









De novo biosynthesis and nicotinamide biotransformation of nicotinamide mononucleotide by engineered yeast cells
经工程化酵母细胞实现烟酰胺单核苷酸的从头合成与烟酰胺的生物转化

摘要
β-烟酰胺单核苷酸(NMN)是哺乳动物体内烟酰胺腺嘌呤二核苷酸(NAD⁺)的前体。关于NAD⁺的研究表明,其在抗衰老和疾病防治中起着关键作用。本研究在酵母毕赤酵母(Pichia pastoris)中建立了两种高效合成NMN的技术路径,实现了以低成本的烟酰胺(NAM)或基础碳源为原料生产NMN。通过系统性改造酵母宿主以适应NMN的生物合成和积累。
为提高从NAM半生物合成NMN的效率,在细胞内表达烟酰胺磷酸核糖转移酶以评估其催化活性。此外,通过共表达NMN转运蛋白进一步提高了细胞外NMN的积累量。通过精细调控基因表达水平,在摇瓶实验中从NAM生产出72.1 mg/L的NMN。
为了实现NMN的从头合成,在酵母细胞中重建了异源生物合成途径。通过调节基因表达水平和增强前体生成能力,优化途径节点,实现了以葡萄糖(36.9 mg/L)或乙醇(57.8 mg/L)为碳源的高效NMN合成。在生物反应器中,通过补料分批培养,从2 g NAM和868 g葡萄糖在165小时内获得了1004.6 mg/L的NMN;从160 g葡萄糖和557 g乙醇在91小时内获得了980.4 mg/L的NMN。
本研究为利用工程化酵母细胞工厂进一步优化NMN的生物合成奠定了基础。
通过代谢工程改造毕赤酵母(P. pastoris),实现了NMN的半生物合成和从头生物合成,并创造了目前已报道的以基础碳源从头合成NMN的最高产量。
引言
β-烟酰胺单核苷酸(NMN)是烟酰胺腺嘌呤二核苷酸(NAD⁺)的关键中间体,而NAD⁺是人体及其他哺乳动物细胞中参与氧化还原反应的必需辅酶。NAD⁺在基因表达、DNA修复和能量生成等多种生物过程中发挥着核心作用,同时在代谢途径中,如三羧酸循环和糖酵解中,作为调节因子起着重要作用(Yoshino等,2021)。最近的研究表明,通过补充NMN,可显著减轻小鼠与年龄相关的生理退化,并逆转伴随的线粒体功能障碍(Gomes等,2013;Mills等,2016)。此外,NMN已被证明对因高脂饮食导致的2型糖尿病具有显著治疗效果(Yoshino等,2011)。临床试验显示,补充NMN可增强人类骨骼肌的葡萄糖代谢(Yoshino等,2021)。鉴于其显著效果,NMN已成为医药、食品和化妆品添加剂领域的热门原料。因此,开发一种具有成本效益且符合生物安全要求的NMN生物生产工艺具有重要价值。
NMN的天然生物合成路径可分为以下几类:(1)在哺乳动物(Guo等,2017;Lin等,2016)和某些细菌(Lee等,2017)中,NMN由烟酰胺磷酸核糖转移酶(Nampt)催化合成,以烟酰胺(NAM)和5-磷酸核糖-1-焦磷酸(PRPP)为起始原料;(2)在缺乏Nampt的细菌中,NMN主要通过NAD⁺降解生成(Göckel & Richert, 2015);(3)在同时缺乏上述两种路径的细菌中,以Francisella tularensis为例,NMN由NMN合成酶(FtNadE)催化,由烟酸单核苷酸(NaMN)转化而成(Sorci等,2009);(4)在某些哺乳动物中,NMN通过烟酰胺核糖激酶(Nrk)催化的磷酸化过程由烟酰胺核糖(NR)合成(Yoshino等,2018)。
