







–DOI: 10.1126/science.abf3041
Single-cell eQTL mapping identifies cell type–specific genetic control of autoimmune disease
留意更多内容,欢迎关注微信公众号:组学之心
研究团队和研究单位
:::: column
::: column-left
Joseph Powell–Garvan Institute of Medical Research

:::
::: column-right
Alex W. Hewitt–Menzies Institute for Medical Research, University of Tasmania

:::
::::
文章简介
研究背景
基因表达在免疫细胞中的个体差异既是免疫相关疾病易感性的原因,也是其结果。研究发现多种遗传变异影响免疫调节和疾病发展。然而,由于循环免疫群体的复杂性,这些变异的作用机制难以解析。通过结合转录谱和遗传变异,可以识别基因表达的调控因子,尤其是全基因组关联研究(GWAS)中与常见免疫疾病相关的调控区变异。有研究表明,这些变异多通过改变基因表达而非直接影响蛋白质功能发挥作用。
利用bulk RNA测序与遗传信息结合,发现了表达数量性状位点(eQTL),其中很多是组织特异性的,但难以揭示单细胞间的基因表达异质性。确定疾病风险单核苷酸多态性(SNP)影响基因表达的细胞和环境背景,有助于理解疾病机制和制定治疗策略。传统方法如FACS和反卷积分析虽然识别了细胞类型特异性的eQTL,但有其局限性,难以识别稀有细胞类型和捕捉细胞内的异质性。
研究内容
-
单细胞RNA测序的应用: 使用单细胞RNA测序(scRNA-seq)数据,对982名健康人的1,267,758个外周血单核细胞进行分析,识别了26,597个顺式表达数量性状基因座(eQTL)和990个反式eQTL,揭示了这些变异的细胞类型特异性。
-
动态等位基因效应分析: 研究展示了B细胞从幼稚到记忆状态转变中的动态等位基因效应,以及等位基因如何导致个体间免疫功能差异。
-
孟德尔随机化方法: 使用孟德尔随机化方法确定了305个风险基因座在细胞水平上导致自身免疫疾病的因果途径。
研究意义
确定了与疾病相关的单核苷酸多态性(SNP)对基因表达的细胞和环境背景影响,有助于理解疾病机制和制定治疗策略。
研究结果
1.OneK1K 队列
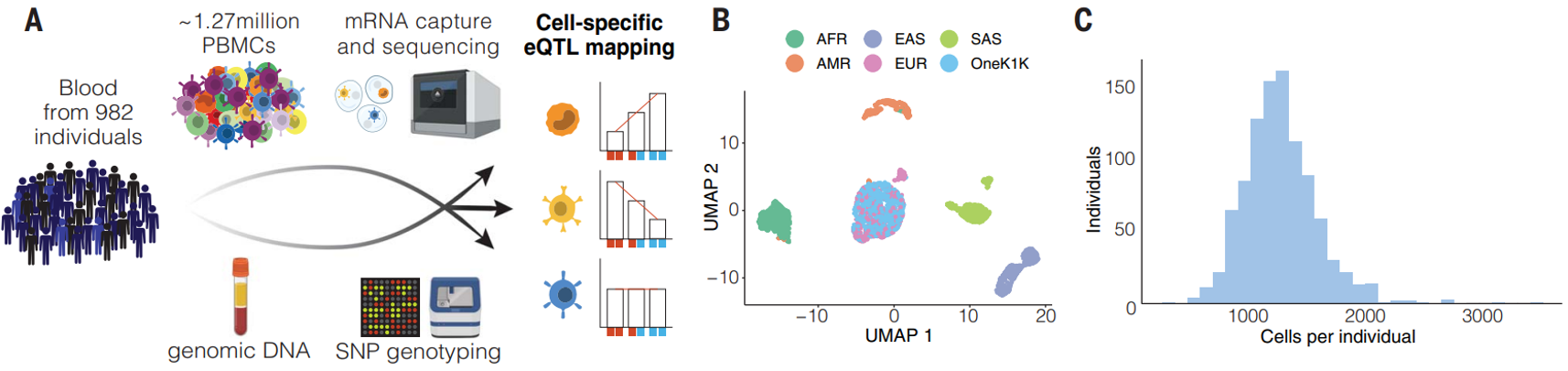
(A)样本取自 982 名健康个体外周血,通过整合 scRNA-seq 和 SNPs 基因分型数据进行单细胞eQTL分析,表征了一个大型队列 (OneK1K) 中循环免疫细胞的转录变异,以探索等位基因变异如何以细胞类型特异性的方式与基因表达的变化相关联。
(B)UMAP 分析显示 OneK1K 队列的个体与 1000 基因组计划之间的遗传关系。OneK1K 队列的个体与具有北欧遗传血统的个体嵌入其中,说明队列是由北欧血统的个体组成。其中:AFR, African; AMR, ad-mixed American; EAS, East Asian; EUR, European; SAS, South Asian.
(C)单细胞数据质控后,保留了 1,267,758 个细胞以供进一步分析。
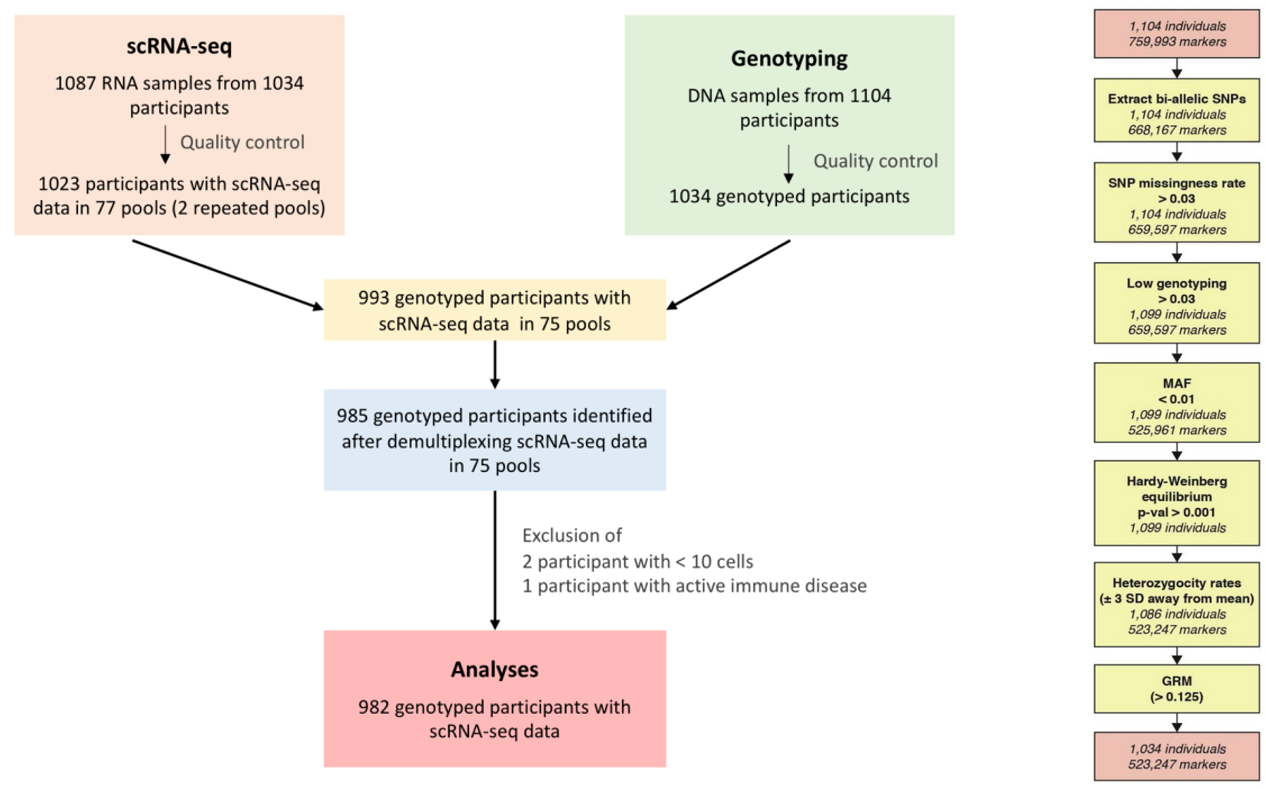

生成了 759,993 个 SNP 的基因型数据,并根据Haplotype Reference Consortium panel 估算了 SNP。经过质量控制后,研究保留了 5,328,917 个次要等位基因频率大于 0.05 的 SNP,和 265,023个外显子markers。
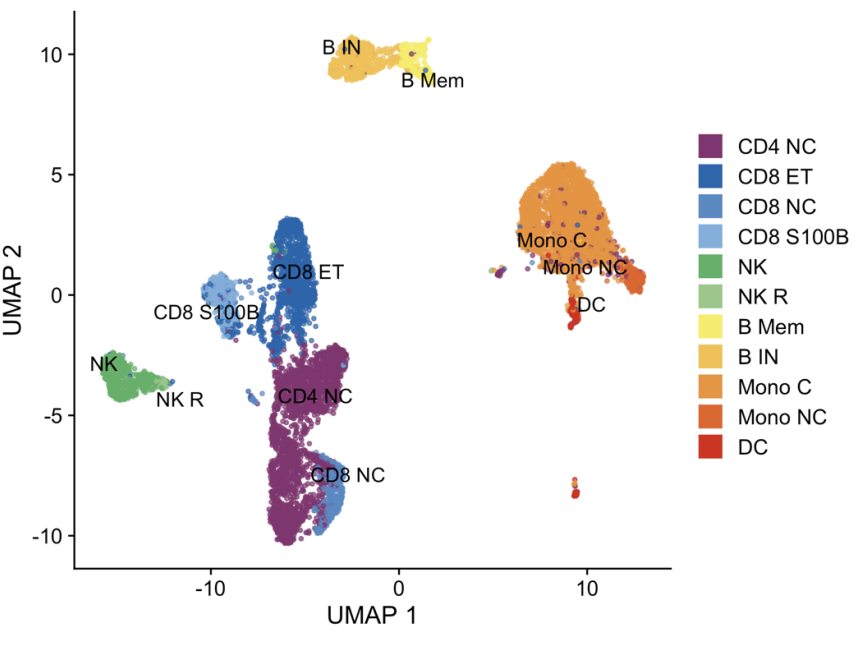
2.单细胞亚型分类
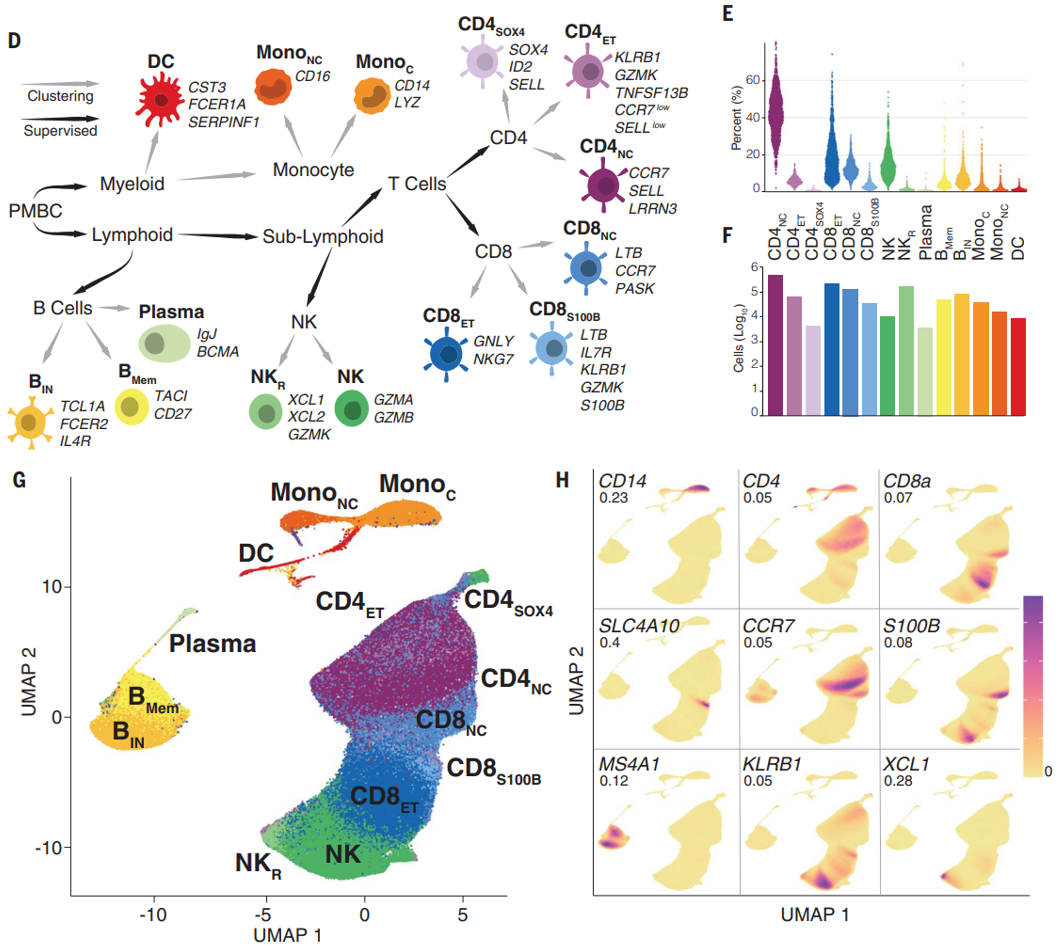
(D)作者先前开发了一个细胞分类框架–scPred,根据每个细胞的转录组数据,可以将其独立地分类为髓系和淋巴系中 14 种不同的免疫细胞类型之一。该框架结合了分层监督和非监督分类方法,并使用 FACS 分类的 PBMC scRNA-seq 数据作为参考。
