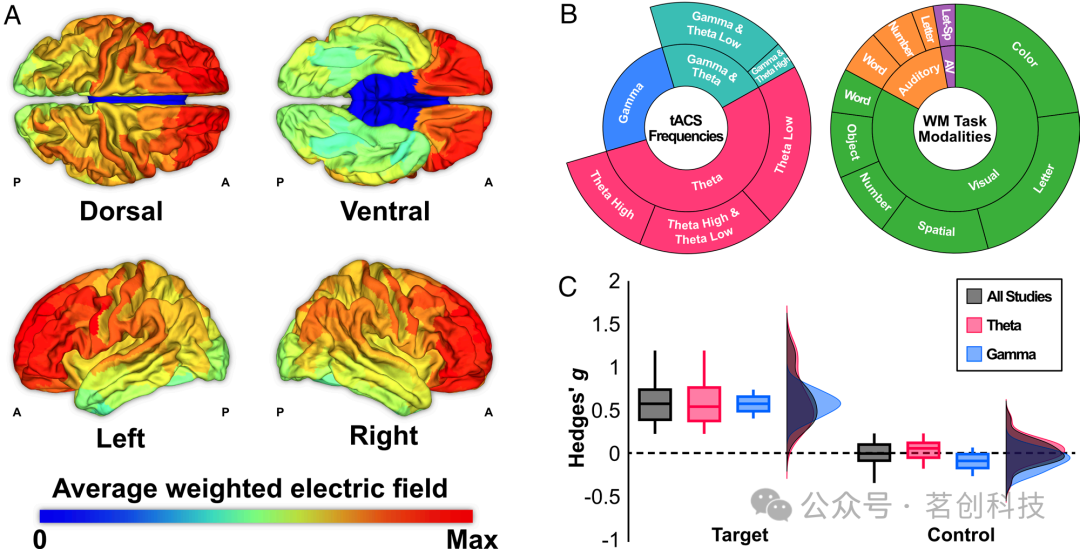
图1.对现有WM研究的概述。
这些结果测量值既反映了“目标”条件(通常是主要干预措施),也反映了各种控制条件,例如非目标刺激位置、频率和强度。当仅考虑目标条件时,tACS对WM的表现具有积极效果(Hedges’g=0.54;图1C)。然而,控制条件并未改变WM表现。这些无显著性条件对元建模方法至关重要,因为它们为我们的统计分析提供了脑图每个位置的方差,以及关于无显著性脑振荡位点的信息。
WM中θ和γ振荡的功能图
为了绘制领域一般性WM中θ和γ振荡的因果关系,本研究分析了tACS诱导的大脑电场强度与每个空间节点、每个结果测量值以及整个神经正常成人虚拟群体(N=100)的行为效应大小之间的关系。这种性能-电场指数(PEI)代表了特定脑区在记忆加工中可能起因果作用的可能性及其参与程度。由此产生的群体平均地形图包含正负PEI值,分别对应于因频率特异性脑刺激而改善或降低的WM表现。
额颞区θ节律同步导致显著的正PEI值(PEIMAX=0.260,P<0.001;图2和3)。特别是,左右眶额区(右:PEI=0.214,P=0.005;左:PEI=0.150,P=0.049)、右内侧前额叶皮层(PEI=0.179,P=0.018)、右内侧颞叶皮层(PEI=0.166,P=0.029)和右侧岛叶区域(PEI=0.175,P=0.021)的因果改善非常明显。