最近,与化学合成相比,通过关键中间体进行生物转化已成为生产NMN的优选策略。Liu等(2021)和Maharjan等(2021)通过全细胞催化,从NAM分别合成了496.2和771.5 mg/L的NMN。Shoji等(2021)进一步在Bacillus mycoides中鉴定出NMN转运蛋白(BMpnuC),使通过全细胞催化从NAM生产NMN的产量提高到6.79 g/L。Huang等(2022)通过代谢工程改造大肠杆菌的半生物合成途径,利用来自噬菌体KVP40的烟酰胺磷酸核糖转移酶(Nampt,VpNadV),实现了16.2 g/L的NMN产量。此外,研究人员还报道了以替代底物烟酸(NA)进行NMN半生物合成的策略,但由于NA成本高且转化效率低,并未被广泛采用。
尽管通过大肠杆菌的半生物合成实现了高水平的NMN生产,但其在食品应用中可能存在内毒素残留的安全隐患,且需要严格的纯化过程。因此,FDA认定为安全(GRAS)的菌株可作为替代宿主。毕赤酵母(Pichia pastoris,又称Komagataella phaffii)是一种GRAS酵母宿主,广泛用于药物和食品添加剂的生产。其工业应用的优势包括培养基简单、温和的翻译后修饰、强大的分泌能力以及良好的多酶共表达能力(Meehl & Stadheim, 2014;Peña等,2018;Safder等,2018)。相比之下,目前报道的技术路线主要依赖于以NAM为必需底物的NMN半生物合成。而NAM主要通过化学合成制备,使NMN半生物合成实质上成为一种化学生物结合策略。此外,NMN可通过其内源性的NAD⁺补救途径自然生成,这一过程可被利用作为替代的NMN生物合成途径。最近研究探索了从天冬氨酸(Asp)开始的从头NMN生物合成路径,从1.00 g/L Asp底物中获得了153 mg/L的NMN(Wang等,2024)。然而,NMN的内源从头生物合成路径通常从蛋白原性氨基酸启动,并在微生物细胞中受到严格调控(Begley等,2001)。最近,通过从莽草酸开始的NAD⁺生物合成路径在大肠杆菌中实现了高水平的NAD(H)生产(Ding等,2021)。莽草酸因其作为细胞代谢自然分支点的代谢多功能性,成为了极佳的前体(Dosselaere & Vanderleyden, 2001)。这启发我们在酵母细胞中构建异源莽草酸-喹啉酸(QA)路径,并通过内源性的QA磷酸核糖转移酶(NadC)将QA转化为NaMN(图1)。随后,由NMN合成酶催化NaMN转化为NMN,从而通过以基础碳源培养的工程化细胞实现NMN的从头合成。
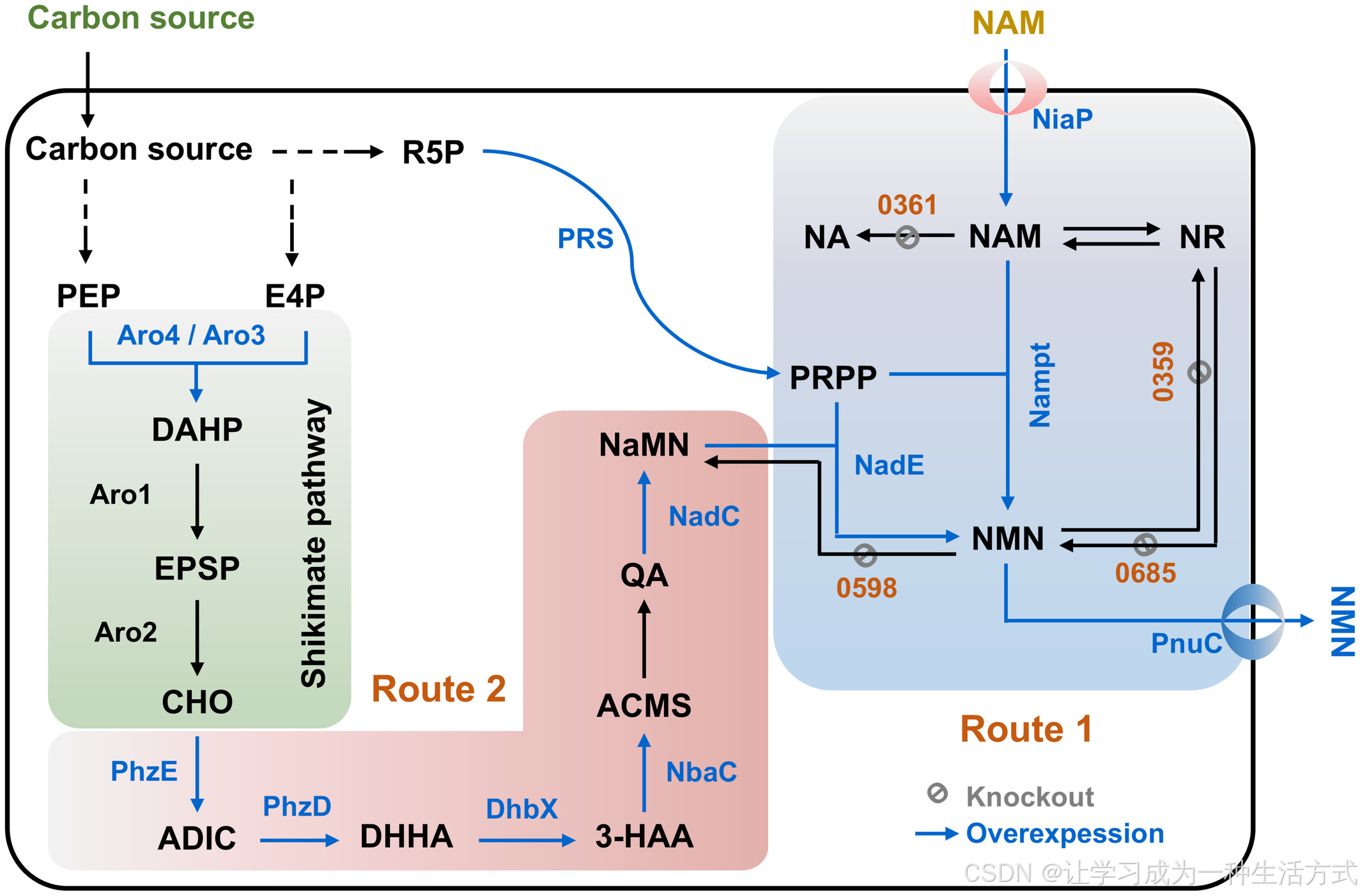
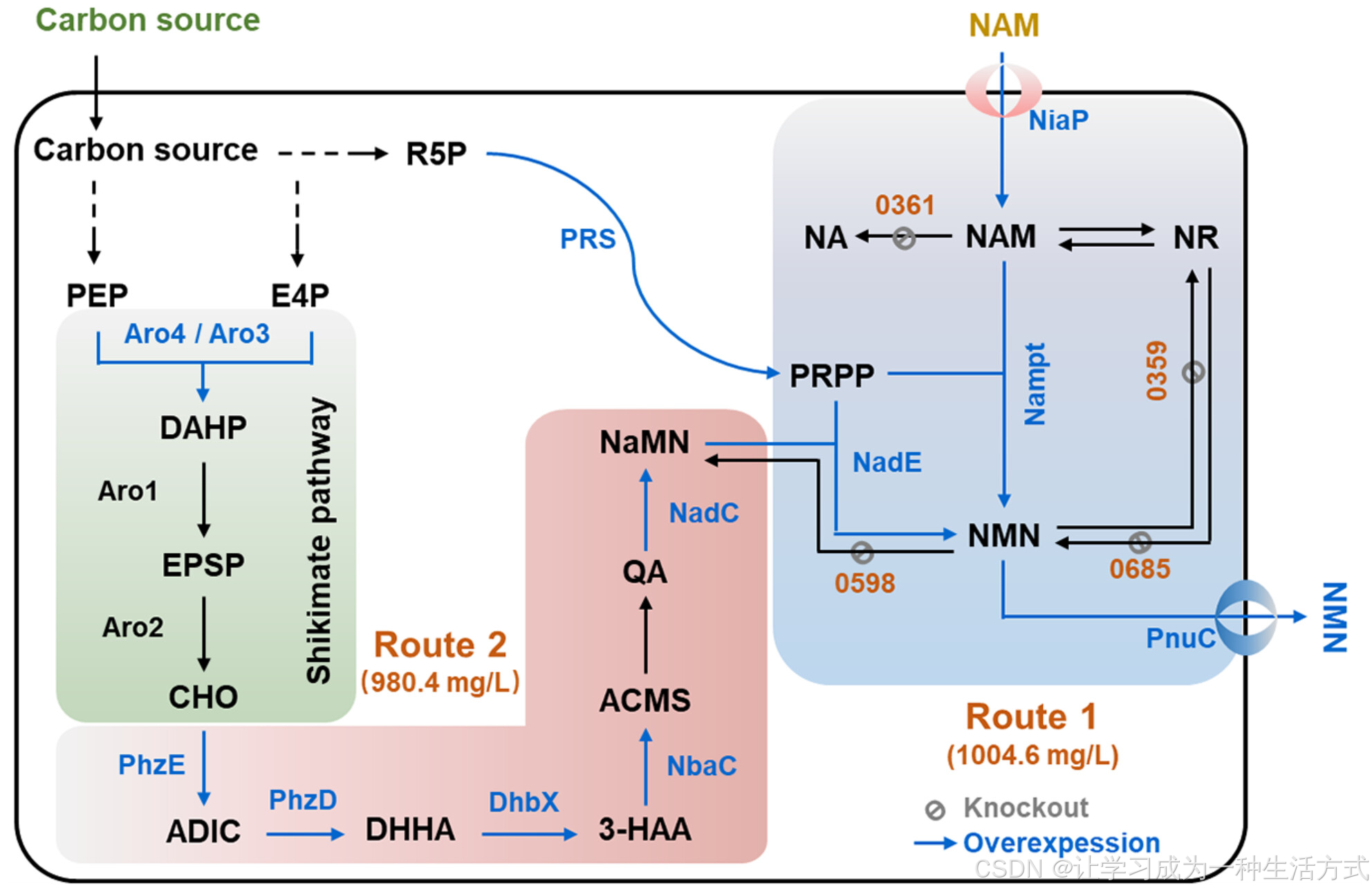