(E-F)细胞组成范围从 0.7% 的树突状细胞 (DC) 到 36.6% 的 CD4+ 幼稚细胞和中央记忆 T (CD4NC) 细胞,比例的平均值和范围与其他研究的相符。
(G-H)使用均匀流形近似和投影 (UMAP) 对细胞类型进行可视化这些细胞类型之间的聚类情况,并展现了一些细胞亚群的biomarker,表明scPred分类的准确性很好。
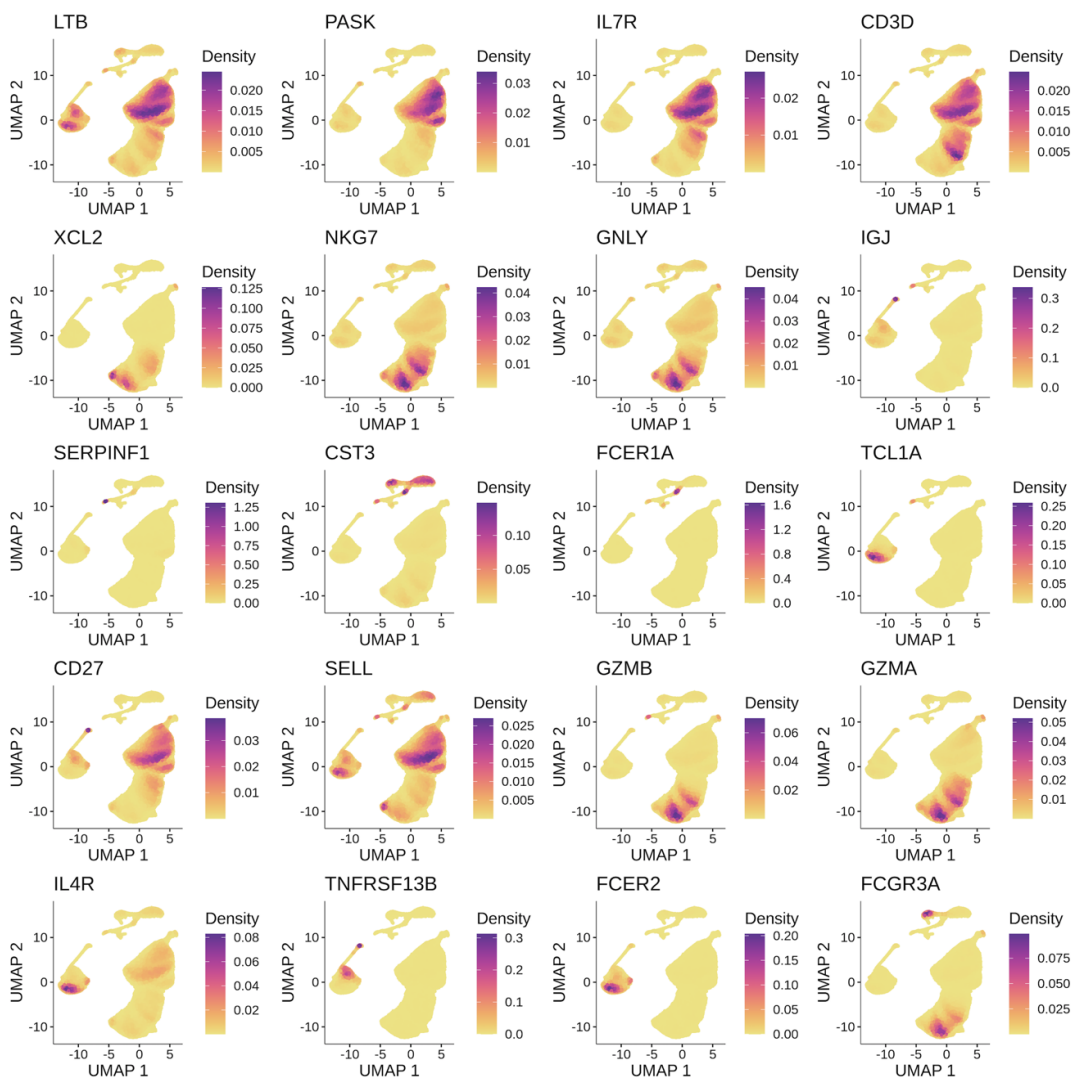
更多的细胞类型biomarkers展示。
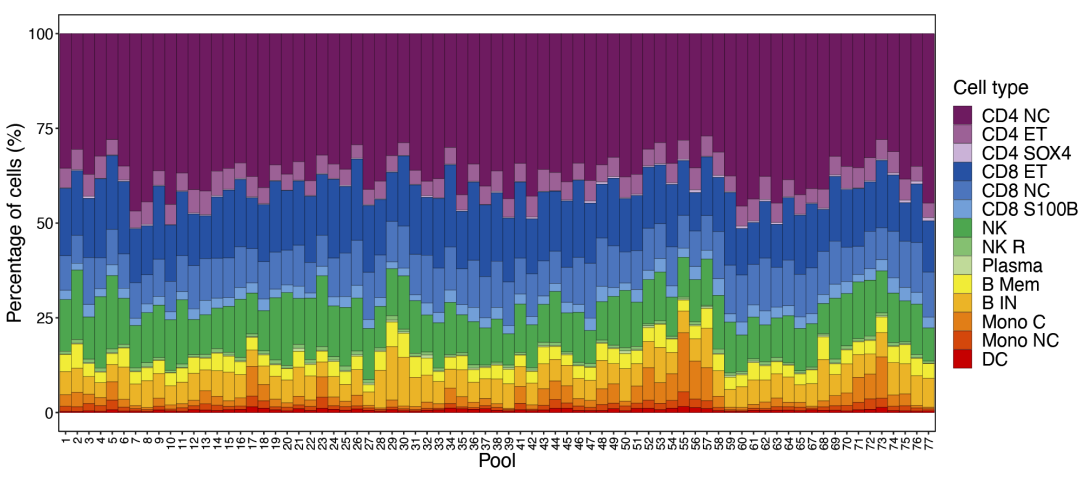
75 个测序池中细胞类型比例的分布。每个包含 12-14 个单独的样品。池 40 和 66 是重复的,已被删除。
虽然大多数个体都有所有 14 种细胞类型的 scRNA-seq 数据,但由于抽样差异,一些细胞类型 (主要是表达 SOX4 的 CD4+ T 细胞、浆细胞和非经典单核细胞)未被测序。因此,后续 eQTL 分析的样本量因细胞类型而异。
3.单细胞 eQTL 分析揭示了因常见变异而发生的转录变化的细胞类型特异性
eQTL的鉴定与统计分析
为了了解个体之间的遗传变异如何以细胞类型特异性的方式影响基因表达,研究测试了基因两端(包括基因体)1 Mb 顺式区域内的 SNP 基因型与 14 种细胞类型中基因表达之间的关联。
这种方法可识别每种细胞类型的 eQTL,进而能够评估遗传对基因表达的影响在 PBMC 之间的共享程度。顺式区域内的多个 SNP 可能与基因表达相关,因为连锁不平衡引起的基因型与与基因表达水平相关的众多独立位点之间存在相关性。为了区分这些情况,研究对每个已识别的 eQTL 进行了条件分析,将领先的 eQTL SNP(eSNP)作为后续分析中的条件协变量。
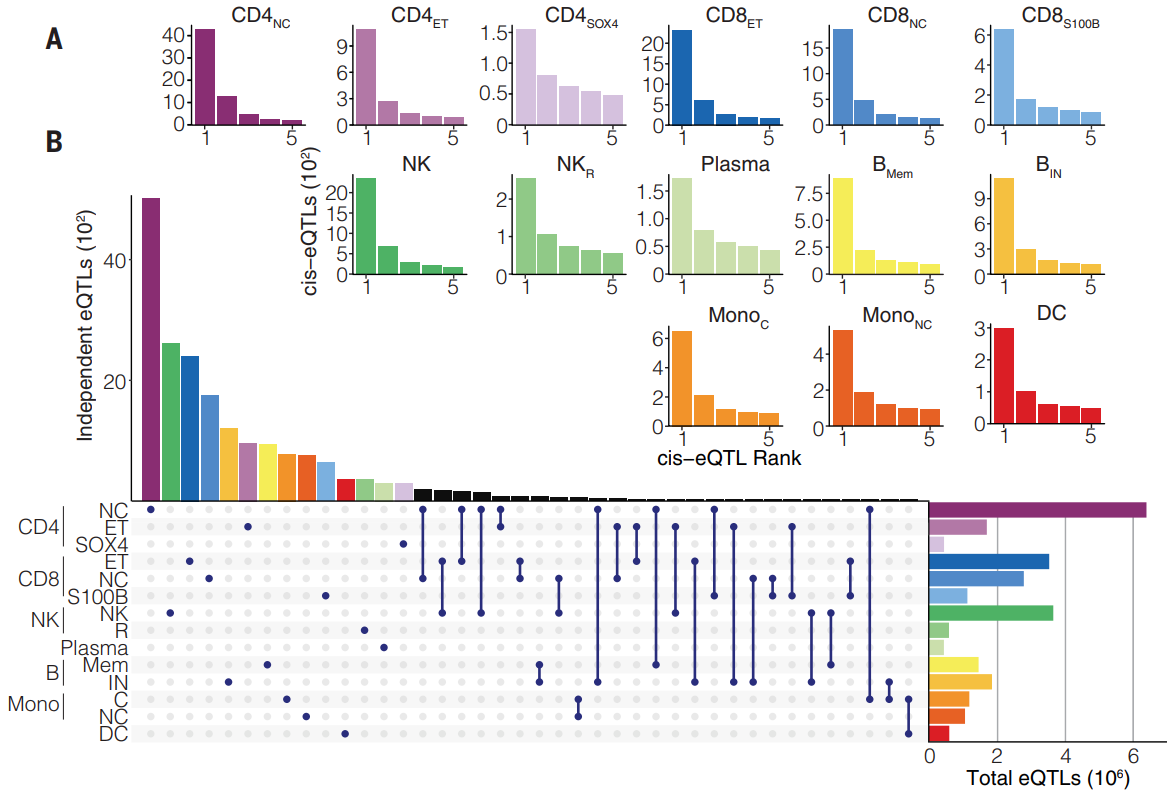
(A)展现五轮条件 eQTL 分析结果,在每轮分析中,在研究范围的 FDR 下检测到的 cis-eQTL 数量小于 0.05。研究对 39.7% 的测试基因鉴定了 26,597 个 eQTL,其中 16,597 个(eSNP1)来自第一轮分析,另外 10,000 个(eSNP2 至 eSNP5)来自四轮条件测试。
(B)独立 eQTL 的数量因细胞类型而异,在 CD4NC 细胞中鉴定出 6473 个,在浆细胞中鉴定出 399 个。
条件性eQTL分析与独立基因座
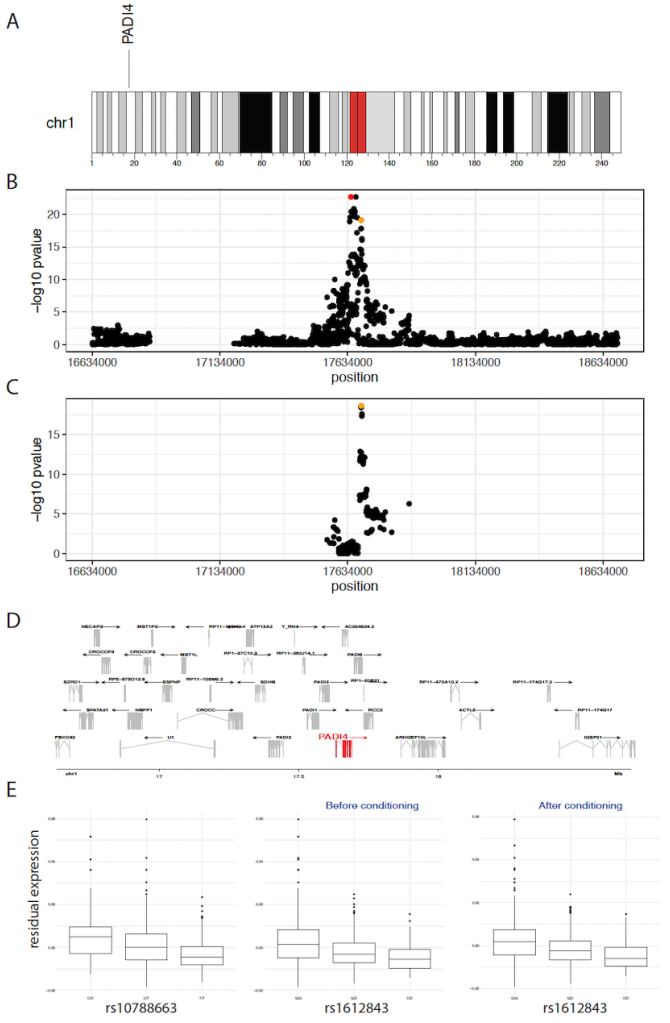
这些条件性 eQTL 识别了顺式区域内有多个独立基因座的情况,这些基因座的基因型与基因的表达水平相关。例如,在 CD4NC 细胞中,研究确定了 PADI4 的主要 eQTL。PADI4 在基因和细胞水平上都与类风湿性关节炎 (RA) 的发病机制有关。该 eQTL 的top eSNP1 是 rs10788663,其中每个 T 等位基因拷贝导致每个细胞平均减少 0.28 个 mRNA 转录分子。
在随后的一轮条件分析中,研究将 rs10788663 作为协变量,并再次测试整个顺式区域的关联,确定了一个由top eSNP2 rs1612843 标记的次要独立 eQTL。平均而言,携带每个 rs1612843 C 等位基因拷贝的个体每个细胞的 mRNA 转录分子减少 0.24 个。
rs10788663 位于第一个内含子中,而 rs1612843 位于 PADI4 的第 15 和 16 外显子之间的内含子中,这表明独立的转录因子可能调节多个独立位点,并且是调节 PADI4 表达所必需的。在 OneK1K 队列中,rs10788663 和 rs1612843 之间的连锁不平衡为 0.0678,这进一步证明多个独立的 eQTL 影响 CD4NC 细胞中 PADI4 的表达。事实上,在确认两个独立基因座的预期附加效应后,研究观察到携带纯合 T/T 和 C/C 的个体与携带 C/C 和 G/G 的个体相比,rs10788663 和 rs1612843 的平均差异为 1.04 个 mRNA 转录本。
细胞类型特异性与普遍性探索

(A)展示不同细胞类型之间零p值的比例(π1)。颜色越深代表零p值比例越高,这意味着在这些细胞类型对之间,基因表达的差异越显著。
