










Uncoupling FRUITFULL’s functions through modification of a protein motif identified by co-ortholog analysis
通过共直系同源体分析识别出的蛋白质基序修饰来解耦 FRUITFULL 的功能
摘要
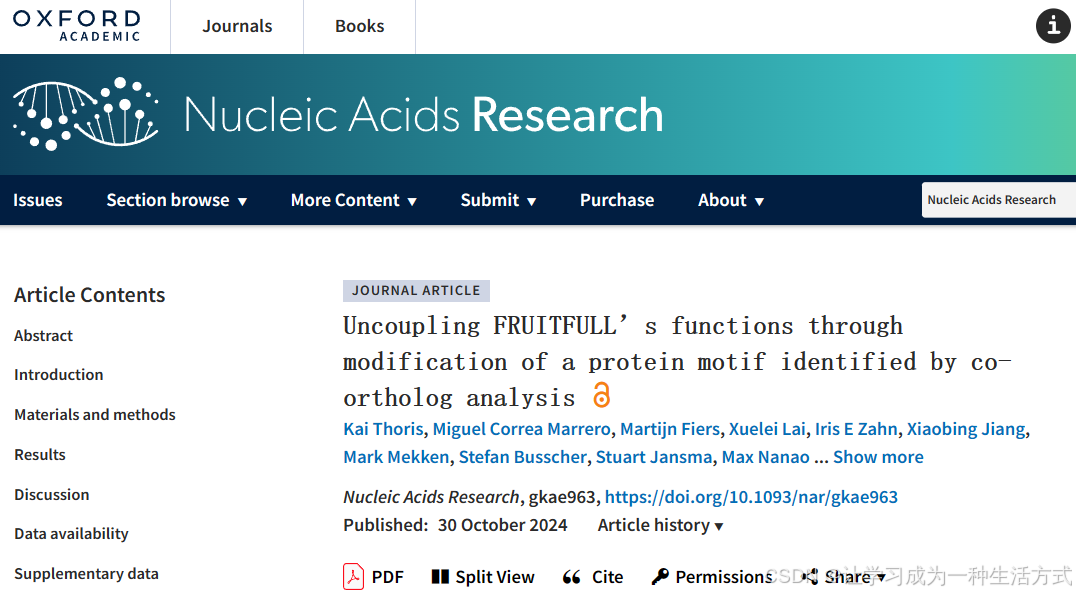
许多植物转录因子(TFs)具有多功能性,能够在多个组织中调控生长和发育。这些转录因子通常可以根据组织类型与不同的蛋白质伙伴结合,从而调控组织特异性的目标基因集。然而,如何确保这种特异性相互作用的实现仍然很不清楚。在本研究中,我们通过亚功能化的 FRUITFULL(FUL)亚家族 MADS 结构域转录因子的共直系同源体,研究了蛋白质-蛋白质相互作用的特异性。在拟南芥中,FUL 是多功能的,在开花和结果中发挥重要作用,而在番茄的共直系同源体 FUL1 和 FUL2 中,这些功能已部分分化。通过连接蛋白质序列和功能,我们发现了一个关键的氨基酸基序,它决定了 MADS 结构域转录因子的相互作用特异性。在拟南芥的 FUL 中,这个基序决定了与 AGAMOUS 和 SEPALLATA 蛋白的相互作用,并与一部分目标的调控相关。这一发现为解析多功能 MADS 转录因子的生物学功能提供了极好的机会。
引言
许多编码转录因子(TF)的基因在不同组织中表达并影响多种性状。这些多功能转录因子通常可以根据组织中可用的相互作用蛋白,与不同的蛋白质伙伴结合。在植物中,广泛存在的全基因组复制(WGD)导致了复杂的蛋白质相互作用网络,其中密切相关的(旁系同源的)多功能转录因子经历了不同程度的亚功能化,产生了独特且冗余或部分冗余的功能。在大型植物转录因子家族中,包括bHLH家族、MADS-box和AP2/ERF家族,这种现象尤其常见。例如,bHLH基因INDEHISCENT (IND)、SPATULA (SPT) 和 HECATE 1-3 (HEC1-3) 在雌蕊模式化、茎端分生组织特化和光形态建成方面具有独特和重叠的功能;MADS-box基因APETALA 1 (AP1)、CAULIFLOWER (CAL) 和 FRUITFULL (FUL) 在茎端分生组织、茎和果实发育中部分具有冗余功能;AP2/ERF家族基因APETALA 2 (AP2)、SCHLAFMÜTZE (SMZ)、SCHNARCHZAPFEN (SNZ) 和TARGET OF EAT 1-3 (TOE1-3) 在开花时间调控、花器官发生和果实发育中具有冗余和独立的功能。
基因的多功能性反映了现有网络在不同组织中的高效利用,但这些过程之间的相互关联阻碍了性状特异性适应,从而导致进化上的限制。这种相互关联在植物育种中也是一个挑战,因为多功能基因的突变会导致不良的多效性表型。理想情况下,育种者可以获得分离多功能基因的特定功能而不干扰其他功能的工具。近年来,通过检查顺式调控区域以识别组织特异性元件是一种有效的基因功能解析方法,已经开始取得成果。然而,在蛋白质水平上的功能解析尚未被广泛报道,尽管这可能是针对特定功能的强大策略,因为转录因子通常在不同组织中与不同的蛋白质相互作用。将蛋白质基序与组织特异性功能关联并不简单,因为这需要对特定蛋白质-蛋白质相互作用的氨基酸有详细了解。我们推测,研究亚功能化的旁系同源基因的蛋白质分化可以帮助识别这些氨基酸,从而能够在其他物种中解耦多功能直系同源基因的多效性功能。在这里,我们探索了这种方法,并描述了一种基于共直系同源体亚功能化来解析多功能转录因子的成功方法。
作为概念验证,我们聚焦于多功能转录因子FUL。FUL属于MADS-domain转录因子的MIKC型亚家族,该亚家族的成员参与了多种植物发育途径。这个亚家族的蛋白质特征在于四个保守结构域的存在:MADS (M) 结构域、中间 (I) 结构域、角蛋白样 (K) 结构域和羧基末端 (C) 结构域。为了发挥作用,MADS-box转录因子通过I和K结构域形成二聚体或四聚体,从而通过M结构域在所谓的CArG-box (CC[A/T]6GG) 上结合DNA。尽管CArG-box结合位点是共享的,似乎不同的MADS-domain复合物部分地调控了不同的目标基因集。最近的研究发现了一些影响特异性的因素,包括对CArG-box序列偏好和DNA形状识别的细微差异。拟南芥的FUL是多功能基因的典型例子,调控角果和基叶的发育、开花时间、植物结构和生命周期。它属于被子植物特有的AP1/FUL基因谱系中的euFUL分支。值得注意的是,在核心真双子叶植物的早期发生了两次复制事件,形成了三个分支:AP1、euFULI 和 euFULII。拟南芥的FUL属于euFULI分支,而表达很低的AGL79位于euFULII分支。
在其他一些真双子叶植物家族中,基因复制事件导致了多个 FUL 拷贝的出现,通过亚功能化减少了祖先的多效性。在茄科家族中,euFULI 分支的早期复制事件导致所有物种中出现了两个共直系同源体,即形成了 FUL1 和 FUL2 两个谱系。euFULII 分支中也发生了一次复制事件,产生了包含 MBP10 和 MBP20 样基因的两个亚谱系。番茄中的这些基因与拟南芥中多功能的 FUL 功能相似,参与调控开花时间、花序结构、果实发育和成熟,尤其是 FUL1 通过亚功能化表现出减少的蛋白质-蛋白质相互作用,并丧失了早期的果实功能。