图1 代谢工程策略用于NMN的生物合成 路线1:通过在细胞培养过程中补充NAM实现NMN的半生物合成; 路线2:通过构建异源NMN合成路径实现NMN的从头生物合成。 缩写说明:
-
PEP:磷酸烯醇丙酮酸
-
E4P:赤藓糖-4-磷酸
-
R5P:核糖-5-磷酸
-
DAHP:3-脱氧-D-赤藓庚糖酸-7-磷酸
-
EPSP:5-烯醇丙酮基莽草酸-3-磷酸
-
CHO:莽草酸
-
AIDC:2-氨基-2-脱氧异莽草酸
-
DHHA:2,3-二氢-3-羟基蒽尼酸
-
ACMS:2-氨基-3-羧基粘康酸半醛
-
3-HAA:3-羟基蒽尼酸
-
PRPP:5-磷酸核糖二磷酸
-
NaMN:烟酸单核苷酸
-
QA:喹啉酸
-
NA:烟酸
-
NR:烟酰胺核糖
关键酶和蛋白:
-
PhzE:ADIC合酶
-
PhzD:DHHA合酶
-
DhbX:DHHA脱氢酶
-
NbaC:3-HAA 3,4-双加氧酶
-
NadC:QA磷酸核糖转移酶
-
NadE:NMN合成酶
-
Nampt:烟酰胺磷酸核糖转移酶
-
NiaP:NAM转运蛋白
-
PnuC:NMN转运蛋白
-
Aro4和Aro3:DAHP合酶
-
Aro1:五功能芳香族蛋白
-
Aro2:莽草酸合酶
-
PRS:PRPP合酶
-
0361:编码烟酰胺酶的基因PAS_chr3_0361
-
0598:编码假定蛋白的基因PAS_chr2-1_0598
-
0359:编码核糖基烟酰胺激酶的基因PAS_chr2-2_0359
-
0685:编码核苷酸酶的基因PAS_chr2-1_0685
本研究旨在通过工程化毕赤酵母实现NMN的半生物合成和从头生物合成,并评估两种技术路线的生产效率(图1)。采用系统性代谢工程策略减少NMN的旁路消耗并加强其生成路径。最终,在3 L生物反应器中,通过在半生物合成中添加NAM或通过基础碳源进行从头生物合成,分别获得了约1000 mg/L的NMN。本研究为未来NMN的全生物合成研究提供了有价值的参考。
实验方法
菌株、培养基和生长条件
本研究中使用的质粒和菌株列于附表S1和S2。用于质粒存储和扩增的大肠杆菌E. coli Top10,以及表达载体pGAPZ B和pUC18,均存储于实验室。所有化学试剂均购自Tansoole(上海,中国)。大肠杆菌和酵母菌株的培养条件参考文献(Ren等,2020)。对于平板培养,加入2%(质量/体积)的琼脂。
首先,将20 μL冻存菌株(−80°C)接种到3 mL YPD培养基中,在20 mL血清瓶中培养36–48小时。然后,将100 μL菌液接种到10 mL YPD培养基中,培养至OD600达到6.0。通过5000g离心3分钟收集细胞,并用无菌水重悬后以终浓度(OD600)1.0接种到发酵培养基中。在6孔深孔板中(每孔50 mL中装10 mL培养基)进行发酵培养以生产NMN。
发酵培养基
发酵培养基的组成包括:2%蛋白胨、1%酵母提取物、2%葡萄糖、0.1%硫酸镁七水合物(MgSO₄·7H₂O)、0.6%磷酸二氢钾(KH₂PO₄)、1.64%磷酸氢二钾(K₂HPO₄)、0.1%柠檬酸钠和0.05%烟酰胺(NAM,所有组分均以质量/体积计)。此外,每隔24小时添加2%(质量/体积)的葡萄糖至终浓度。