(B)展示了BIN细胞类型与其他单个细胞类型之间的p值分布。每个直方图表示BIN与其他某个特定细胞类型的比较。水平虚线表示对零p值比例(π1)的高度估计。直方图中p值集中在靠近0的位置表明这些细胞类型间存在显著差异,而p值均匀分布则表示差异不显著。
在一种细胞类型中鉴定出的顺式 eQTL 在另一种细胞类型中显示其检验统计量膨胀的平均比例为 p1 = 0.53(0.19 至 0.96)。这表明,如果样本量较大,目前在一种细胞类型中鉴定出的顺式 eQTL 应该在一种或多种其他细胞类型中达到研究范围内的显著性。然而,它们的等位基因效应的大小可能因细胞类型而异。
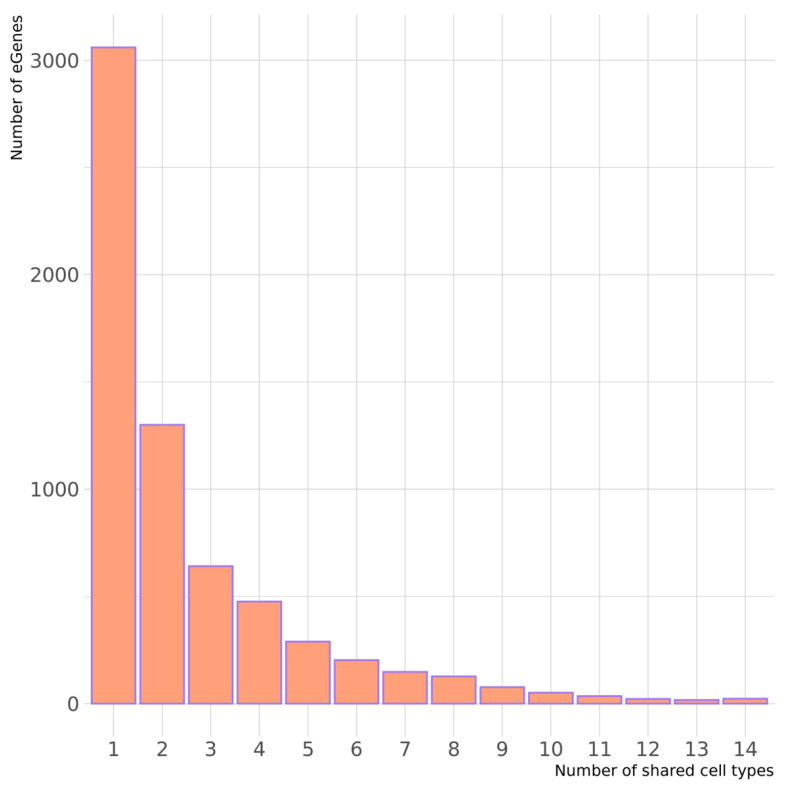
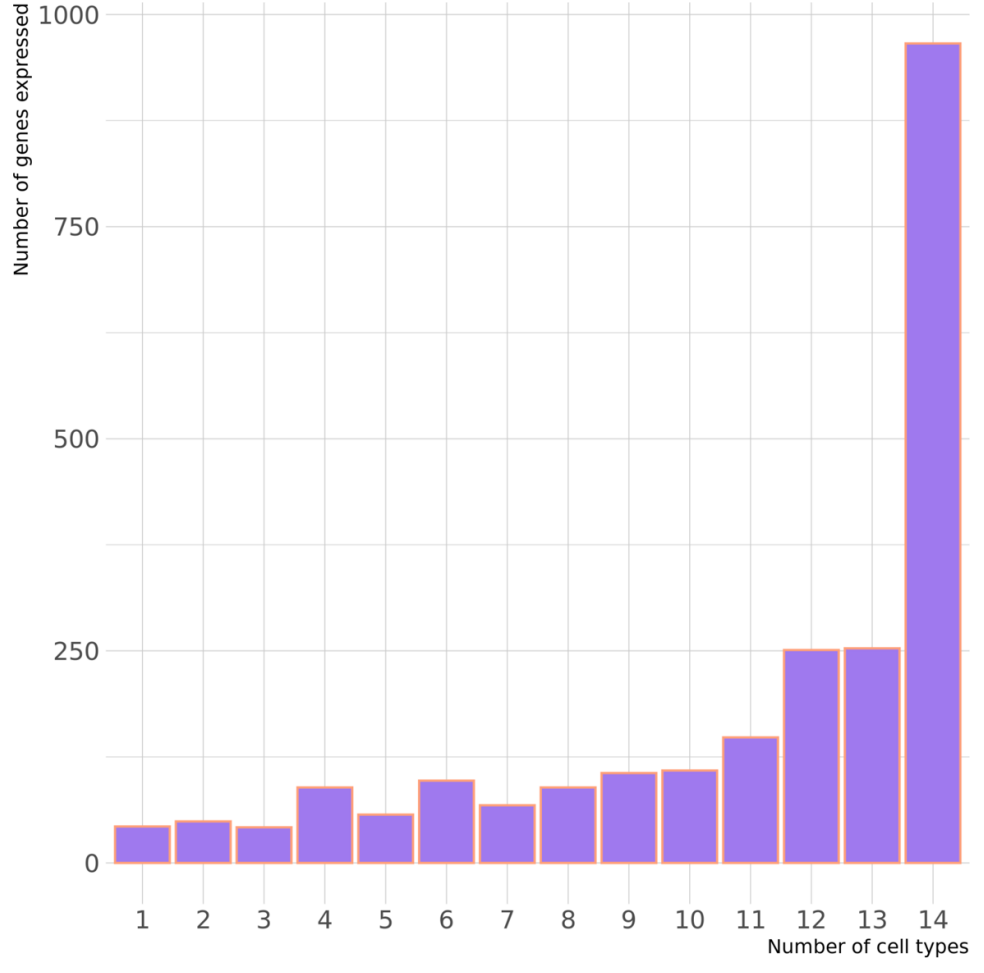
上图显示在一种细胞类型中鉴定的 eGenes 数量,以及在两种或多种细胞类型中共享的数量。对于仅在一种细胞类型中鉴定出的 3060 个 eQTL(eGenes)基因,并没有发现任何证据表明它们在其他细胞类型中存在等位基因效应,这表明这些确实是细胞类型特异性的。
细胞类型特异性 eQTL 的观察有多种可能的解释:该基因可能仅在一种细胞类型中可检测到表达,在多种细胞类型中检测 eQTL 的统计能力可能较低,或者跨细胞类型存在真正的调控异质性。
对细胞类型特异性eQTL的进一步分析
为了评估这些不同的情况,研究对每个具有至少一个 eQTL 的基因(eGene n = 6469)进行了一系列分析。这些 eGene 中只有 43 个(0.7%)在一种细胞类型中表达。其余 6426 个在多种细胞类型中表达,除了具有显著 eQTL 的细胞类型外,这些基因平均在 11 种细胞类型中表达。
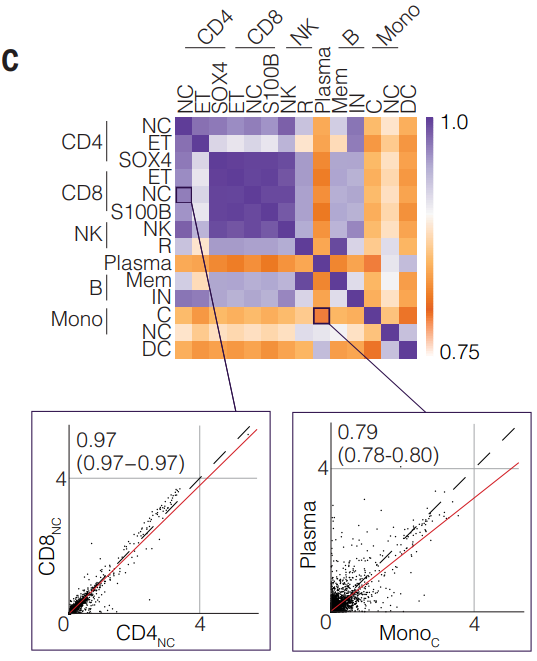
当测试一对细胞类型之间这 6426 个 eGene 中每个基因的表达水平的相关性时,发现共表达的整体一致性很高。而且几种细胞类型之间 eGene 表达水平的平均相关性模式遵循造血谱系关系,反映了它们在免疫系统中的紧密关联。
因此,大多数eGenes并不是因为这些基因仅在一种细胞类型中表达(即细胞类型特异性表达),而是因为这些基因的调控因子(如转录因子)在不同细胞类型中有特异性表达。这表明调控因子的细胞类型特异性可能是导致观察到的基因表达差异的主要原因。
确定这些 eGene 在多种细胞类型中表达后,研究接下来试图评估观察到细胞类型特异性 eQTL 是否是由于在多种细胞类型中检测等位基因效应的统计能力低。
作者实施了一个经验框架来测试非显著细胞类型中 eGene 等位基因效应的检验统计量的等级。在几乎所有情况下,都观察到跨细胞类型的检验统计量没有或极少富集,这表明在大多数情况下,细胞类型特异性 eQTL 是由特定的细胞调节机制引起的。在确定明显富集的情况下,存在与造血谱系密切相关的细胞类型。然而,对于大多数 eGene,没有在检验统计量中确定富集,这再次表明效应是细胞类型特异性的。这些结果共同表明,为 2367 个 eGene 确定的大多数 eQTL 仅针对单一细胞类型。
多种细胞类型中eQTL的共享性分析
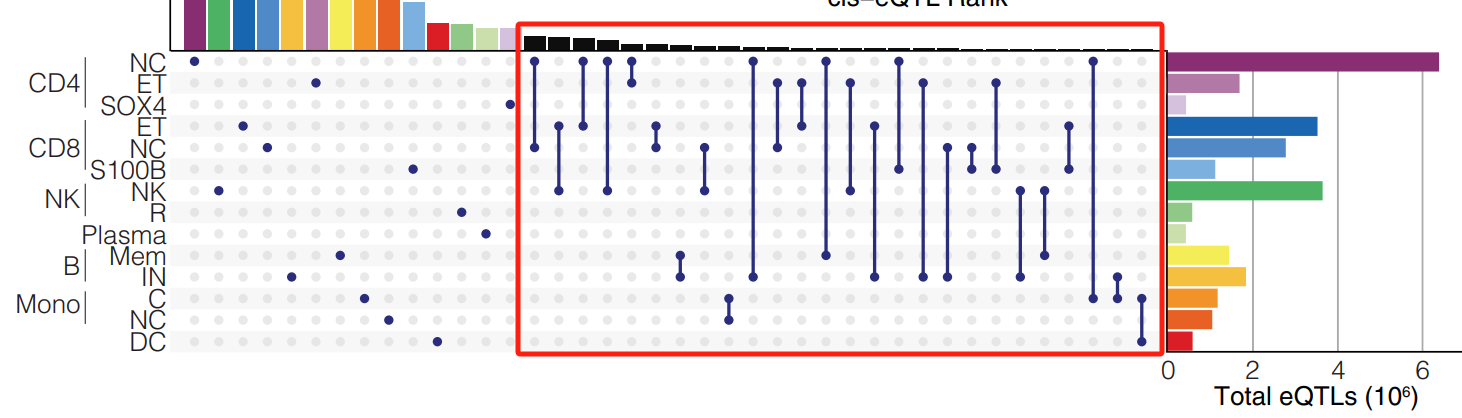
对于剩余的 4102 个 eGene,在两种或更多种细胞类型中总共鉴定了 14,230 个 eQTL,尽管对于其中 1386 个 eGene,研究在细胞类型之间观察到了不同的 eSNP。
在这种情况下,一种假设是相同的变体是多种细胞类型中 eQTL 的基础,top eSNP 的差异是由于基因表达模式的变化。另一种假设是 eQTL 来自影响不同细胞类型表达的独立变体。
为了检验这些假设,研究执行了回归策略,以评估在回归出另一种细胞类型的 eSNP 的影响后 eSNP 的检验统计量的变化,在不同细胞类型中鉴定出不同的顶级 eSNP。如果 eSNP 标记该基因的相同因果变体或彼此处于连锁不平衡状态,则原始 eSNP 的等位基因效应大小将在条件分析中减小。类似地,如果它们标记独立变体,等位基因效应将保持相对不变。
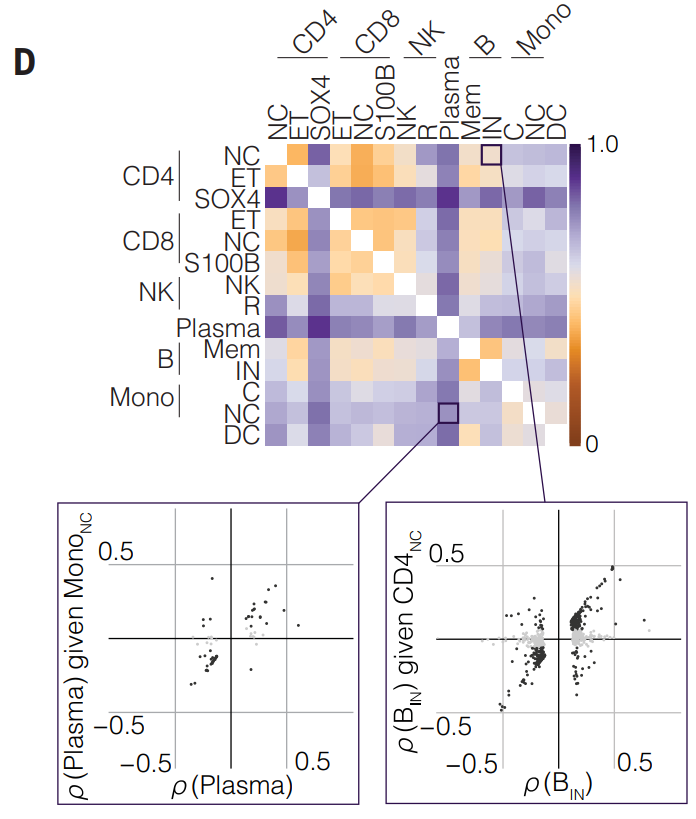
研究通过将来自第一种细胞类型的lead eSNP 调节到来自第二种细胞类型的lead eSNP 上,对每对细胞类型(182 对)测试每个 eGene 是否被两个不同的变体标记。其中热图显示了不同细胞类型之间的等位基因效应的相关性,底部的两幅散点图展示了对特定细胞类型的lead eSNP(导出的显著SNP)进行调节后的等位基因效应变化。