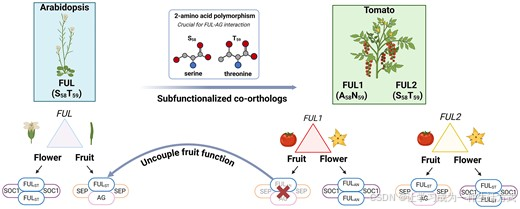
在本研究中,我们将番茄共直系同源体 FUL1 和 FUL2 中存在的多态性与它们的蛋白质功能差异联系起来,随后利用这些结果来分离它们的多功能拟南芥同源基因 FUL 的不同功能。我们展示了 FUL1 和 FUL2 之间的两个氨基酸多态性决定了与 AGAMOUS (AG) 和 SEPALLATA (SEP) 的果实特异性物理相互作用,而与茎端分生组织特异性的 SUPPRESSOR OF OVEREXPRESSION OF CONSTANS (SOC1) 的相互作用独立于该多态性。DAP-seq 实验揭示了 FUL-AG-SEP、FUL-SOC1 和 FUL-FUL 目标基因集之间的差异,这些差异与不同的结合位点偏好相关。在植物体内,对 FUL 中的两个氨基酸进行修饰会专门导致果实功能的降低。我们还展示了这两个氨基酸多态性是决定 MADS 结构域相互作用特异性的 12 个氨基酸基序的一部分。鉴于近年来大量植物基因组的测序结果以及广泛存在的谱系特异性全基因组复制(WGD),我们预计我们的方法可以普遍用于在蛋白质水平上解耦多功能转录因子的功能。
材料与方法
电泳迁移率变动分析
根据 Smaczniak 等人的方法(17)对电泳迁移率变动分析(EMSA)进行操作,进行了少量修改。所有编码序列(包括 FUL、SOC1、AG、SEP3、FUL-L-GFP、FUL-L-3xFLAG、FULAN、FUL1、FUL2、TAG1、FULSD、FUL1ST(C9)、FUL1AD、FUL1AT 和 FUL2AN(C8))均克隆至 pSPUTK(Stratagene)蛋白质表达载体中。蛋白质通过 TnT®SP6 高产小麦胚芽蛋白表达系统(Promega, L3260)按照制造商说明体外产生。所使用的 DNA 探针均约为 100 bp,从基因组 DNA 扩增并随后克隆至 pJET 载体(Thermo Scientific, K1231)中。接着,利用标记有 DY-682 的引物靶向载体骨架进行 PCR 扩增,并用柱子纯化(MACHEREY-NAGEL 740609.50S)。对于超移 EMSA,在孵育开始 30 分钟后向 EMSA 结合反应中加入 1 μl 的抗 FLAG 抗体(Miltenyi Biotec, 130-101-572)。所有 EMSA 凝胶使用 LiCor Odyssey 成像系统在 700 nm 处进行可视化。
DAP-seq
该方法按照先前描述的方法(24,35)进行,稍作修改。实验针对感兴趣的二聚体/四聚体(FUL-SOC1、FUL-AG-SEP3 和 FUL-FUL)以及 FUL-AG 和 FUL-SEP3 作为对照样本进行。此外,还包括一个输入样本以校正背景信号。蛋白质通过 TnT®SP6 高产小麦胚芽蛋白表达系统(Promega L3260)体外生成,按照制造商的说明操作。所有蛋白质均克隆至 pSPUTK(Stratagene)蛋白表达载体中,仅在 FUL 上附加了 3×FLAG 标签。对于单一蛋白质混合物(例如 FUL),使用 2 μg 质粒作为输入,而对于异质蛋白质混合物(FUL-SOC1、FUL-AG-SEP3、FUL-AG 和 FUL-SEP3),使用等摩尔比例的质粒,总量为 2 μg。所有后续步骤均在室温下进行,并使用 DNA LoBind® 管(Eppendorf 0030108051)以最大化样本回收率。体外生成的蛋白质(48 μl)和 DNA 库(400 ng)在 360 μl 的 EMSA 结合混合物(17)中孵育 2 小时,以确保最佳的蛋白质/DNA 结合条件。然后将 EMSA 结合反应加入 20 μl 已清洗的抗 FLAG® 磁珠(Sigma-Aldrich M8823)中,通过加入含有蛋白酶抑制剂(Roche 11697498001)的裂解缓冲液(Miltenyi Biotec 130-091-125)将体积增加至 1 mL。该混合物在管旋转器(Thermo Scientific 88881001)上孵育 2 小时,然后用 400 μl 1× TBS 进行三次洗涤。接下来,将结合的蛋白质通过与含有 150 ng/μl FLAG 肽(APExBIO A6001)的 400 μl 1× TBS 缓冲液孵育 45 分钟从磁珠上洗脱出来,之后在磁性支架上收集上清液。随后,以相似方式进行第二次洗脱,以获得总量为 800 μl 的洗脱液。收集的上清液在 95°C 下孵育 10 分钟后立即进行柱纯化(MACHEREY-NAGEL 740609.50S),并用 50 μl 洗脱缓冲液洗脱。洗脱的 DNA 片段使用 Q5® 高保真 DNA 聚合酶(NEB M0491)和带有独特条形码的 Illumina TruSeq 接头进行 20 个循环的扩增,随后使用 AMPure XP 磁珠(Beckman A63880)纯化。纯化的样本加载至 1% 琼脂糖凝胶,切下 250 至 600 bp 的片段,从凝胶中纯化(MACHEREY-NAGEL 740609.50S),并使用 AMPure XP 磁珠再次进行纯化。带有不同条形码的样本以等摩尔比混合后,用 NovaSeq(Novogene)进行双端测序(150 次循环)。每个样本约获得 10–30 百万读数,每种实验条件均进行三次重复。