对于补料分批发酵,菌株在1 L摇瓶中200 mL YPD培养基中培养12-16小时作为种子液。将种子液转移至3 L发酵罐中的2 L发酵培养基中。在批次培养阶段,pH保持在4.5,补料阶段通过自动控制添加氢氧化铵(50%,体积/体积)将pH调整至7.0。通过自动控制通气和搅拌,溶解氧(DO)维持在30%以上。
质粒与菌株的构建
本研究中构建了大量质粒和重组菌株。质粒与菌株的详细构建信息见附加文件。所有质粒和菌株及其对应的基因型信息分别列于附表S1和S2中。用于质粒构建的引物列于附表S3中。基因操作通过之前开发的CRISPR-Cas9编辑工具完成(Liu等,2019a)。用于在毕赤酵母中表达的酶和转运蛋白的信息列于附表S4中。
分析方法
6孔深孔板中的NMN含量通过已报道的荧光衍生化方法进行分析(Marinescu等,2018)。取69 μL培养液样品至96孔板中,加入28 μL苯乙酮溶液(20% DMSO,体积/体积)和28 μL氢氧化钾溶液(2 mol/L)。在冰上孵育2分钟后,每孔加入125 μL甲酸溶液(88%,体积/体积),随后在37°C孵育10分钟。利用微孔板读数仪(BioTek Synergy 2)测定445 nm处的紫外发射强度,激发波长为382 nm。生物反应器中的NMN产量通过高效液相色谱法测定(Huang等,2022)。采用反相色谱柱(YMC-Triart C18,4.6×250 mm),以30 mM KH₂PO₄:甲醇(95:5,体积/体积)为洗脱液,流速为1 mL/min,检测波长设为254 nm。细胞湿重分析参考Xu等(2019)。
统计分析
数据来自三次生物重复实验,每次实验重复三次,结果以平均值±标准偏差表示。实验至少重复两次。采用独立样本Student's t检验评估组间数据差异,统计显著性定义为p < 0.05。
结果
构建用于NMN半生物合成的异源途径
由于毕赤酵母本身缺乏NAM到NMN转化的酶,首先在宿主中引入了来自哺乳动物和细菌的三种不同Nampt酶(图2A)。经过密码子优化的Nampt基因,包括来自Haemophilus ducreyi的Nampt-HD(Marinescu等,2018)、来自小鼠(Mus musculus)的Nampt-MM(Marinescu等,2018)和来自Chitinophaga pinensis的Nampt-CP(Shoji等,2021),在毕赤酵母Δku70突变体(部分缺乏非同源DNA末端连接,Liu等,2019a)中合成并表达。
对每个转化菌株的细胞裂解物进行的Western blot分析显示,所有Nampt酶均成功表达,但表达水平存在差异(图2B)。各菌株的NMN生产能力与其Nampt表达水平大致相关。来自C. pinensis的Nampt-CP表现出最高的NMN产量(图2C、D),与之前的研究结果一致(Shoji等,2021)。在0.5 g/L NAM补料条件下,Nampt-CP表达菌株在96小时内的NMN产量达到13.3 mg/L,比亲本菌株Δku70提高了71%(图2C)。此外,与Δku70相比,Nampt-CP表达菌株的细胞生长略有增加(图S1)。