- 显著变化: 如果散点图中有点远离对角线,这表明调节前后等位基因效应发生了显著变化,意味着两种细胞类型的eQTL可能是相同的,或是由于eSNPs之间的连锁不平衡。
- 缺乏变化: 如果变化不大(即点集中在对角线附近),则提供了eQTL独立性的证据,即在不同细胞类型中,基因表达受不同的调控因子影响。
虽然大多数淋巴免疫细胞 eQTL 在调节后的相关系数发生了相当大的变化,但在髓系免疫细胞中,eQTL 相关系数保持相似(这部分附图较多,暂且不放上来)。这一发现表明,与髓系细胞相比,淋巴细胞类型更有可能在细胞类型之间共享基因表达的遗传控制。
4.细胞类型特异性染色质可及性是一定比例细胞类型特异性顺式 eQTL 的基础
为了探索顺式 eQTL 背后的功能调控,我们测试了 eSNP 位置与 8876 个细胞的转座酶可及染色质测序 (scATAC-seq) 数据单细胞分析生成的开放染色质区域的重叠情况。
将细胞分为 14 种细胞类型,并针对每种有 5 个以上分类细胞的细胞类型调用开放染色质峰。这种过滤保留了 11 种细胞类型。
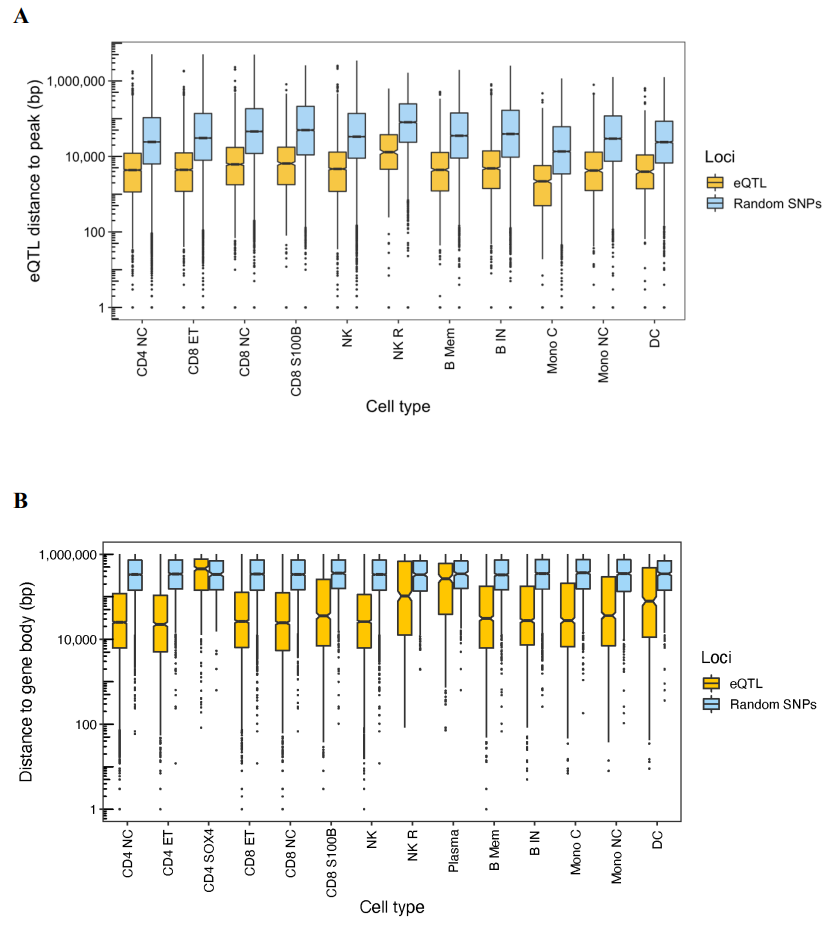
scATAC-seq平均为每个细胞类型确定了 52,048 个开放染色质区域的峰,eSNP 与最近峰之间的平均距离范围为 7485 至 31,383 个碱基对。为了确定顺式 eQTL 的位置是否明显更接近开放染色质区域,研究比较了顺式 eQTL 之间的距离。研究使用了bootstrapping方法来检验这种位置关系的显著性,该方法创建一个对照分布:研究者随机选择了SNP,这些SNP的位置分布与实际eSNP到开放染色质峰的距离分布相同。通过比较实际eQTL的位置与这个随机分布,研究者可以确定eQTL是否倾向于更靠近开放染色质区域。
(A-B)研究观察到除 CD4SOX4 细胞外所有细胞类型的顺式 eQTL 距离之间存在显著差异 (FDR) < 0.05。所以,细胞类型特异性染色质可及性可能导致等位基因对细胞类型之间基因表达的影响发生变化。
5.单细胞eQTL在多种族队列和bulk eQTL研究中得到了验证
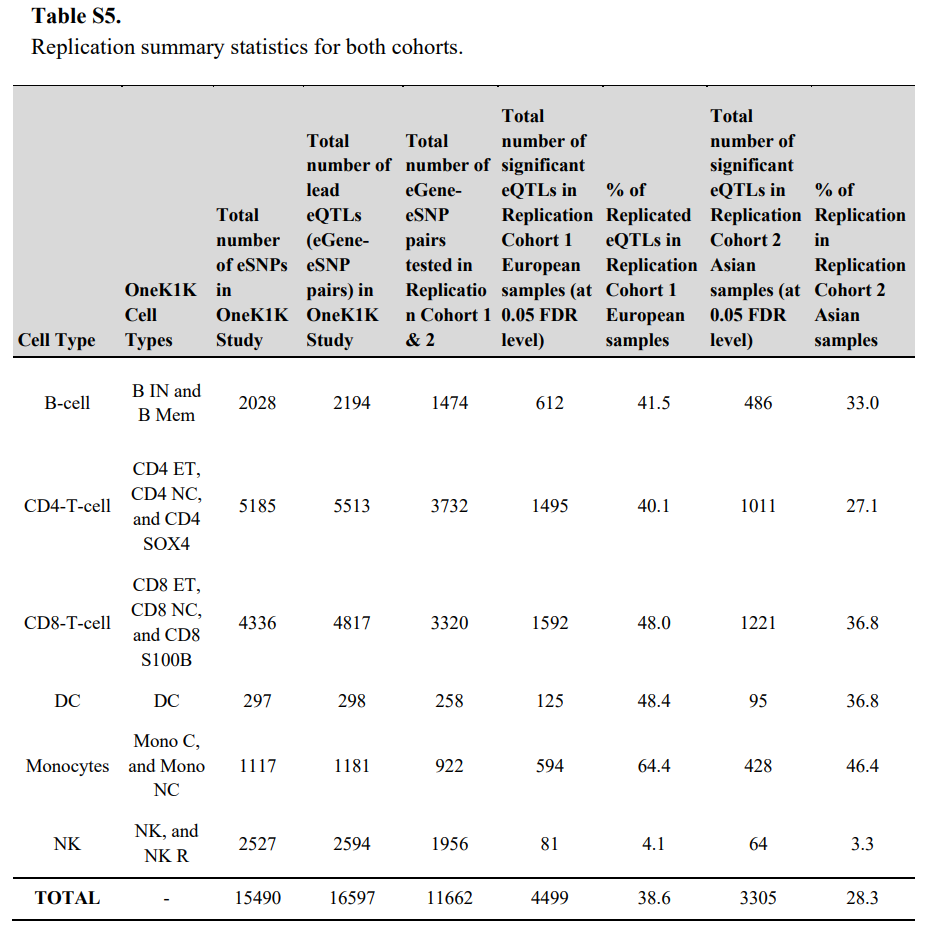
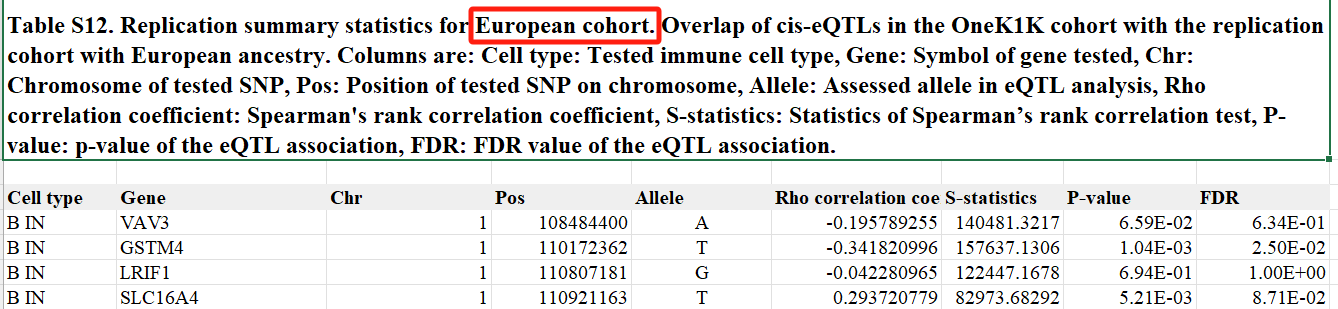
为了验证细胞特异性 eQTL 发现,研究在两个独立的欧洲和亚洲血统队列中验证了主要 eSNP 结果,这两个队列分别由 113 人和 89 人组成。在 16,597 对 eSNP1-eGene 对中,10,071 对的次要等位基因频率在两个队列中均大于 0.05。其中,欧洲队列中有 3198 对(26%)和亚洲队列中有 2243 对(22%)在 5% 的 FDR 阈值下得以验证。
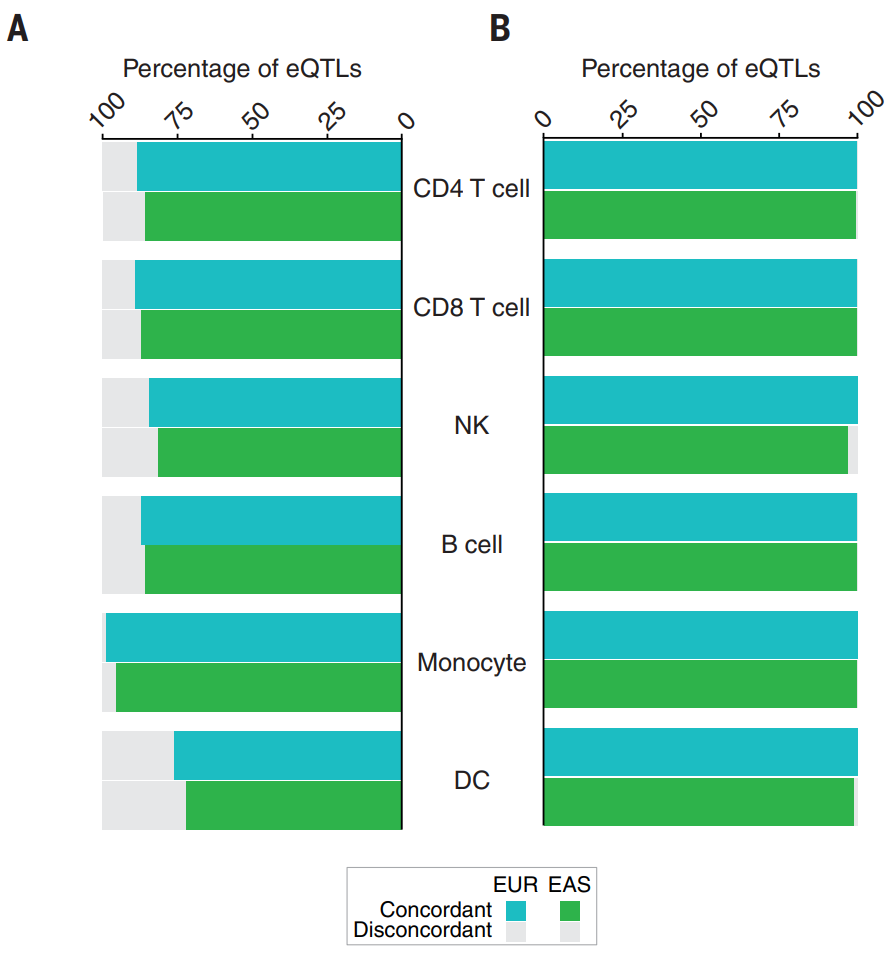
实际上在观测队列和验证队列中样本量相等的假设下校正 FDR 分布,欧洲队列和亚洲队列的复制率分别为 87% 和 78%。同样,所有测试基因座的等位基因方向一致性在欧洲队列中为 76.0% 至 98.1%,在亚洲队列中为 72.2% 至 95.4%。对于 FDR 小于 0.05 的 eQTL,这种一致性分别增加到 99.3% 至 100% 和 96.9% 至 99.8%。不同队列之间复制率的差异可能反映了不同人群之间 eSNP 等位基因频率的差异。结果表明,细胞类型特异性 eQTL 可能在很大程度上在人群之间共享。
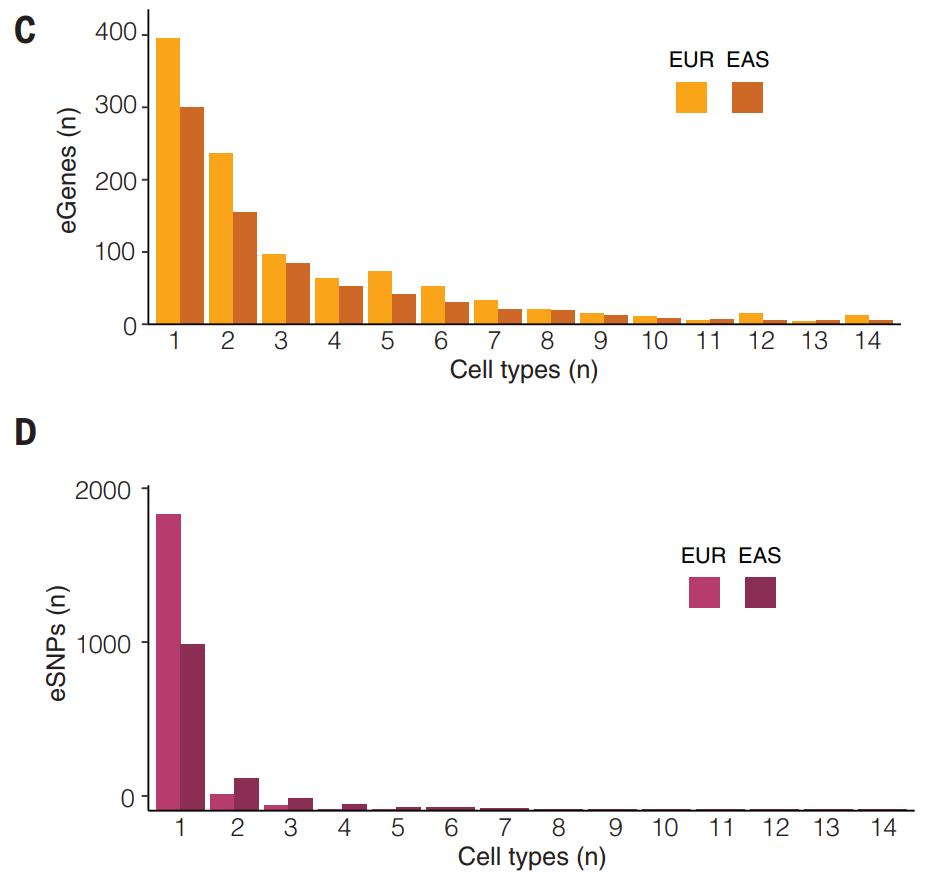
(C-D)OneK1K eQTL 的发现在验证队列中的所有细胞类型中进行了测试。当 FDR 小于 0.05 时,验证的 eQTL 和 eGene 主要在单一细胞类型中识别,这为基因座对 PBMC 中基因表达的细胞类型特异性影响提供了进一步的证据。
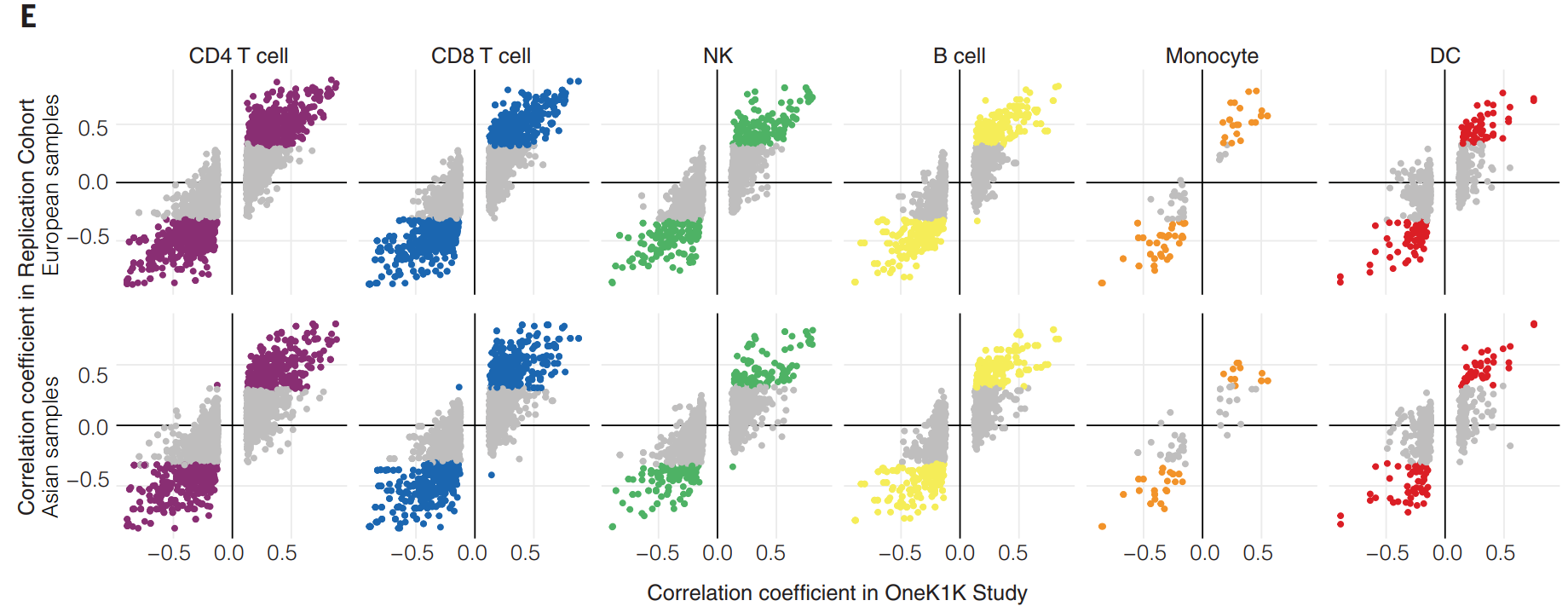
(E)显示了 OneK1K 和验证队列之间的相关系数一致性,包括欧洲和亚洲样本。横轴表示在OneK1K研究中的相关系数,纵轴表示在复验证据中的相关系数。相关系数大小越接近1或-1,表示相关性越强;接近0则表示无相关性;相关系数的正负值表示等位基因效应相对于参考等位基因的方向。一致的正负方向表示在不同人群中该eQTL的效应方向一致。可见不同人群中的eQTL效应具有一致性,eQTL在不同细胞类型中的表现不同。