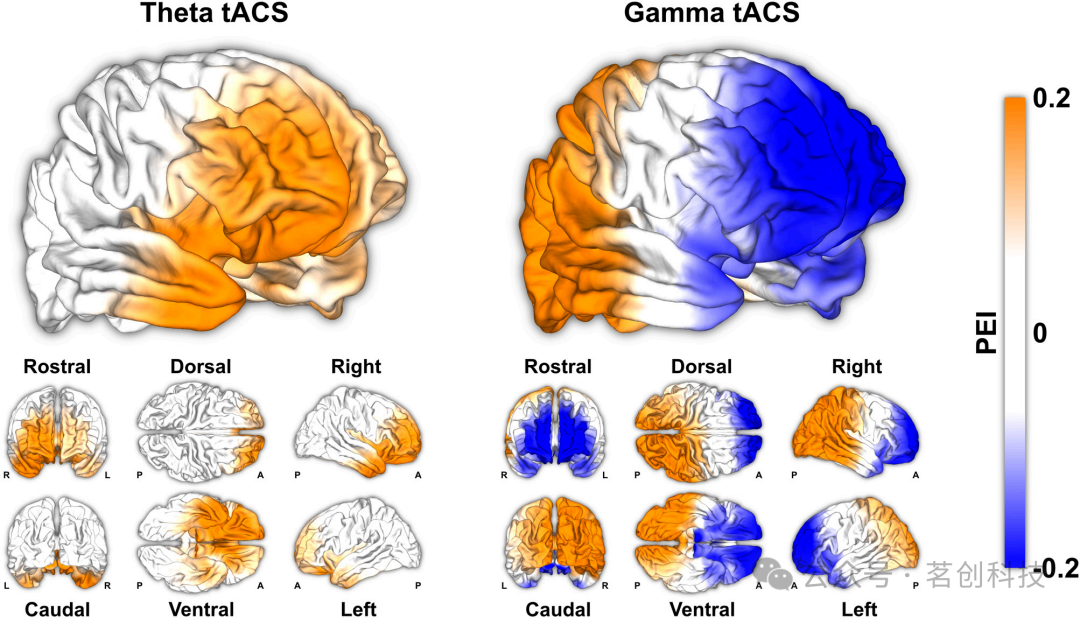
图2.WM中大脑振荡的因果功能图。
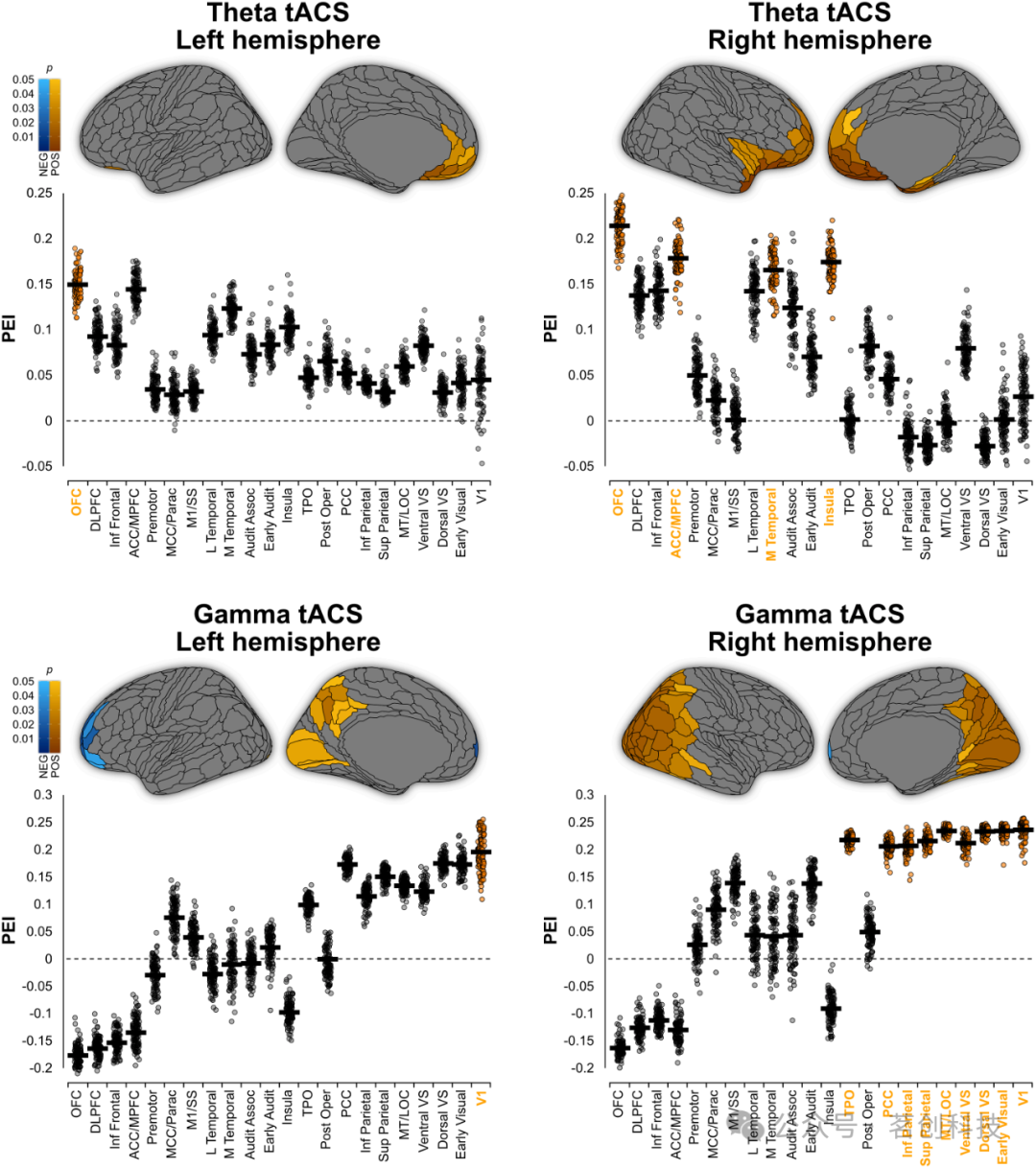
图3.分区后的功能图。
对于γ节律同步,双侧枕叶、顶叶和额叶区域出现了显著效应(PEIMAX=0.259,P<0.001;图2和图3)。正PEI出现在右后和枕叶皮质区,特别是早期视觉区域(PEI=0.235,P=0.021)、背侧视觉流(PEI=0.234,P=0.016)、腹侧视觉流(PEI=0.213,P=0.029)、枕外侧复合体(PEI=0.235,P=0.016)、颞顶枕交界处(PEI=0.219,P=0.025)、后扣带皮层(PEI=0.206,P=0.035)、下顶叶(PEI=0.207,P=0.034)和上顶叶(PEI=0.216,P=0.027)。此外,本研究发现双侧初级视觉区V1具有明显的正PEI(左:PEI=0.195,P=0.046;右:PEI=0.237,P=0.015)。亚区分析显示,84个区域(左半球和右半球分别为20个和64个;PEI范围:-0.240至0.241,P=0.049至0.013;图3)具有显著的PEI,横跨Brodmann区域7(顶上沟)、31(背侧后扣带皮层)、39(颞顶交界处)和40(顶内沟)。