值得注意的是,亲本菌株似乎通过内源性从头合成途径在极低水平上生成NMN(图2C中Δku70菌株在有无NAM补料条件下的表现)。综上所述,产量最高的Nampt-CP表达菌株被选择用于后续实验。
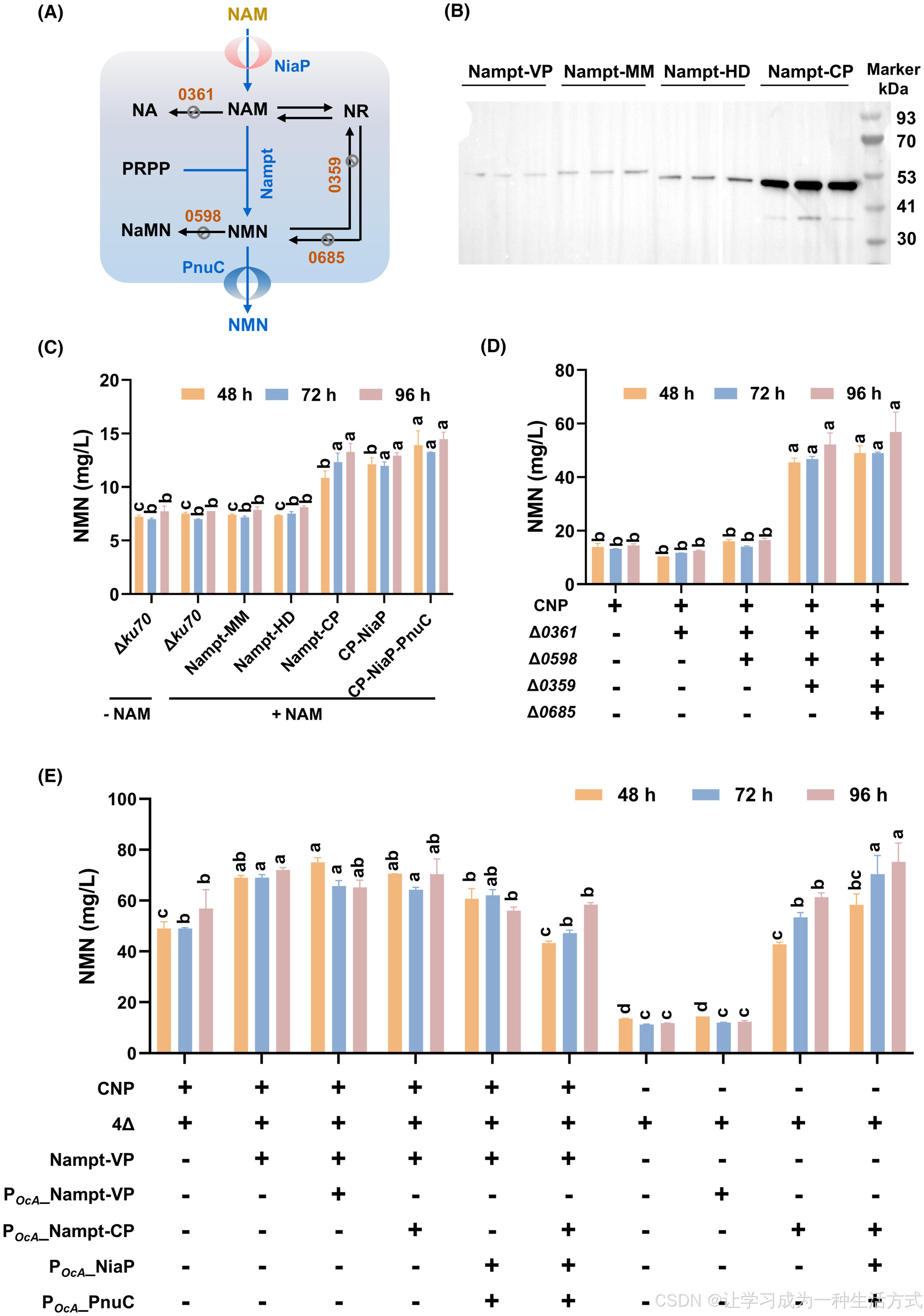
图2 通过NAM供给实现的毕赤酵母细胞NMN半生物合成 (A) NMN半生物合成的基本工程化方案; (B) 在毕赤酵母中表达的不同来源Nampt酶的Western blot分析:
-
来自小鼠(M. musculus)的Nampt-MM,56.7 kDa;
-
来自Haemophilus ducreyi的Nampt-HD,55.8 kDa;
-
来自Chitinophaga pinensis的Nampt-CP,53.6 kDa;
-
来自Vibrio噬菌体的Nampt-VP,55.6 kDa; (C) 表达不同来源Nampt酶和转运蛋白的菌株的NMN产量; (D) 在(C)的基础上敲除NMN/NAM消耗路径的菌株的NMN产量; (E) 改善Nampt酶和转运蛋白表达后菌株的NMN产量。
在毕赤酵母Δku70(关键的非同源末端连接修复基因KU70缺失)中,Nampt-MM、Nampt-HD、Nampt-CP、Nampt-VP、NiaP和PnuC基因的表达均由PGAP启动子(或在特殊说明时使用POcA启动子)驱动;CP-NiaP表示Nampt-CP和NiaP的共表达;CP-NiaP-PnuC(CNP)表示Nampt-CP、NiaP和PnuC的共表达。Δ0361、Δ0598、Δ0359和Δ0685分别表示对应基因的敲除,4Δ表示四个基因的同时敲除。
提高NMN产量的工程化策略
为增强NAM的吸收和NMN的分泌,合成了来自Burkholderia cenocepacia的NiaP和来自Bacillus mycoides的PnuC转运蛋白(Shoji等,2021),并将其与Nampt-CP在毕赤酵母中共表达。结果显示:在96小时培养后,在NAM(0.5 g/L)补料条件下,CP-NiaP(CN)和CP-NiaP-PnuC(CNP)菌株的NMN产量分别为12.9和14.5 mg/L,表明PnuC的表达使NMN产量提高了9%。
尽管CNP菌株有效将NAM转化为NMN,但观察到添加的NAM容易转化为烟酸(NA)(图S2),且NA与NAM直接竞争底物,抑制NMN的生成。通过敲除与NAM和NMN代谢相关的基因(PAS_chr3_0361、PAS_chr2-1_0598、PAS_chr2-2_0359和PAS_chr2-1_0685),分别构建了多重敲除菌株。96小时后,CPNΔ0361Δ0598Δ0359和CPN4Δ菌株的NMN产量分别为52.2 mg/L和56.9 mg/L,分别是CNP菌株的2.6倍和2.9倍。
通过使用更强的POcA启动子优化Nampt酶和转运蛋白的表达后,CPN4Δ-VP菌株在96小时的NMN产量达到72.1 mg/L,相比CPN4Δ菌株提高了26.7%。然而,进一步提高Nampt-CP和Nampt-VP的表达后,NMN产量未能进一步提升,可能是底物和产物的转运受到限制。
最终,选择CPN4Δ-VP菌株用于生物反应器的发酵实验。
NMN从头生物合成的设计与构建