生物信息学分析
使用 Trim Galore (https://github.com/FelixKrueger/TrimGalore) 对读数进行修剪,并用 HISAT2 (36) 将其比对到拟南芥基因组 (TAIR10)。然后,BAM 文件用于 MACS2 峰值调用(37),显著性阈值为 P < 0.05 或 P < 0.0001,并使用 MACS2 bdgdiff(37)进行差异峰值分析。然后使用 ChiPseeker (38) 对峰值进行注释。为了获得基序文件,我们使用 MEME (39) 和 GEM (21) 进行了额外的峰值分析。所有峰值和测序数据均通过 IGV 基因组浏览器 (40) 进行可视化。
酵母双杂交
酵母双杂交实验严格按照描述的方法(41)进行。含有 MADS 结构域编码序列 (CDS) 的酵母克隆体分别在 pDEST22 (AD-载体, Invitrogen) 或 pDEST32 (BD-载体, Invitrogen) 中获得,部分克隆体来自 De Folter 和 Immink 2011 的收藏,或使用 Gateway 系统克隆并转化至酵母菌株 PJ69-4A (pDEST22) 和 PJ69-4α (pDEST32)。为在蛋白质序列间交换氨基酸,使用了叠加延伸 PCR 组合 PCR 片段。PCR 扩增和克隆的引物列在补充表 S4 中,突变蛋白的序列列在补充表 S5 中。相互作用筛选在 LWH 缺陷培养基上进行,并补充不同浓度的 3-氨基-1,2,4-三唑 (3-AT)。培养板在室温下孵育,3-6 天后成像。
拟南芥过表达和互补株系的生成
拟南芥植物在生长箱中生长,相对湿度为 70%,温度为 20°C,在 LED 光照下(150 μmol m⁻² s⁻¹)以 16 小时光照/8 小时黑暗的昼夜周期生长,种植在岩棉塞(Grodan)上,每周用 Hyponex 肥料(1 g/L)浇水两次。本研究中使用的所有植物均为 Col-0 背景,包括互补株系、过表达株系和 ful-7 突变体 (SALK_033647)。所有互补株系的构建均使用 Gateway 重组克隆(Invitrogen)生成,有些在 Golden Gate 或 GreenGate 载体(42,43)中预组装。引物列在补充表 S4 中。原生拟南芥 FUL 启动子 (5.2kb) 和终止子 (1.2kb) 与 FUL (AT5G60910)、FUL1 (Solyc06g069430)、FUL2 (Solyc03g114830)、MBP10 (Solyc02g065730) 或 MBP20 (Solyc02g089210) 的编码序列融合。带有氨基酸交换的互补株系 (FULAN, FUL1ST, FUL2AN) 通过重叠延伸 PCR 和/或 Golden Gate 克隆引入突变后获得。随后,将质粒转化至农杆菌株 C58C1,并在 ful-7 突变体 (SALK_033647) 背景中转化。
qRT-PCR 分析
从每个独立株系(单株)采集主花序上的 5 个第 12–16 阶段的角果(位置 5–10)(44)。使用 CTAB/LiCl 提取 RNA,并用 DNase(TURBO™ DNase,Invitrogen, AM1907)处理。cDNA 合成使用 SuperScript™ II 逆转录酶(Invitrogen, 18064014)进行。使用 iQ™ SYBR® Green(Bio-Rad, 1708880)进行 FUL、FUL1 和 FUL2 以及参考基因 UBC21 的 RT-qPCR,采用标准的 40 个循环的两步程序,退火温度为 60°C。测定所有引物对的引物效率,仅使用效率相同的引物。所有使用的引物列在补充表 S4 中。
I 结构域基序识别
使用 hmmsearch v3.2.1(45)从不同数据库中收集了来自 1006 个植物物种的 MIKC 蛋白序列,以使用相关的 Pfam 模型(分别为 PF00319.18 和 PF01486.17)(46)扫描包含 MADS 和 K 结构域的序列。去除含有不完整结构域的蛋白质后,使用 Clustal Omega v1.2.4(47)生成多序列比对。去除长度异常的序列(<1.5× 高于或低于中值的标准差),最终得到 11966 个 MIKC 型蛋白质序列的比对。在检测到的 M 和 K 结构域之间的子序列被视为 I 区域。我们使用 MEME v5.0.5(48)在收集的 I 区域中挖掘基序。我们允许最多 75 种不同的基序,设置基序的最小宽度为 4 个残基,最大宽度为 75 个残基。我们在每个序列至多一个基序出现的模型选项下使用 MEME(即 MEME 假设每个序列最多含有一次出现的每个基序)。随后,使用 FIMO 5.0.5(39,49)在 I 和 C 端区域中搜索发现的基序的单独出现。
结构预测
使用 AlphaFold-multimer 模式下的 Colabfold 预测 MADS 复合体(同源和异源二聚体)的结构(50,51)。为每个 MADS 结构域转录因子复合体生成五个模型并进行视觉检查。对于所有复合体,基于 IPTM 分数选择得分最高的模型,并生成符合拟南芥 SEP3 DNA 结合结构域同源二聚体 X 射线晶体结构(PDB 7NB0;rcsb.org (52))的正确 DNA 结合结构域。然后将 AF2 模型提交至 EBI 的蛋白质界面表面和组装(PISA)服务器,以计算界面的埋藏表面积和形成界面的溶剂化自由能增益 ΔG(53,54)。