重要的是,在Brodmann区域9和10内的眶额区域和背外侧前额叶皮层发现了γ调制和WM之间存在显著的负相关关系(PEI=-0.240至-0.195,P=0.013至0.046)。这表明,枕顶区的γ振荡同步改善了WM表现,而额极区域的γ振荡同步则会降低WM表现。
低频和高频θ振荡的不同作用
鉴于低频和高频θ振荡在大脑中可能具有不同的作用,本研究考察了领域一般性WM中低频和高频θ振荡的因果空间图。对于低频θ振荡(3.5-6Hz),眶额区(PEI=0.247,P=0.006)、前扣带皮层(PEI=0.199,P=0.029)、内侧颞叶皮层(PEI=0.217,P=0.017)、外侧颞叶皮层(PEI=0.204,P=0.025)和岛叶(PEI=0.208,P=0.022;图4)与WM 表现呈正相关。这些效应发生在右半球,并且没有观察到负相关关系。对于高频θ振荡(6-8Hz),左额下皮层(PEI=0.326,P=0.017)、左眶额皮层(PEI=0.305,P=0.026)和左背外侧前额叶皮层(PEI=0.272,P=0.049)与WM表现呈显著正相关。相反,高频θ振荡显示在特定视觉和顶叶皮层亚区的WM表现有下降的趋势。
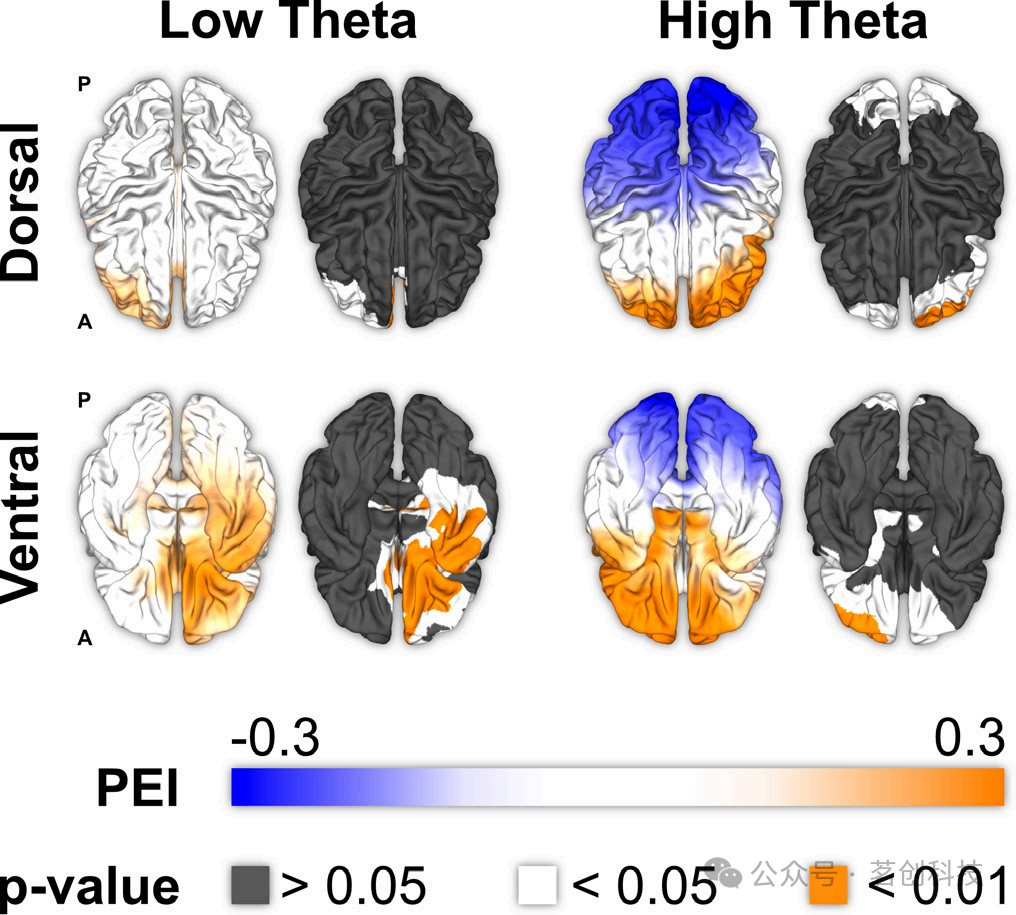