NMN的半生物合成需要NAM作为直接底物,其成本远高于基础碳源,且工业用NAM主要通过化学合成获得。为降低成本并实现完全生物生产,研究设计了一条从基础碳源出发的NMN从头合成途径。在毕赤酵母中,内源NMN从头合成途径始于氨基酸,但受到细胞的严格调控。因此,研究通过在细胞中引入一条以莽草酸为起始物的异源合成途径,实现高效的NMN合成和积累(图3A)。
图3 通过异源合成途径实现毕赤酵母的NMN从头生物合成
(A) 工程化毕赤酵母细胞内的内源性和异源性NMN生物合成路径; (B) 在毕赤酵母中表达的NMN从头合成途径相关酶的Western blot分析:
-
NadE,29 kDa;NbaC,21 kDa;PhzE,69 kDa;NadC,33 kDa;PhzD,23 kDa;DhbX,27 kDa; (C) 系统代谢工程策略提高NMN产量。
Δku70、Δ0361、Δ0598、Δ0359和Δ0685的定义与图2相同。PhzE、PhzD、DhbX、NbaC、NadC、NadE、Nampt、PnuC、Aro4、Aro3K222L和PRS基因的表达均由PGAP启动子驱动,除非特别说明。其基因和蛋白功能如图1所述。
构建从头合成的宿主菌株
在敲除了四个基因(PAS_chr3_0361、PAS_chr2-1_0598、PAS_chr2-2_0359和PAS_chr2-1_0685)的Δku70Δ0361Δ0598Δ0359Δ0685菌株中,引入了以莽草酸为起点的异源合成路径(图3A)。该路径中包括的关键基因(PhzE、PhzD、DhbX和NbaC)经过密码子优化,并在Δku70Δ0361Δ0598Δ0359Δ0685中表达。此外,通过过表达NadC和NMN合成酶(FtNadE),实现喹啉酸(QA)到NaMN及NaMN到NMN的转化。同时,为增加5-磷酸核糖二磷酸(PRPP)供应,引入了经过改造以消除反馈抑制的BaPRSL135I。最终,通过Western blot分析确认了所有相关酶的成功表达(图3B)。
改造后菌株的NMN产量达到18.7 mg/L,相比于Δku70菌株和Δku70Δ0361Δ0598Δ0359Δ0685菌株分别提高了163%和68.5%(图3C)。进一步通过额外过表达NadE,NMN产量在24小时内达到31.9 mg/L,表明NadE是关键酶,其高表达显著提升了NMN合成效率。
为了提高莽草酸的前体供应,表达了Aro3K222L和Aro4K229L的反馈不敏感突变体,分别实现了26.4 mg/L和27.5 mg/L的NMN产量(图3C)。最终,构建的菌株(Δ0361Δ0598Δ0359Δ0685-NbaC-NadC-PhzE-PhzD-DhbX-2NadE-PRS-PnuC-Aro3K222L, 简称4ΔNNPPD2NPPA)在48小时内产生了36.9 mg/L的NMN,相比于未改造的Δku70菌株,产量提高了4.9倍。此外,当以乙醇作为碳源时,NMN产量达到57.9 mg/L,比葡萄糖作为碳源提高了60%(图S6)。
在生物反应器中进行NMN半生物合成与从头合成
为评估产能,分别在3-L补料分批发酵罐中培养了半生物合成菌株(CPN4Δ-VP)和从头合成菌株(4ΔNNPPD2NPPA)。