研究能够验证在 eQTL-Gen Consortium 和 GTEx Consortium 的血液样本bulk RNA 测序研究中识别的 62.5% 和 40.4% 的顺式 eQTL。
6.识别 B 细胞景观中的动态 eQTL 等位基因效应

(A-C)文章分析了 eQTL 在未成熟和幼稚 B (BIN) 细胞到记忆 B (BMem) 细胞的拟时序景观中的动态效应。根据细胞在轨迹上的相对位置,将细胞分为六个部分 (Q1 至 Q6)。Biomarkers的可视化揭示了从 BIN (Q1) 到 BMem 细胞 (Q6) 的衍生轨迹的分级变化。
TCL1A 和 IL4R 在幼稚 B 细胞中高度表达,并且在向 BMem 细胞的转变过程中被发现下调。相反,随着细胞转变为记忆型,典型 BMem 细胞标记物 CD27 的表达增加。 IgJ 表达是免疫球蛋白 M (IgM) 和 IgA 产生的标志,在较高分位数中上调,表明它们含有准备分化成浆细胞的细胞。
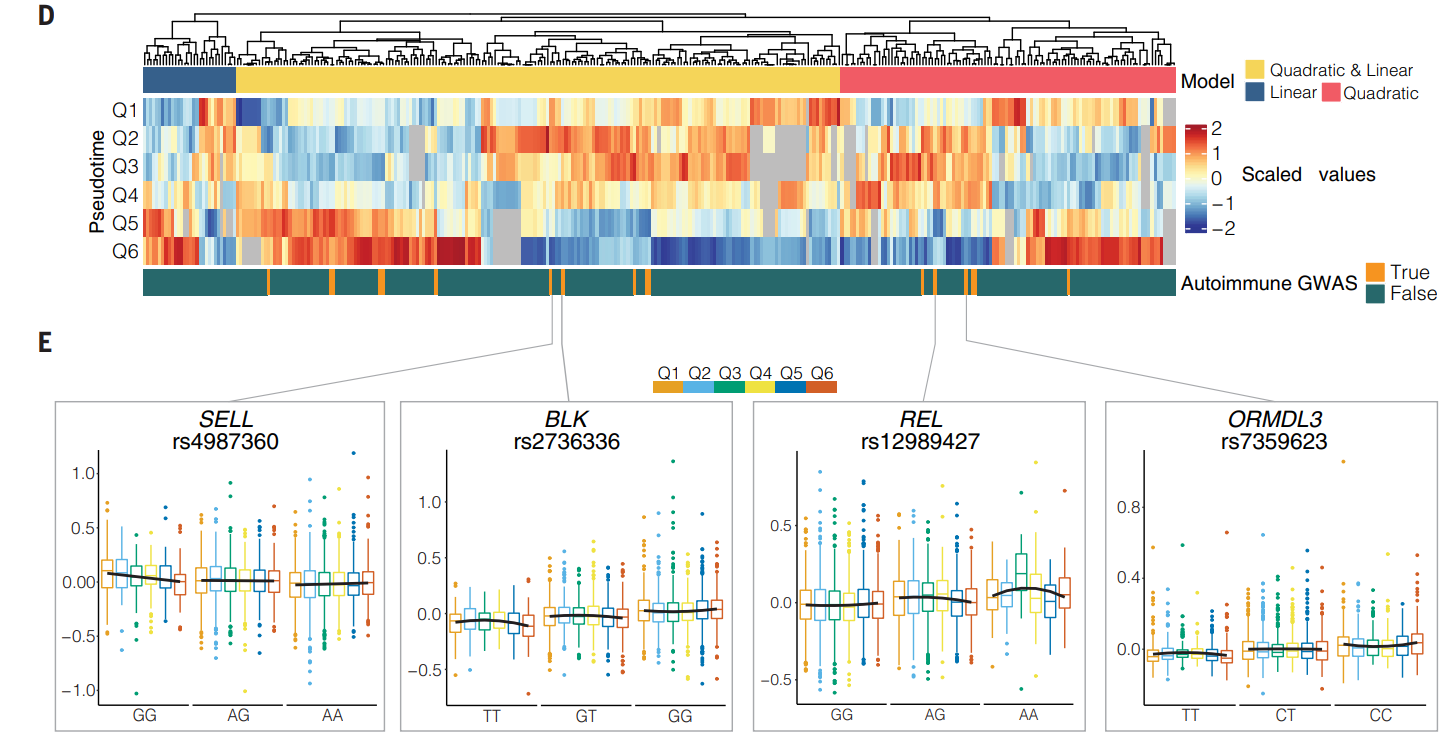
研究尝试去确定 eQTL 等位基因效应在幼稚到记忆性 B 细胞转变轨迹中表现出线性或非线性变化的情况。通过使用线性和二次模型测试基因型和分位数等级之间的相互作用来确定动态 B 细胞 eQTL。
(D)在 BIN 和 BMem 细胞中鉴定的 3074 个顺式 eQTL 中,1988 个在至少三个拟时序分位数中表达并测试了动态效应。确定了其中 333 个的等位基因效应在整个轨迹中发生了显著变化(FDR < 0.05)。底部的绿色条形表示显示已知处于高度连锁不平衡 (R2 > 0.8) 的 SNP 与通过自身免疫性疾病的 GWAS 鉴定的变异。eGene未在给定分位数中表达的情况以灰色显示。
(E)许多具有动态 eQTL 效应的基因在微调 B 细胞迁移、激活、存活或功能方面发挥作用。
- SELL :SELL 参与整合素介导的组织迁移和组织内迁移。B 细胞迁移到生发中心并在生发中心内组织是产生适当记忆和体液输出的关键组成部分。内含子变体 rs4987360-G 对 SELL 表达的等位基因效应在未成熟细胞中最大,在随后的每个分位数中减小。
- BLK:对于影响 Src 家族酪氨酸激酶 B 淋巴细胞激酶 (BLK) 表达的 SNP,发现了相反的趋势,BLK 是一种负责调节 B 细胞受体下游信号传导幅度的基因。rs2736336 和 rs2409780 均在 Q5 和 Q6 中表现出最大的等位基因效应。而且 BLK 启动子中的变体 rs2736336 与系统性红斑狼疮 (SLE) 相关,而内含子变异 rs2409780 与 SLE 和 RA 相关的变异处于高度连锁不平衡(判定系数 (R2) = 0.99,连锁不平衡系数 (D′) = 0.99)。
- REL:B 细胞表面受体下游信号传导并影响随后 B 细胞增殖和存活的基因是 c-Rel,由转录因子 REL 编码。rs12989427 与与 SLE 相关的变体处于高度连锁不平衡(R2 = 0.88,D′ = 0.98),等位基因效应遵循非线性关系,在 B 细胞轨迹的中间点达到峰值。
- ORMDL3:ORMDL3通过抑制细胞凋亡和促进自噬来促进成熟B细胞存活。rs7359623和rs8067378与风险变异处于高度连锁不平衡状态(R2>0.8,D′>0.9),与一系列自身免疫疾病有关,并且在整个轨迹中对B细胞中的ORMDL3具有动态eQTL效应。
7.遗传变异以细胞类型特异性的方式控制转录调控,从而调节免疫途径
尽管人们普遍认为免疫调节在个体之间是可变的,但导致这种变化的因素却知之甚少。通过选择文献中描述的影响免疫调节的基因,研究展示了基因位点如何以细胞类型特异性的方式导致免疫调节基因表达的变化。
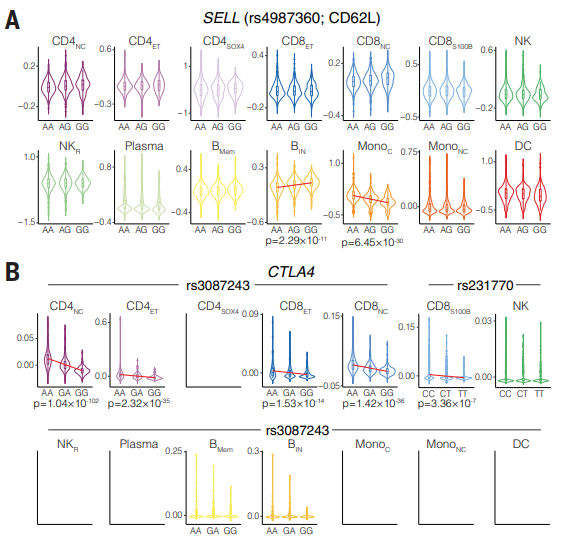