结果
拟南芥 FUL 通过与不同的蛋白质伙伴结合来执行其多重功能
拟南芥 FUL 的多功能性体现在其蛋白质-蛋白质相互作用模式中,其中它可以与 MADS 结构域转录因子 SEPALLATA 1 (SEP1)、SEP2、SEP3、SEP4、AGAMOUS (AG) 和 SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1) 等蛋白相互作用。FUL 在执行重要功能的组织中形成不同的复合体,在花序分生组织(IM)中与 SOC1 结合,在果实中与 AG 和 SEP1-3 结合。为了确定 FUL 在 IM 和果实中的优先复合物组成,我们使用包含典型 CArG-box 的探针和不同组合的 FUL/SOC1(图 1A)及 FUL/AG/SEP3(图 1B)进行了 EMSA 实验。为了区分大小相似的二聚体和四聚体,我们还加入了 FUL-GFP (F-G) 和 FUL-FLAG (F-F) 蛋白,并添加抗 FLAG 抗体以对包含 FUL 的复合体进行超移。这表明,当 FUL 和 SOC1 混合时,FUL-SOC1 异源四聚体最为显著,而组合 FUL、SEP3 和 AG 则产生了不同的复合体,其中 FUL-AG 二聚体和 FUL-SEP3-AG-SEP3 四聚体是含 FUL 的最主要复合体(图 1A–C;补充图 S1)。因此,在 IM 中,FUL-SOC1 四聚体与目标基因结合,而在果实中,FUL-AG 二聚体和 FUL-SEP-AG-SEP 四聚体可能最显著地结合以调控其表达(图 1C)。此后,我们将分别称这些为 FUL–SOC1(IM)和 FUL–AG–SEP(果实)复合体。
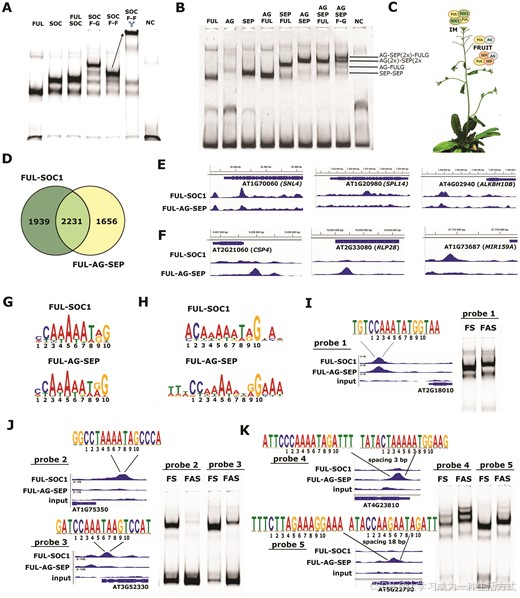
图1. FUL 通过形成不同复合物来执行其多重功能,从而调控不同的目标基因集。(A) 使用包含单个典型 CArG-box 的 SAUR10 探针及不同组合的 FUL 和 SOC1 蛋白进行的 EMSA 实验。在 SOC F-F 的右侧通道上方的 "Y" 表示加入了 FLAG 抗体,箭头表示包含抗体的复合物的迁移(SOC1(2x) 和 FUL-FLAG(2x) 的四聚体)。虽然 FUL 可以形成同源二聚体,SOC1 可以形成同源二聚体和四聚体,但右侧通道的明显迁移表明,当 FUL 和 SOC1 结合时,FUL-SOC1 四聚体最为显著。(B) 类似于 (A) 的 EMSA 实验,但使用 FUL、SEP3 和 AG 蛋白。FUL 同源二聚体的迁移速度略快于 AG 和 SEP 同源二聚体(通道 1–3)。FUL 与 AG 形成强二聚体(通道 4),并与 SEP3 主要形成四聚体(通道 5)。AG-SEP 也可以形成四聚体,其迁移速度比 FUL-SEP 稍慢(通道 6)。当 FUL、SEP 和 AG 结合时(通道 7),FUL-GFP 的测试(通道 8)表明主要形成以下复合物:SEP 同源二聚体、FUL-AG 二聚体、AG-SEP-AG-SEP 四聚体和 FUL-SEP-AG-SEP 四聚体。(C) 示意图表示 FUL-SOC1 四聚体在 IM 中是主导复合物,而由 FUL、AG 和 SEP 组成的复合物在角果(果实)中占主导地位。(D) 维恩图显示了 FUL-SOC1(三个重复结合)和 FUL-AG-SEP(三个重复结合)的 DAP-seq 的独特和重叠目标基因集(P < 0.05)。(E, F) 基因组浏览器快照展示了三种显著富集于 FS(E)和三种显著富集于 FAS(P < 0.0001)的基因(F)。Y 轴表示基于三个重复与三个输入重复的平均值的归一化读数。在大多数情况下,Y 轴的量程为 [0, 4],仅在 RLP28 中,峰值更高,量程为 [0–10]。(G, H) 使用 MEME 确定的 PWM 的序列标志图,使用所有富集位点计算 FUL-SOC1(顶部)或 FUL-AG-SEP(底部)的 PWM(G),或仅使用最前 100 个差异富集的峰值(H)。(I-K) 使用 FUL-SOC1(FS)和 FUL-AG-SEP(FAS)进行 EMSA 实验,以测试探针与不同类型的 CArG-box(在峰值上方指示)的结合能力。(I) 具有典型 CArG-box 的 SAUR10 探针;(J) FUL-SOC1 在 DAP-seq 中优先结合的探针片段;(K) FUL-AG-SEP 在 DAP-seq 中优先结合的探针片段。
为了探讨 FUL-SOC1 和 FUL-AG-SEP 是否结合到不同的靶基因集合上,我们使用 FUL-FLAG 蛋白分别与 SOC1 以及 AG 和 SEP3 组合,进行了 DAP-seq(24,35)实验。在 FUL-SOC1 的 DAP-seq 中,共识别出 4170 个显著结合事件,而在 FUL-AG-SEP 中识别出 3887 个(P < 0.05,参见补充表 S1),在 P < 0.0001 的显著性水平下分别有 628 和 964 个事件(参见补充表 S2)。为了快速评估识别到的靶位点的性质,我们进行了 GO 术语富集分析,结果显示“心皮形成”和“花分生组织决定性”是最富集的生物过程之一(参见补充图 S2),在最富集的分子功能中则包括“DNA 结合转录因子活性”(参见补充图 S3),这些结果进一步验证了分析的质量。正如转录因子复合体的预期,大多数结合事件定位于靶基因的启动子区域(参见补充图 S4)。由于两个复合体都预计会结合 CArG-box,因此我们并不意外地发现 FUL-SOC1 和 FUL-AG-SEP 数据集之间有 2231 个结合事件重叠。然而,在 P < 0.05 的截断值下,FUL-SOC1 或 FUL-AG-SEP 的列表中也出现了相当数量仅出现在其中一个复合体的结合事件(图 1D)。随着显著性阈值的变化,“独特”和“重叠”列表的组成也会发生变化,因为“独特”峰值中的许多反映的更多是峰值高度的不同,而非明确的“结合”与“未结合”状态。