对于半生物合成,批次培养阶段将pH控制在4.5左右,随后在补料阶段调整至7.0。以40 g/L葡萄糖进行批次培养,湿细胞重达到约200 g/L后,将葡萄糖补料速率调整为2.5 g/L/h,并向培养液中添加2 g/L NAM。最终在165小时内积累了1004.6 mg/L的NMN,消耗了约1 g/L的NAM和868 g的葡萄糖(图4A)。
对于从头合成,补料阶段加入乙醇作为主要碳源。在类似条件下,NMN产量达到980.4 mg/L,证明从头合成路径能够有效替代NAM供给,同时降低生产成本。
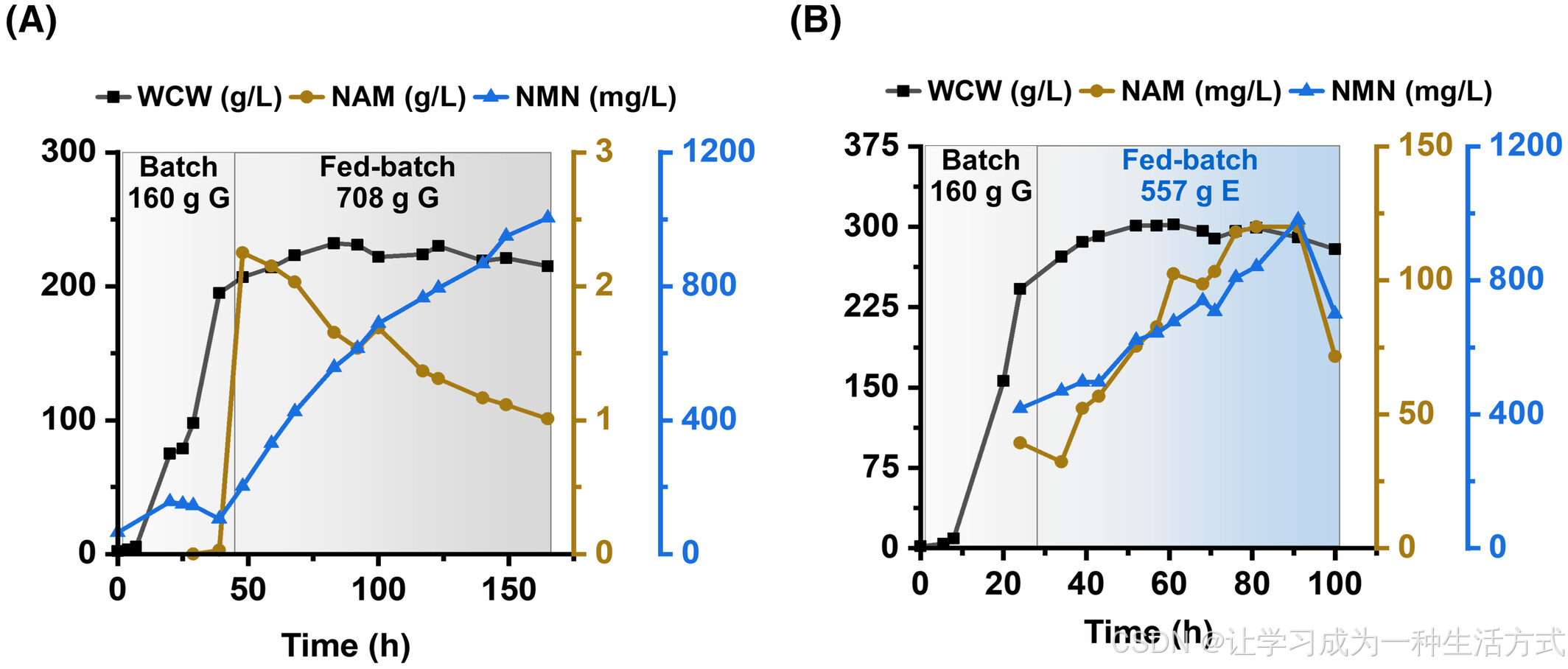
图4 P. pastoris 在 3L 生物反应器中生产 NMN (A) 使用 CPN4Δ‐VP 菌株通过 NAM 进料进行 NMN 的半生物合成;(B) 使用 4ΔNNPPD2NPPA 菌株从基本碳源(葡萄糖和乙醇)进行 NMN 的从头合成。在分批阶段,培养液 pH 控制在 4.5,随后在补料分批阶段调整至 7.0。E 表示乙醇;G 表示葡萄糖;WCW 表示湿细胞重量。
对于由 4ΔNNPPD2NPPA 菌株进行的 NMN 从头合成,pH 控制与半合成培养相同。分批培养中加入 40 g/L 葡萄糖,之后以 7.5 g/L/h 的速率补充葡萄糖,直到湿细胞重量达到约 200 g/L。随后,补料逐渐更换为乙醇,并以 4.5 g/L/h 的速率补充。NMN 的浓度在 91 小时内逐渐累积,最终从 160 g 葡萄糖和 557 g 乙醇中生产了 980.4 mg/L 的 NMN(图 4B)。同时,合成了 120 mg/L NAM 作为主要副产物。由于乙醇的价格低于葡萄糖和 NAM(NAM: ~20,000 美元/吨;乙醇: 250–350 美元/吨;葡萄糖: 300–400 美元/吨,Huang 等,2022;Sun 等,2024),NMN 的从头生物合成相比半生物合成可能是一种更具前景的技术路径。
讨论
NMN 是一种关键化合物,以其为哺乳动物细胞提供 NAD+ 的生理重要性而闻名,因而 NMN 产品迅速商业化(Yoshino 等,2021)。本研究开发了两种有前景的策略,即通过工程化 P. pastoris 进行 NMN 的一步活细胞发酵的半生物合成和从头生物合成。系统代谢工程方法被应用于 P. pastoris,以实现高效的 NMN 生物合成,包括阻断支路途径、表达和评估功能酶及转运蛋白、构建异源途径、平衡合成途径的表达等。通过这些努力,生物反应器发酵中,无论是通过 NAM 补充进行半生物合成,还是从基本碳源进行从头合成,都实现了约 1000 mg/L 的 NMN 产量。
目前,酶或全细胞催化是 NMN 合成的主要方法,但工艺复杂且成本较高(Shoji 等,2021)。近期,基于 NAM 补料的发酵方法在大肠杆菌中实现了高水平 NMN 的生产,工艺简单,生产效率和转化率高(Huang 等,2022),为 NMN 及其衍生物的半生物合成开辟了新途径。然而,生产宿主大肠杆菌并非食品领域的理想菌株。本研究采用一种类似的半生物合成方法,在安全的(GRAS)P. pastoris 中以简单工艺和低成本底物实现了 NMN 生产。更重要的是,我们还实现了一种新的从头合成 NMN 的技术路径,通过工程化 P. pastoris 菌株从基本碳源如葡萄糖和乙醇发酵实现。这种从头生物合成在酵母宿主中实现了与半生物合成相当的产量,但无需 NAM 补充且所需时间更短。总体而言,这为未来通过安全的微生物细胞工厂全面提升 NMN 的全生物合成提供了新思路。