(A)血液和淋巴结之间的白细胞再循环是免疫系统的基本特性,它依赖于由 SELL 基因编码的淋巴结归巢受体 CD62L(L-选择素)。研究发现在 rs4987360 的影响下,先天免疫系统和适应性免疫系统之间 SELL mRNA 的反向调节,rs4987360 是与单核细胞血细胞计数相关的 rs4987353(R2 = 1,D′ = 1)连锁不平衡中的常见多态性。 rs4987360- G 等位基因降低了 MonoC 中的 SELL mRNA,但增加了 BIN 细胞中的 SELL mRNA,说明单个遗传等位基因如何通过不同的细胞类型来影响基因表达。刚好第六部分的动态 eQTL 分析已经发现 rs4987360 的等位基因效应在 B 细胞状态景观中各不相同。rs4987360 关联在来自 eQTL-Gen 和 GTEx 的大量 RNA 测序 eQTL 数据中复制,并在大量 B 细胞和单核细胞中具有相反方向的等位基因效应。
(B)CTLA4 是一种对基因量敏感的 T 细胞必需抑制受体。与 SELL 相反,CTLA4 下游的 rs3087243-G 等位基因与 1 型糖尿病 (T1DM) 和 RA 易感性相关,该等位基因以相同的等位基因方向作用于多种细胞类型,通过降低四种 T 细胞亚群中的 CTLA4 mRNA 表达。多态性 rs231770 距离 rs3087243 不到 10 kb,但处于连锁平衡 (R2 = 0.5)。rs231770-T 同样与表达 S100B (CD8S100B) T 细胞的 CD8+ T 细胞中 CTLA4 mRNA 表达降低有关,并且与自身免疫性疾病重症肌无力有关。
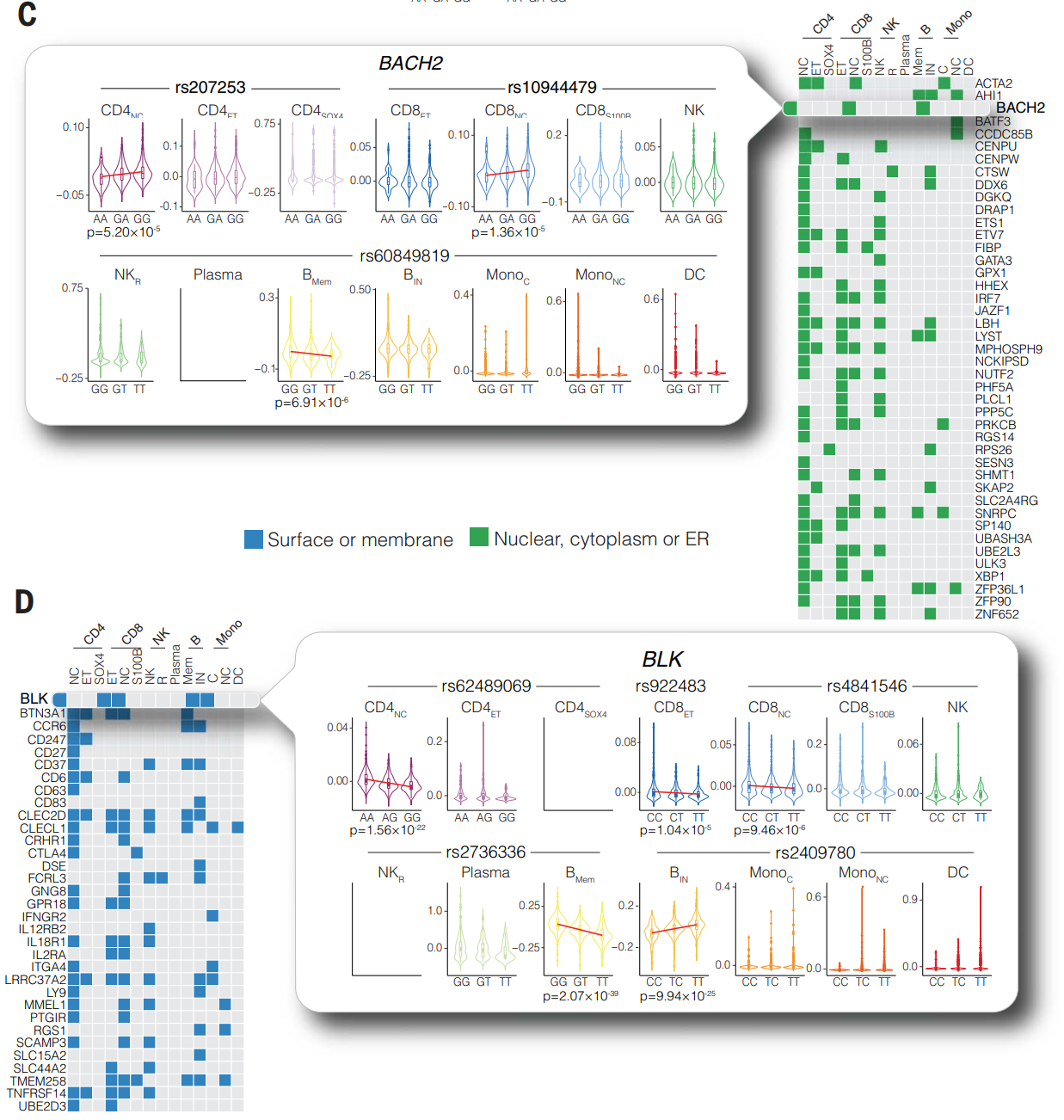
通过关注与自身免疫性疾病有关的基因,研究评估了等位基因效应在不同细胞类型中的差异,重点关注编码膜、核、细胞质或内质网 (ER) 蛋白的基因。
(C)BACH2:一种参与分化记忆 B 细胞和 T 细胞的必需转录因子。研究确定了 rs10944479,它对 CD8NC 细胞中的 BACH2 具有 eQTL 效应。研究在 CD4NC、CD8NC 和 BMem 细胞中鉴定了 BACH2 的 eQTL,尽管每种细胞类型中控制表达的基因座彼此独立(R2 = 0 至 0.11)。研究证明 rs60849819-T 与 BMem 细胞中 T 等位基因纯合的个体中 BACH2 的显著下调有关,而 rs207253-A 在 CD4NC 细胞中具有类似的效果。
(D)BLK:五个 eSNP 与 CD4NC、具有效应记忆表型的 CD8+ T 细胞 (CD8ET)、CD8NC、BMem 和 BIN 细胞中的 BLK 表达有关,并且与 RA、SLE、干燥综合征和系统性硬皮病有关。其中一个基因座 rs2736336 导致 BMem 细胞中 BLK 的差异表达。rs2736336 与 SLE 有关,携带自身免疫风险等位基因的副本与 B 细胞的过度活化有关,T 细胞共刺激能力增强。结果表明 rs2736336 的等位基因变异导致个体间维持 B 淋巴细胞耐受性的差异。Src 家族酪氨酸激酶(例如 BLK)是信号通路的重要组成部分,作用于抗原受体的下游,并决定细胞因抗原参与而接收的信号强度。
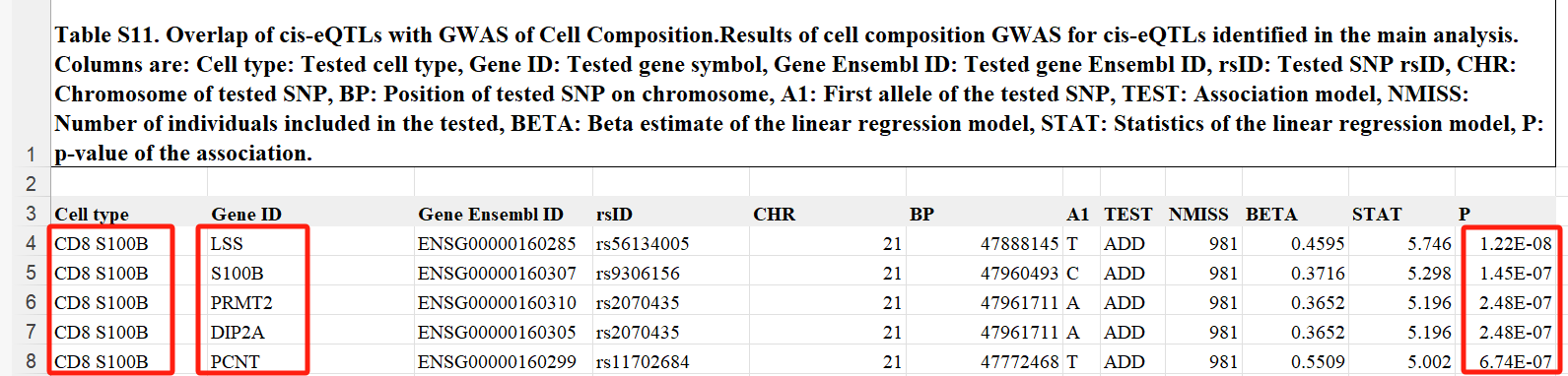
最后,研究评估 eQTL 对 OneK1K 队列中细胞组成的影响。对于每个 eSNP1,测试了个体基因型与细胞类型比例之间的关联。在研究范围显著性阈值(p < 3.0 × 10−6)下,确定了五个关联,所有这些关联都会影响 CD8S100B 细胞的比例。eGenes(LSS、S100B、PRMT2、DIP2A)和 PCNT 均位于 21q22 染色体上的 1 Mb 区域内,并且 SNP 彼此之间存在中等至高度的连锁不平衡(R2 = 0.31 至 0.97),这表明单个变体会影响 CD8S100B 细胞的比例。(部分展示表S11)
8.细胞类型特异性 trans-eQTL 的鉴定表明远端基因组调控具有高度的细胞类型特异性
研究进行了 trans-eQTL 分析,测试了每个 cis-eQTL 中的top eSNP 与所有其他基因的基因表达水平,但不包括 cis-eGene 和主要组织相容性复合体 (MHC) 基因座 ±2 Mb 范围内的基因。在研究范围内的 FDR 为 0.01 时,确定了 990 个 trans-eQTL(每个 cis-eSNP 中位数为 1 个)。
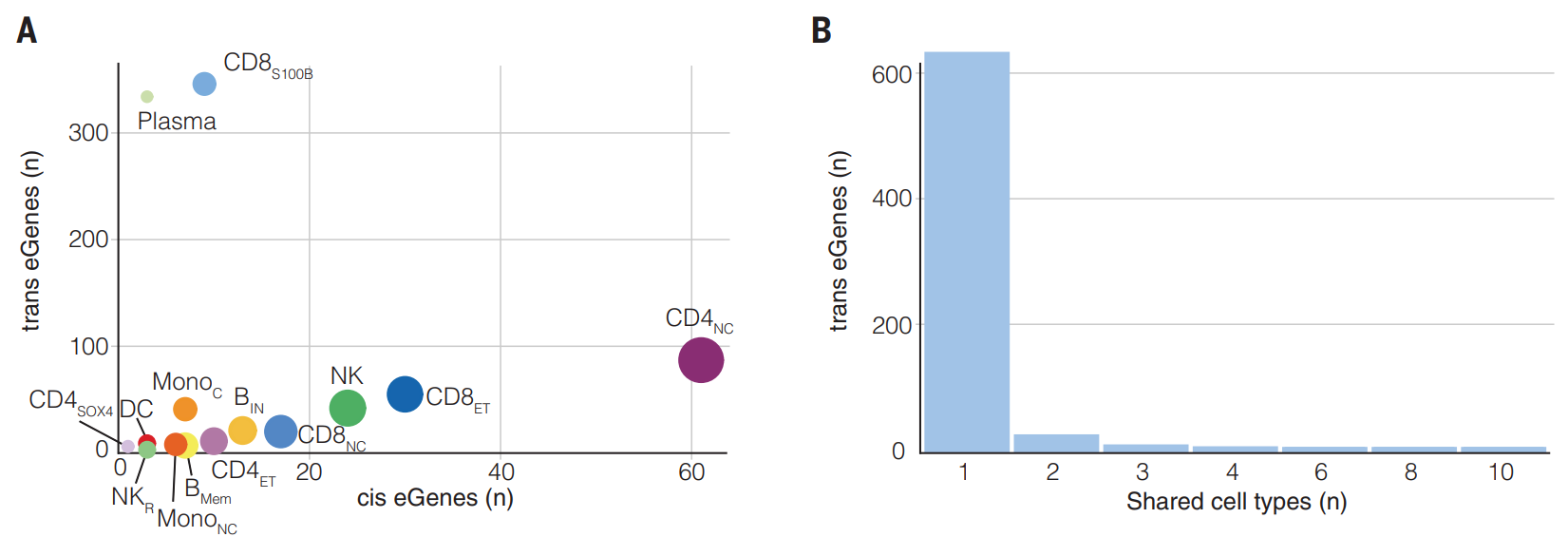