图4.低频和高频θ振荡的因果功能图。
WM准确性与加工时间中的θ和γ振荡
最后,本研究考察了θ和γ振荡是否对WM准确性(例如,正确反应的百分比、错误次数、灵敏度)和加工时间(例如,反应时、完成时间)产生不同影响。研究结果发现,准确性与θ振荡呈正相关,特别是在右侧眶额叶皮层(PEI=0.218,P=0.012;图5)、前扣带皮层(PEI=0.184,P=0.034)、内侧颞叶皮层(PEI=0.187,P=0.031)和岛叶(PEI=0.187,P=0.031)。WM准确性的提高与后扣带皮层(PEI=0.286,P=0.027)、上顶叶(PEI=0.305,P=0.018)和下顶叶(PEI=0.305,P=0.018)、颞顶枕交界处(PEI=0.322,P=0.012),以及所有枕叶皮层区域(P<0.05)的γ振荡有关,并且形成一个显著的簇(pcluster=0.031)。眶额皮层的一些子区域,特别是Brodmann区域10和11(PEI=-0.341至-0.260,P=0.007至0.045),显示γ节律与WM准确性之间呈负相关。虽然没有单个完整的皮层区域达到显著性水平(P>0.05),但确实识别出了一个显著的后部簇(pcluster=0.004)。在加工时间方面,无论是θ还是γ振荡,均未达显著性水平(P>0.05)。然而,前额叶的八个亚区(在Brodmann区域9和10内,左和右)显示出θ节律与加工时间之间呈正相关关系(PEI=0.368至0.310,P=0.018至0.049)。