在 P < 0.0001 水平下查看靶基因列表(参见补充表 S2),我们识别出一些在两个复合体中仅出现显著结合事件的有趣靶点。例如,在 FUL-SOC1 数据中,对植物结构和开花时间相关的基因 SPL14、在短日照条件下通过 HDAC19 组蛋白去乙酰化复合物抑制开花的 SNL4、以及参与花期转换的 RNA N6-甲基腺苷去甲基化酶编码基因 ALKBH10B 检测到显著峰值(图 1E)。这些结果表明 FUL-SOC1 可能确实以更高的亲和力结合某些开花相关基因,而非 FUL-AG-SEP。然而,这种分析方法得出的一些推测性差异结合事件高度依赖于选择的显著性阈值,有时仅反映了结合亲和力的细微差异,例如 ALKBH10B 的峰值在两个复合体中在 P < 0.05 时均显著,但仅在 FUL-SOC1 中在 P < 0.0001 时显著(图 1E)。因此,我们进行了差异读数计数分析,并对变化倍数最高的靶基因进行了排序(参见补充表 S3)。对于 FUL-AG-SEP 的显著差异结合位点,例如受体样蛋白 RLP28 和与长角果生长相关的冷休克域蛋白 CSP4(图 1F);对于 FUL-SOC1,MIR159A 上游的一个区域被高度富集,该区域靶向几个 MYB 结构域转录因子(图 1F)。因此,尽管 FUL-SOC1 和 FUL-AG-SEP 的体外结合靶点集合大部分重叠,但也存在明显的差异,并且这些差异在一定程度上与两个复合体的推定生物学功能相关。
有趣的是,基于所有靶点计算的 FUL-SOC1 和 FUL-AG-SEP 位置权重矩阵(PWM)序列标识之间没有明显差异(图 1G),但在使用结合差异性最大的前 100 个位点时,识别出两个明显不同的 PWM(图 1H)。在 FUL-SOC1 的 PWM 中,规范基序第一个位置的 C 的限制较宽松(CC[A/T]6GG),这一点很引人注目,因为该 C 在不同 MADS 结构域复合体的结合基序中高度保守(25)。另一方面,基于 FUL-AG-SEP 特异靶点的 PWM 比标准的 CArG-box 基序显示出更多限制,并且在 PWM 的 5′ 和 3′ 端都出现了 A/T 延伸(图 1H)。为了验证 FUL-SOC1 和 FUL-AG-SEP 确实结合在不同位点上,我们使用从 DAP-seq 中选择的五个不同探针片段进行了 EMSA 实验。FUL-SOC1 和 FUL-AG-SEP 都以类似的亲和力结合到规范的 SAUR10 CArG-box 探针上(图 1I),但只有 FUL-SOC1 能够以高亲和力结合到探针片段 2 和 3 中的“宽松” CArG-box 位点上(图 1J)。探针片段 4 和 5 是从在 DAP-seq 中由 FUL-AG-SEP 特异结合的位点扩增的,每个包含两个 CArG-box,分别间隔 3 和 18 bp(图 1K)。探针 4 中的一个 CArG-box 与 FUL-AG-SEP 特异的 PWM 非常相似,并且该探针确实被 FUL-AG-SEP 更强烈地结合。探针 5 中的 CArG-box 各包含 C/G 在框的 A/T 核心中(见图 1K),通常认为这种变化会完全阻止结合(25,27)。然而,EMSA 显示 FUL-SOC1 和 FUL-AG-SEP 都能以类似的亲和力结合此探针,这可能得益于 CArG-box 两端完美的 A/T 延伸。因此,尽管之前已显示出 flanking A/T 核苷酸的贡献(25,26),但这些核苷酸似乎对 FUL-AG-SEP 的结合特别重要,而 FUL-SOC1 通常能够高亲和力结合各种 CArG-box,对 flanking 核苷酸的依赖性较小。然而,其对扰动 CArG-box(即 A/T 核心中的 C/G)的结合也可能通过双侧的 A/T 延伸得以实现,这在探针 5 中得到了验证。DAP-seq 和 EMSA 在探针 5 上的结果差异可能是由于基于基因组的 DAP-seq 片段和基于 PCR 的 EMSA 片段之间的 DNA 形状差异造成的,因为 DNA 形状读取也已显示对结合亲和力有所贡献(22,23,26)。
综上所述,拟南芥的 FUL 与不同的 MADS 结构域蛋白结合,这些 MADS 结构域复合体在结合不同的 CArG-box 基序上存在差异,这可能有助于差异化的靶基因调控,使特定的 FUL 复合体能够在调控开花时间和果实发育中发挥其功能。
番茄 FUL 蛋白在相互作用模式和功能上有所不同