(A-B)在每种细胞类型中确定的 trans-eGene 数量与 cis-eQTL 总数弱相关(Spearman r = 0.37)。与 cis-eGene 相比,大多数 trans-eGene 特定于一种细胞类型,并且没有一种在细胞类型中普遍存在。
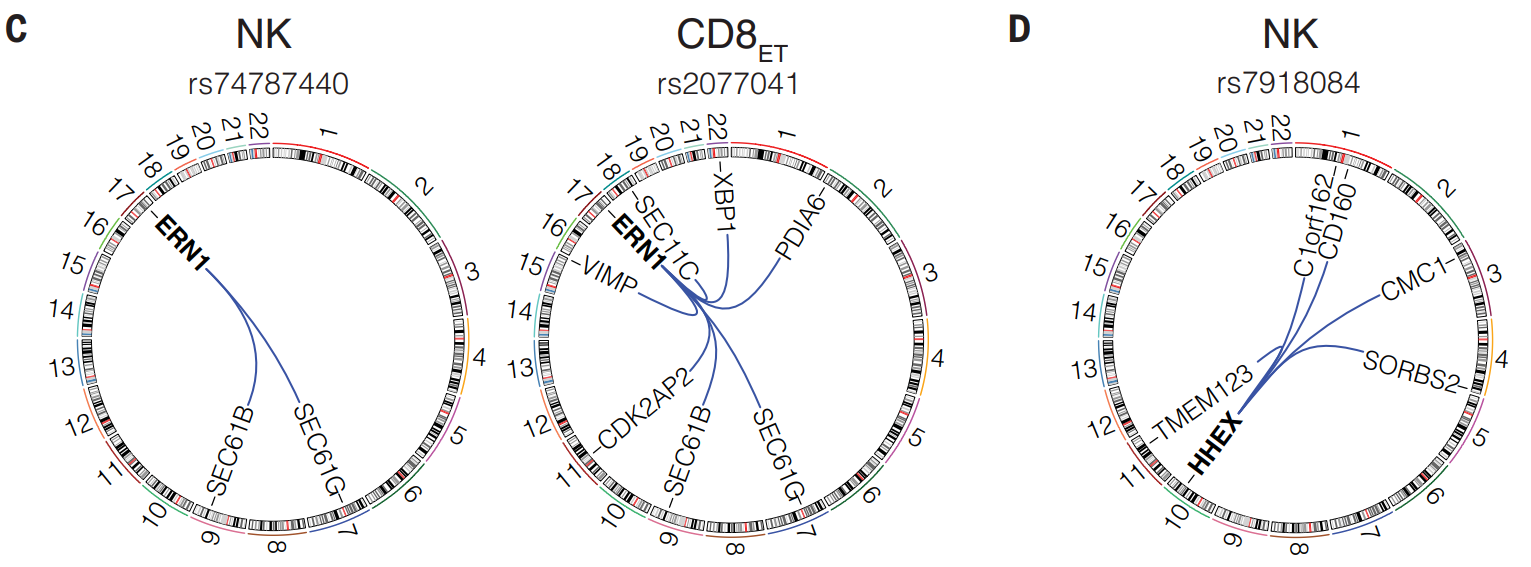
(C)研究确定了 630 种细胞类型特异性 trans-eQTL 效应。例如,rs2077041 对 CD8ET 细胞中的 ERN1 表达具有顺式效应,C 等位基因会降低表达。该基因座在七个 trans-eGenes 中具有相同的等位基因方向效应。携带 rs2077041 C 等位基因拷贝的个体 XBP1 和 SEC61G、SEC61B 和 SEC11C 下调,这些基因参与了蛋白质跨 ER 膜的转运、信号肽去除和整合。rs74787440 还被发现对自然杀伤 (NK) 细胞中的 ERN1 表达具有显著的顺式效应。然而,这种相同的变异对 SEC61G 和 SEC61B 具有反式效应,但对与 rs2077041 相关的其他基因没有影响。
(D)在已确定的自身免疫风险基因座上发现了反式 eQTL,包括 rs7918084-T,它是 NK 细胞中 HHEX 的顺式 eQTL,与特应性哮喘和外周血嗜酸性粒细胞计数有关。HHEX 结合并抑制促凋亡因子 BIM,从而增加 NK 细胞的数量。

在 NK 细胞中,rs7918084-T 在四条染色体上产生反式 eQTL 效应,降低 CD160、CMC1、SORBS2、TMEM123 和 C1orf162 的表达。 CD160 是一种刺激性受体,在促进 NK 细胞干扰素-g (IFN-g) 产生方面发挥重要作用。
9.遗传风险变异与单细胞顺式 eQTL 的共定位确定了自身免疫性疾病的细胞类型特异性机制
研究使用共定位和孟德尔随机化方法测试了每种细胞类型的顺式 eQTL(使用 eSNP1)在 2335 个性状相关 SNP 中的富集程度。共定位发现 19% 的顺式 eQTL 具有与 GWAS 风险变体相同的因果基因座。
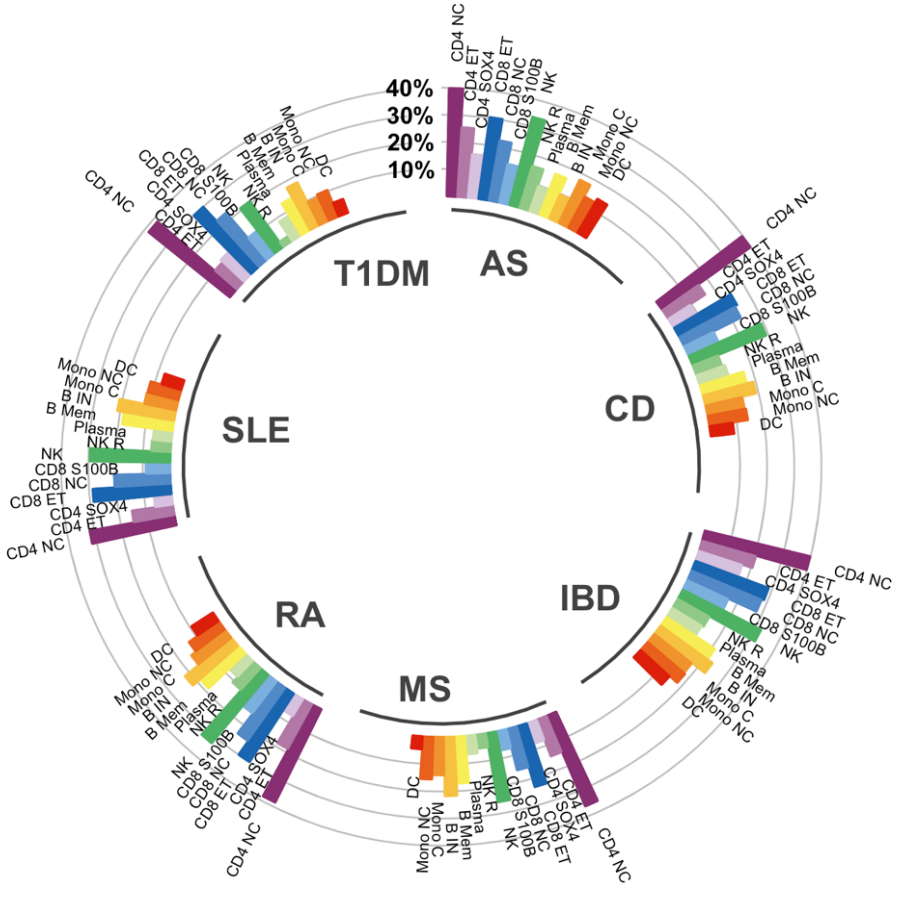
图中展示 GWAS 确定的七种自身免疫性疾病(MS=多发性硬化症、RA=类风湿性关节炎、SLE=系统性红斑狼疮、AS=强直性脊柱炎、CD=克罗恩病、T1DM=1 型糖尿病、IBD=炎症性肠病)的风险变异之间重叠细胞类型特异性 eQTL 的百分比。CD4NC 和 NK 细胞中的重叠最高。类似地,在 NK 募集 (NKR) 细胞中,炎症性肠病 (IBD)、RA、强直性脊柱炎 (AS) 和克罗恩病 (CD) 的重叠富集程度较高,而多发性硬化症 (MS)、SLE 和 T1DM 的重叠富集程度较低。这些结果凸显了这些常见自身免疫性疾病的遗传风险多基因效应在细胞水平上发挥作用的复杂性。
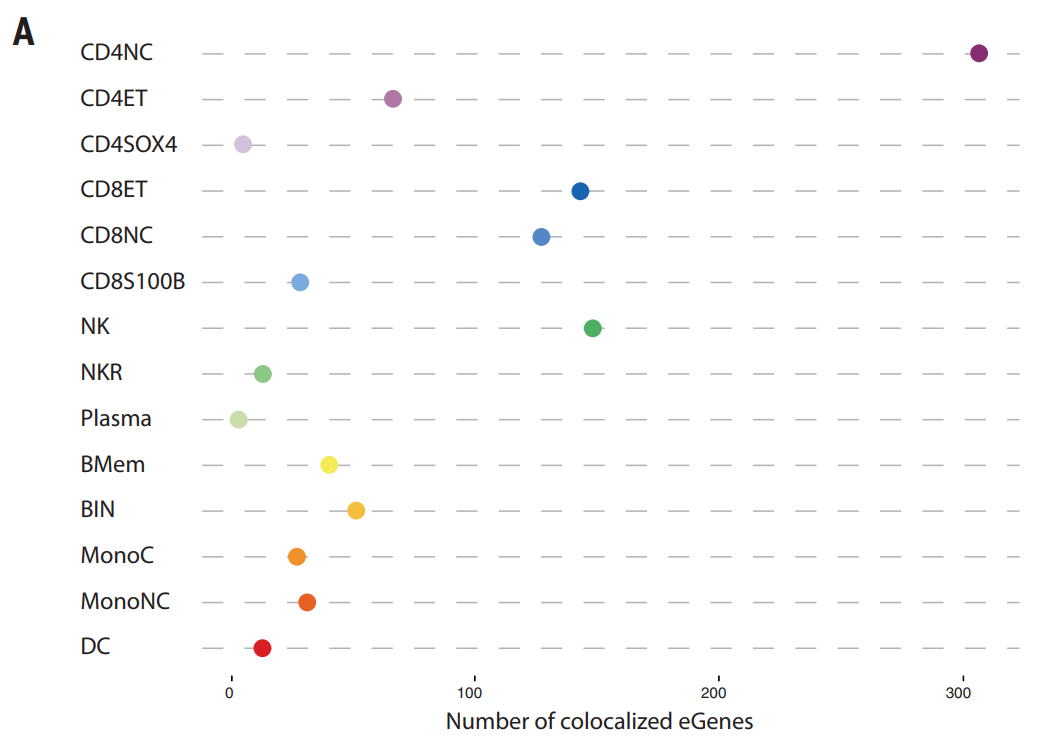
(A)研究使用贝叶斯因子分解与 MS GWAS 关联共定位的 cis-eGenes。共定位确定了 530 对通过 eQTL 具有共同因果关系的基因-细胞类型对。MS 疾病风险基因座的 eQTL 重叠具有高度的细胞类型特异性:在 108 个基因中,69 个仅在一种细胞类型中显示 eQTL 重叠。另外有 20 个基因的 eQTL 在两种细胞类型中被鉴定,10 个基因的 eQTL 在三种细胞类型中被鉴定,5 个基因的 eQTL 在四种细胞类型中被鉴定。相比之下,对于 METTL21B,在 CD4NC、CD4ET 和 CD8NC 细胞中观察到重叠的 eQTL。这些结果与研究对细胞类型特异性 eQTL 的观察结果一致,并为常见自身免疫性疾病的遗传风险提供了进一步的证据,这些疾病以高度细胞类型特异性的方式起作用,其中每个基因座通过改变有限数量的细胞类型的功能来发挥作用。不过,总的来说,遗传风险是由免疫系统赋予的。
虽然重叠的 GWAS SNP 和 eQTL 暗示改变的基因表达与疾病发病机制有关,但还有两种替代假设。一种解释是,GWAS 基因座和 eQTL 具有相同的因果变异,但对两种表型的影响是独立的,即多效性。第二种解释是,有两个独立的因果基因座,一个用于 GWAS 关联,另一个用于 eQTL。但它们彼此之间仍处于连锁不平衡状态。
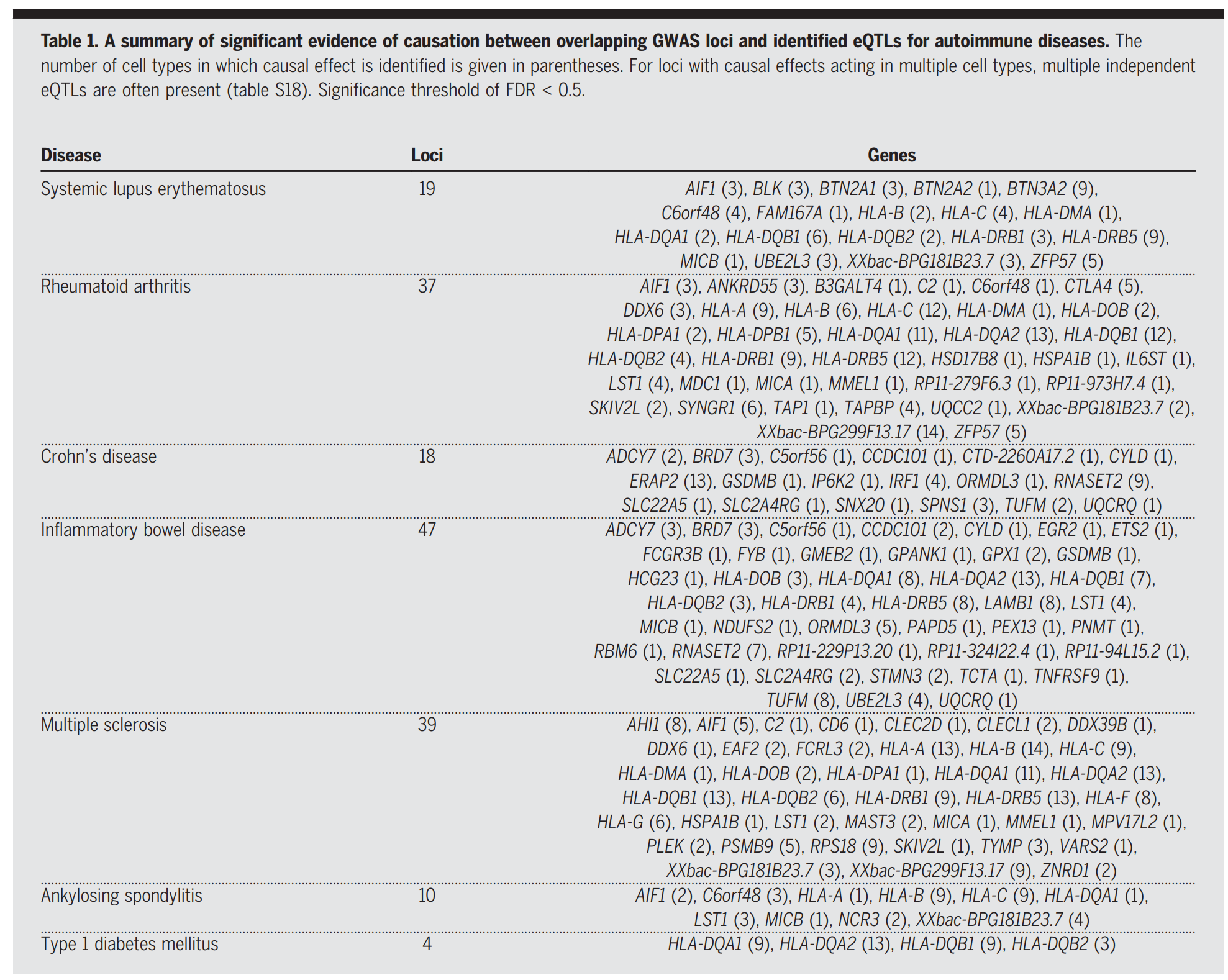