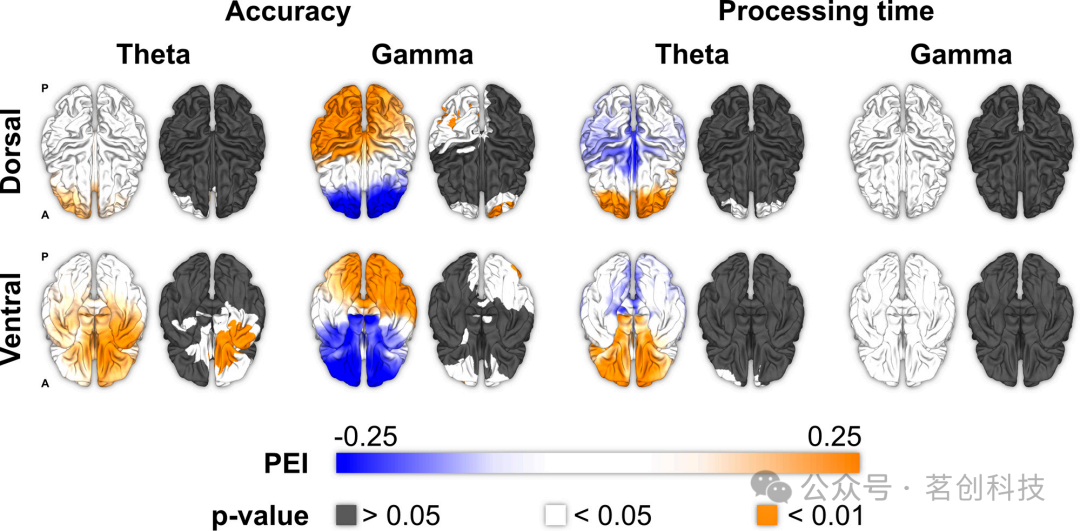
图5.WM准确性和加工时间。
结论
总的来说,前额叶和内侧颞叶的θ振荡以及枕顶叶的γ振荡是控制工作记忆(WM)表现的重要功能解剖中枢。本研究的功能图谱综合了十多年来使用安慰剂对照的经颅交流电刺激方法进行现代神经调控研究的结果。这些结果为工作记忆的机制理解提供了新的视角,并为认知障碍中工作记忆缺陷的神经调控治疗提供了基于假设驱动的治疗靶点。
参考文献:Miles Wischnewski, Taylor A. Berger, Alexander Opitz, Ivan Alekseichuk, Causal functional maps of brain rhythms in working memory. https://doi.org/10.1073/pnas.2318528121
小伙伴们关注茗创科技,将第一时间收到精彩内容推送哦~


















 1129
1129

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