番茄中包含四个 FUL 类基因,这些基因是由两个茄科特异性复制事件产生的,一个位于 euFULI 类群,另一个位于 euFULII 类群(31)。四个番茄 FUL 基因表现出功能分化,这在它们的蛋白质-蛋白质相互作用特征中得以体现(32)。多功能的番茄 FUL2 具有广泛的相互作用模式,与其在开花、果实发育和果实成熟过程中的不同功能一致(32,33),而功能细化的 FUL1 和 MBP20 则表现出更特定的相互作用特征,非功能化的 MBP10 几乎没有相互作用(32)。有趣的是,FUL2 可以与番茄的 AG 同源物 TAG1(TOMATO AG 1)相互作用,而其旁系同源物 FUL1 则不能(32)。另一方面,FUL1 和 FUL2 都可以与番茄的 SOC1 共同同源物 TM3(TOMATO MADS 3)和 STM3(SISTER OF TM3)相互作用(32,62),这表明尽管 FUL1 与 FUL2 在蛋白质相似性上很高(参见补充图 S5),但 FUL1 已经实现了功能细化。
我们通过测试番茄 FUL 类蛋白与一组已知的拟南芥 FUL 相互作用蛋白的相互作用能力,确认了番茄 FUL1 相对于拟南芥 FUL 的功能细化(图 2A)。与番茄的相互作用研究结果一致,FUL1 无法与 AG 相互作用,且相互作用伙伴的数量比 FUL2 少,而 FUL2 可以与 AG、SEP3 和其他七种蛋白相互作用。同样,MBP20 的相互作用伙伴整体上少于 FUL2,且无法与 AG 相互作用。FUL1、FUL2 和 MBP20 都能够与 SOC1 强烈相互作用,表明它们氨基酸序列中的多态性并不影响一般的相互作用能力。有趣的是,在酵母双杂交试验中,番茄的 FUL 类蛋白均无法自二聚,而拟南芥 FUL 在 EMSA 和酵母双杂交试验中都可以自二聚(图 1A 和 2A)。
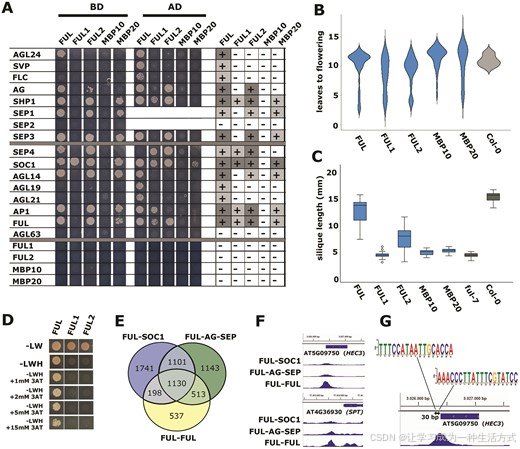
图 2. FUL 类蛋白具有不同的蛋白质-蛋白质相互作用模式,并在拟南芥果实中的功能存在差异。(A) 酵母双杂交实验显示了来自番茄(FUL1、FUL2、MBP10、MBP20)和拟南芥(FUL)的 FUL 类蛋白与一组拟南芥 MADS 结构域蛋白的物理相互作用。右侧的列总结了两种方向(AD 与 BD,BD 与 AD)的数据。在 -LWH 平板上以 5mM 3-AT 进行筛选。由于 AD 或 BD 结构域与 MADS 结构域蛋白的 N 末端融合可能会影响相互作用(例如(55)),我们在两种方向上进行筛选(见图 A 上方标记)。SEP1-BD 和可能的 SEP2-BD 显示自激活,因此仅在 AD 结构域融合时筛选。SEP3-BD 使用了缺少 C 末端的截短版本,以避免自激活(56)。(B) 小提琴图显示了独立的 35S:thFUL 转基因株系的开花时间分布,以开花前的叶片数量来测量。每个构建至少有 30 个转基因株系进行了表型分析。x 轴表示 FUL = 35S:FUL,FUL1 = 35S:FUL1,依此类推。(C) 箱线图显示了在 ful-7 突变体背景下独立的 pFUL:thFUL:tFUL 株系的角果长度分布。每个构建至少有 15 个转基因株系进行了表型分析。x 轴表示 FUL = pFUL:FUL:tFUL,FUL1 = pFUL:FUL1:tFUL,依此类推。(D) 酵母双杂交实验测试了 FUL、FUL1 和 FUL2 在不同浓度的竞争物 3-AT 下的自二聚能力。(E) Venn 图显示了 FUL-FUL、FUL-SOC1 和 FUL-AG-SEP 的 DAP-seq 数据集中的独特和重叠的显著靶点(P < 0.05)(每种数据集组合了三个重复)。(F) 基因组浏览器快照展示了 FUL-FUL、FUL-SOC1 或 FUL-AG-SEP 在 HEC3 和 SPT 位点的情况。y 轴显示基于三个重复与三个输入重复的平均归一化读数。在 SPT 的情况下,y 轴范围为 [0–4],在 HEC3 的情况下为 [0–15]。(G) HEC3 峰下存在的非经典 CArG-box。
鉴于拟南芥中的 FUL–SOC1 复合体调节向开花的过渡,而 FUL–AG–SEP 复合体参与果实发育,我们预测番茄 FUL2 蛋白能够在很大程度上补偿拟南芥 ful 突变体,而 FUL1 和 MBP20 只能拯救花期过渡表型。为了验证这一点并进一步测试功能细化的番茄 FUL 类基因是否可以用于分离拟南芥 FUL 的功能,我们生成了两种类型的转基因株系。由于拟南芥 ful 突变体的开花时间表型较轻(6),我们在 Col-0 背景下生成了 35S:thFUL 转基因株系,以清晰显示调节开花时间的能力(thFUL,指番茄 FUL 的同源基因,包括 FUL1、FUL2、MBP10 或 MBP20)。在 WT 背景中过表达 MBP20、FUL1、FUL2 和 FUL 生成了非常早开花的转基因株系,显示了较轻微效应的株系(图 2B),说明这四个基因在拟南芥中以相似方式影响开花时间的能力。
在第二组转基因株系中,将 ful-7 (27) 敲除突变体与番茄 FUL 类基因和拟南芥 FUL 进行互补,每个基因均使用拟南芥 FUL 的启动子和终止子(pFUL:(th)FUL:tFUL),以评估对突变体短角果表型的补救效果(图 2C)。有趣的是,拟南芥 FUL 几乎完全拯救了果实表型,而 FUL2 是唯一能够部分补救该表型的番茄共同同源基因。用 FUL1、MBP10 和 MBP20 进行互补后产生的角果与 ful-7 突变体的角果相似。因此,FUL1 和 MBP20 无法与 AG 相互作用可能影响了它们在拟南芥果实中的功能,而它们在与 SOC1 相关的开花功能上与拟南芥 FUL 类似。