为了区分这两种假设,研究采用了孟德尔随机化方法,以确定自身免疫性疾病风险基因座起作用的因果方向的证据。使用七种自身免疫性疾病的 GWAS 数据,检验了 14 种细胞类型中所有疾病相关变异(p < 1 × 10−8)与 OneK1K eQTL 之间的因果关系。总共确定了 305 个基因座(FDR < 0.05),其中相关风险基因座被确定为通过改变一种或多种细胞类型中特定基因的表达而对疾病风险产生因果影响。在 305 个基因座中,188 个位于 MHC 区域,该区域的因果效应在细胞类型中普遍存在。其余 117 个基因座显示出高度细胞类型特异性的因果效应模式,其中 76 个基因座被确定为仅在一种细胞类型中具有因果效应。
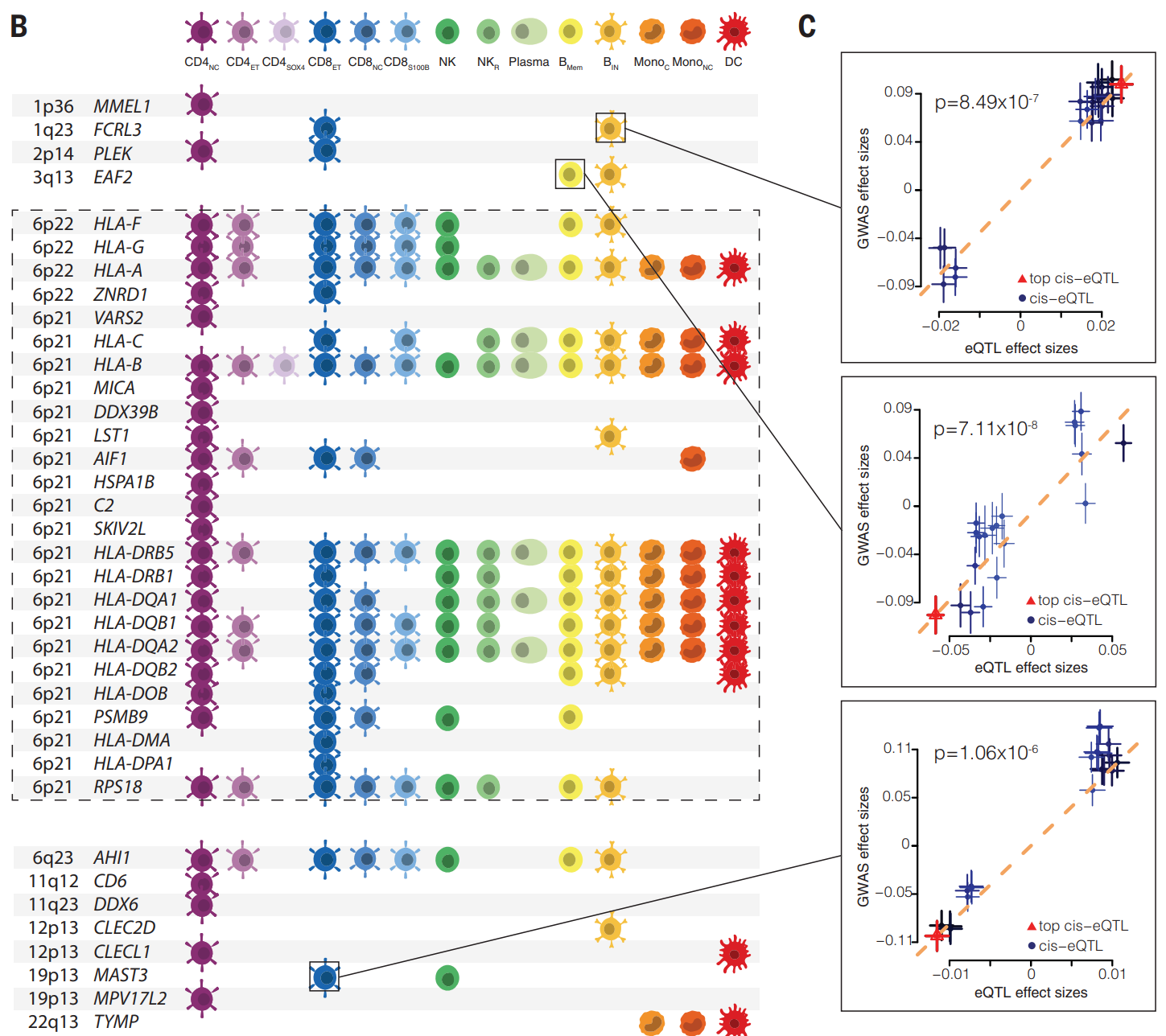
以 MS 为例,研究评估了 90 个风险基因座的因果基因及其作用的细胞类型。其中,研究能够根据 OneK1K 数据中一种或多种细胞类型中 eQTL 的重叠情况测试 57 个风险基因座的因果方向。
(B)孟德尔随机化用于确定 MS 重叠的 GWAS 位点与已鉴定的 eSNP 之间的因果关系。鉴定出 39 个 MS 相关基因 (FDR < 0.05) 的显著结果,其中 12 个位于 MHC 位点(虚线框)外的基因显示出高度的细胞类型特异性效应。在 MHC 区域确定了 73 个基因座,它们对 MS 风险的因果影响主要通过改变多种细胞类型中基因表达的变化起作用。例如,rs9264579 被确定为通过改变所有 14 种细胞类型中人类淋巴细胞抗原 B 类 (HLA-B) 表达起作用,而 rs9501393 仅通过改变 CD4NC 细胞中 SKIV2L 的表达水平产生因果关系。
(C)图中分别显示了 OneK1K eQTL SNP 与 MS GWAS 对 BIN、BMem 和 CD8ET 细胞中 FCRL3、EAF2 和 MAST3 的等位基因效应的效应大小。在 MHC 区域之外还确定了另外 17 个具有因果关系的基因座,它们以更特定于细胞类型的方式起作用。例如,1q23 区域中的 SNP 先前已被确定与 MS 相关,其中 rs7528684 标记的 FCRL3(p = 8.9 × 10−9)位于启动子元件内。分析确定位于近端的 FCRL3 是 CD8ET(p = 5.0 × 10−7)和 BIN(p = 6.6 × 10−7)细胞中 MS 风险的致病基因。
总结
研究内容
本研究揭示了循环免疫细胞中细胞类型特异性eQTL的等位基因结构。研究重点是绘制14种免疫细胞类型对基因表达的遗传效应,识别出超过26,000个独立的顺式eQTL和990个MHC基因座外的反式eQTL。
- eQTL效应:大多数eQTL具有等位基因效应,这些效应在很大程度上是细胞类型特异性的,并在两个独立的队列中得到了验证。确定了某些基因座在关键免疫功能途径中的作用。
- B细胞谱系分析:动态分析揭示了B细胞成熟过程中标志物的预期变化,并促进了动态eQTL的识别,这些eQTL在主要顺式eQTL分析中未被发现。
- 疾病关联:通过将scRNA-seq eQTL数据与GWAS确定的自身免疫风险基因座相结合,识别了这些基因座上的致病基因,并确定了这些基因发挥致病作用的细胞类型。进行了孟德尔随机化和共定位分析,探讨eQTL与疾病风险基因座之间的关系。共定位分析显示eQTL和GWAS风险基因座可能共享相同的致病基因座。
研究方法与优势
相较于从大量RNA测序数据中进行细胞反卷积的替代方法,单细胞eQTL分析具有显著优势,如能够识别以前未表征和罕见的细胞类型,以及准确量化转录丰度。进一步研究单细胞分辨率下的反式变异和基因环境相互作用,特别是在免疫系统中,这对于理解抗原或细胞因子如何影响细胞转录谱变化至关重要。
研究意义
本研究结合遗传流行病学与scRNA-seq数据,揭示了免疫系统个体间差异的驱动因素。研究结果强调了分离遗传变异如何以细胞类型特异性的方式影响编码关键免疫调节和信号通路的蛋白质的基因表达。此外,了解免疫系统调节的遗传基础将对自身免疫性疾病、感染、移植和癌症的治疗产生广泛影响。




















 835
835

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言