据报道,来自不同来源的 Nampt 酶支持高水平半生物合成 NMN 的大肠杆菌菌株构建。这启发我们在本研究中适配不同的 Nampt 酶至酵母宿主,以高效将 NAM 转化为 NMN。C. pinensis 来源的 Nampt-CP 表现良好并促进了 NMN 的生产,与先前研究中其他宿主的结果一致(Shoji 等,2021)。然而,V. bacteriophage KVP40 来源的 Nampt-VP 在大肠杆菌中实现了最高水平的 NMN 生产(16.2 g/L,Huang 等,2022),其在 P. pastoris 中的功能表现较差,尽管对其进行了密码子优化和表达改进。这可能与 P. pastoris 表达的翻译和酶折叠不正确有关。鉴于其在大肠杆菌中的显著酶活性和催化效率,未来应进一步开展酶工程改进工作。
细胞内 NMN 的积累可能会妨碍生理过程。Shoji 等(2021)通过在工程化大肠杆菌中引入 B. mycoides 的 NMN 转运蛋白 PnuC,将细胞外 NMN 的累积量显著提高至 6.79 g/L。尽管在 P. pastoris 中过表达 PnuC 同样促进了 NMN 的生产,其效果却有限。深入探索更高效的转运蛋白将是进一步提高 NMN 生物合成的一种有效策略。然而,通过半生物合成策略,最终在生物反应器水平获得了 1004.6 mg/L 的 NMN,这代表了酵母中 NMN 生产的最高产量。未来需要进一步进行细胞重构和培养优化,以持续提高生产能力,因为与大肠杆菌的成果相比仍有显著差距。
相比之下,通过酵母发酵从基本碳源进行的从头合成混合途径代表了一种吸引人的 NMN 生产路径。通过异源莽草酸途径到 NMN 的重组、支路阻断以及代谢前体积累的改善,在生物反应器水平实现了与半生物合成相当的产量(980.4 mg/L),但所需时间更短。此外,乙醇作为碳源相较葡萄糖更具优势。这可能是由于乙醇的代谢途径本质上比葡萄糖更简单,同时乙醇的能量密度优于葡萄糖,从而为产品合成提供了更多的能量(Sun 等,2024)。未来应进行深度代谢重构和生物工艺优化以进一步提高产量。最近的一项研究中,通过改善大肠杆菌中从天冬氨酸到 NMN 的内源从头合成途径获得了 153 mg/L 的 NMN(Wang 等,2024),但仍需额外补充天冬氨酸作为底物。总体而言,本研究首次报道了通过酵母发酵从基本碳源构建从头合成 NMN 的途径。这是迄今为止从基本碳源通过从头合成获得的最高 NMN 产量,且与现有策略相比代表了一种新型生产模式,显示出未来工业应用的良好潜力。

















 866
866

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