我们质疑 FUL 和 FUL2 蛋白互补能力的差异是否可以通过拟南芥 FUL 能够形成同二聚体而番茄 FUL2 不能来解释。首先,我们确认即使在非常低浓度的竞争抑制剂 3-氨基-1,2,4-三氮唑(3-AT)下,FUL2 也不能自二聚(图 2D 和补充图 S6)。为了进一步研究除了 FUL-AG-SEP 外,FUL–FUL 复合体是否也参与了角果发育,我们还对 FUL-FUL 进行了 DAP-seq 实验。这得到了 2589 个显著富集的位点(P < 0.05),其中许多与其他复合体共享(图 2E)。有趣的是,FUL-FUL 显著结合了 537 个位点,而 FUL-SOC1 或 FUL-AG-SEP 则未显著结合。在将该列表与 ful 突变体角果中差异表达的基因进行重叠分析时,发现心皮发育基因 HECATE 3 (HEC3) 是 FUL-FUL 同二聚体的一个显著靶点(图 2F)。此外,SPATULA (SPT) 作为 HECs 在心皮发育过程中的相互作用伙伴,相比于 FUL-SOC1 或 FUL-AG-SEP 更显著地被 FUL 同二聚体结合(图 2F)。SPT 和 HEC1-3 一同作用以调节心皮融合和雌蕊发育(63),因此结果表明 FUL 的同二聚化对角果发育可能非常重要。我们调查了 HEC3 峰下的 CArG-box 序列,发现其含有两个间隔 30 bp 的非典型 CArG-box,分别为 CC[A/T]6CG 或 CC[A/T]6GC,而非典型的 CArG-box (CC[A/T]6GG)(图 2G)。因此,FUL-FUL 似乎能够结合 FUL-SOC1 或 FUL-AG-SEP 几乎不结合的变异 CArG-box。这一特性可能使同二聚体在拟南芥角果发育中不可或缺。总体而言,我们的数据表明,FUL 的角果功能既依赖于同二聚体的形成,也依赖于与 AG-SEP 的相互作用,而如果能识别出导致与 AG/SEP 相互作用和/或同二聚化丧失的氨基酸基序,同时保留与 SOC1 的相互作用,则有可能实现开花和果实功能的分离。
番茄 FUL1 与 TAG1/AG 的相互作用丧失是由一个 2 个氨基酸的多态性引起的
我们推测,由于 FUL1 和 FUL2 之间的序列相似性很高(参见补充图 S5),识别出导致 FUL1 和 FUL2 在相互作用能力(即与 AG/SEP3 相互作用)上不同的氨基酸可能相对容易。因为 MIKC 型蛋白的 I(间隔)域在蛋白-蛋白相互作用特异性中起重要作用(图 3A)(64,65),我们在 FUL1 和 FUL2 中交换了蛋白域,并评估了其对与 TM3、TM29 和 TAG1 的相互作用的影响,它们分别是番茄中 SOC1、SEP3 和 AG 的同源物(图 3B、C)。首先,我们确认了 I 区域是决定观察到的相互作用模式的关键(构建 C1-C4)。然后,我们根据 FUL1 和 FUL2 之间的序列差异(参见补充图 S5,图 3B;构建 C5-C9)交换了从一个到六个氨基酸的小基序,并发现 M-和 I 域交界处的一个 2 个氨基酸基序(图 3C)是与 TAG1 相互作用所必需的。该基序位于氨基酸位点 58 和 59,在 FUL1 中包含丙氨酸和天冬酰胺(AN),而在 FUL2 中为丝氨酸和苏氨酸(ST)。如果仅将 AN 换成 ST,FUL1 就能获得与 TAG1 相互作用的能力(图 3B、C)。
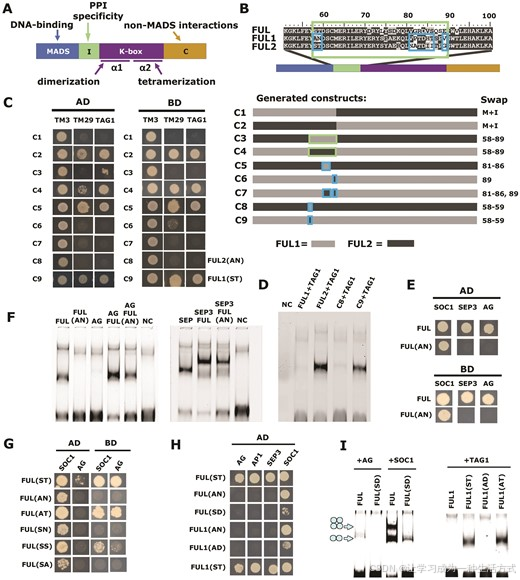
图 3. 番茄 FUL1 与 TAG1/AG 的相互作用丧失是由 I 域中的 2 个氨基酸多态性引起的。(A) MADS 结构域转录因子的结构示意图。(B) 不同构建体的示意图。大方框表示整个 I 域;内部的小方框表示 I 域中已被交换的不同基序。(C) 酵母双杂交结果显示含有不同构建体的诱饵蛋白与 TM3、TM29 或 TAG1 的猎物相互作用(左)或相反(右)。(D) EMSA 显示 AN/ST 多态性交换导致 CArG-box 探针的结合能力增益或丧失,可能是由于与 TAG1 相互作用的增益/丧失引起的。(E) 酵母双杂交结果显示当拟南芥 FUL 的 58–59 位氨基酸 ST 更换为 AN 时,FUL 失去了与 AG 的相互作用。(F) EMSA 结果显示 FUL(AN) 的同二聚体和异二聚体与含 CArG-box 的探针的结合强度减弱或完全消失(参见补充图 S6)。NC = 负对照(仅添加 TNT 反应混合物的探针)。(G, H) 酵母双杂交数据显示了修改后的 FUL 蛋白与 SOC1、AG、AP1 和 SEP3 的相互作用。(I) EMSA 用于测试不同版本的 FUL 和 FUL1 与 AG、SOC1 或 TAG1 的相互作用及其对含 CArG-box 的探针的结合能力。
为了确认这些结果,我们使用体外产生的 FUL1、FUL2、C8(FUL1(ST))、C9(FUL2(AN))和 TAG1 进行了 EMSA 实验(图 3D)。同样地,FUL1(ST) 获得了与 TAG1 相互作用的能力,而 FUL2(AN) 失去了该能力。接下来,我们研究了在拟南芥 FUL 中更换相应的氨基酸是否会产生相同的效果,从而消除 FUL 与 AG 和 SEP3 的相互作用,同时保留与 SOC1 的相互作用。FUL(AN) 保留了与 SOC1 的相互作用,但失去了与 SEP3 和 AG 的相互作用(图 3E)。虽然在一些 EMSA 实验中观察到 FUL(AN) 与 AG 和/或 SEP3 的弱相互作用(图 3F),但在其他实验中未观察到(补充图 S6 和 S7)。
为了确定 Ser58 和 Thr59 是否同等重要,我们使用 FUL(ST)(野生型)、FUL(AN)、FUL(AT)、FUL(SN)、FUL(SS) 或 FUL(SA) 进行了酵母双杂交实验(图 3G 和补充图 S8)。有趣的是,仅在 Thr59 存在的情况下,AG 的相互作用得以保留,无论位置 58 是 A 还是 S。然而,Ser58 对相互作用强度有所贡献,因为 FUL(AT) 与 FUL(ST) 相比,酵母生长较少。类似地,番茄 FUL1(AT) 能够与 AG、AP1 和 SEP3 进行相互作用,与 FUL1(ST) 类似(图 3H),但与 AG 的相互作用酵母生长较少(补充图 S8)。在更大范围的拟南芥 MADS 结构域蛋白中测试不同的 FUL 组合时,FUL(SA)/FUL(SN) 与 FUL(SS) 表现出显著差异,后者可以与许多拟南芥 MADS 结构域蛋白相互作用,类似于 FUL(AT/ST)(补充图 S9 和 S10)。因此,FUL(SS) 特异性地丧失了与 AG 的相互作用,但保留了与其他许多蛋白的相互作用。
发现位置 59 的 Thr 或 Ser 很重要后,我们假设磷酸化可能参与其中。为了测试这一点,我们设计了磷酸模拟版本的 FUL 和 FUL1,将天冬氨酸(Asp;D)引入位置 59 以代替 Thr 或 Asn,模拟磷酸化的 Thr(66)。然而,这导致了整体相互作用能力的显著下降,包括与 SOC1 的相互作用显著减少(图 3H 和补充图 S8)。EMSA 实验还显示,如果 Thr59 被 Asp59 替代,FUL 和 FUL1 无法与(番茄)AG 相互作用,且与 SOC1 的相互作用减弱(图 3I),因此未提供磷酸化参与的证据。这些结果与 Mengler 等人(67)的数据一致,他们未在 FUL 中检测到磷酸化位点。因此,Thr59 对 FUL 与其 MADS 结构域伙伴的相互作用至关重要,除了与 SOC1 的相互作用外,该相互作用允许其他残基位于位置 59。Thr59 可被 Ser59 替代,但这会导致特异性地失去与 AG 的相互作用。
如果与 AG/SEP 的相互作用受损,FUL 在角果中的生物活性会降低
为了测试与 AG/SEP 相互作用的丧失和获得对 FUL 在角果中生物学作用的影响,我们将 pFUL:FUL(AN):tFUL、pFUL:FUL1(ST):tFUL 和 pFUL:FUL2(AN):tFUL 转化到拟南芥 ful-7 突变体中,测量独立转基因株系的平均角果长度(每个构建体超过 15 个株系),并将结果与相应的 pFUL:(th)FUL:tFUL 株系进行比较。符合我们的预期,将 FUL 和 FUL2 中位置 58–59 的 ST 换为 AN 会降低其补偿 ful-7 角果表型的整体能力,而在 FUL1 中引入 ST 代替 AN 则增强了其补偿能力(图 4A)。然而,不同独立转基因株系的表型变化很大,使得统计分析复杂化。只有 pFUL:FUL:tFUL 株系和相应的 pFUL:FUL(AN):tFUL 株系之间的差异具有统计学显著性。
由于补偿能力依赖于转基因表达,我们对至少八个转基因株系在 12–16 期角果中的表达水平进行了测定(44),并将这些与观察到的补偿表型进行关联(图 4B)。对于每种组合(FUL 与 FUL(AN),FUL1 与 FUL1(ST),FUL2 与 FUL2(AN)),编码含有 Ser58 Thr59 蛋白的构建体表现出更陡的斜率和更好的表达水平与补偿能力的相关性,表明这些蛋白在角果中的功能性更高。值得注意的是,FUL1 和 FUL2(AN) 株系对 ful 角果的补偿能力远低于拟南芥 FUL(AN),尽管 FUL(AN) 和 FUL2(AN) 表现出相似的 AG-SEP 相互作用丧失和体外同二聚化丧失(图 3C–H 和补充图 S6 和 S7)。
为了测试 FUL(AN) 和番茄 FUL2(AN)/FUL1 是否在与 AG-SEP 相互作用或同二聚化能力上存在细微差异,我们在不同浓度的竞争性抑制剂 3-氨基-1,2,4-三氮唑(3-AT)下进行了酵母双杂交实验,并进行了额外的 EMSA 实验(补充图 S11 和 S12)。在酵母实验中,即使在低浓度的 3-AT 下,FUL(AN) 也无法与 FUL 或 FUL(AN)、SEP 和 AG 相互作用,与 FUL2(AN) 类似,表明两种蛋白的相互作用丧失程度相同(补充图 S11)。然而,在 EMSA 中,FUL(AN)-AG 相较于 FUL2(AN)-AG 显示出更强的迁移带(补充图 S12),这表明,依赖于条件和/或 DNA 的存在,FUL(AN) 可能比 FUL2(AN) 或 FUL1(ST) 更具与其他 MADS 结构域蛋白相互作用的能力,这可能解释了其在角果中表现出更好的功能性。
总之,我们在此展示,通过修改涉及特定蛋白-蛋白相互作用的氨基酸,可以生物学上分离多功能转录因子的功能,但生物复杂性使得在植物中的解释不那么直接。然而,这里观察到的轻微效应在农业中可能非常重要,例如在不影响开花时间的情况下防止芸薹属植物(68)的豆荚爆裂。
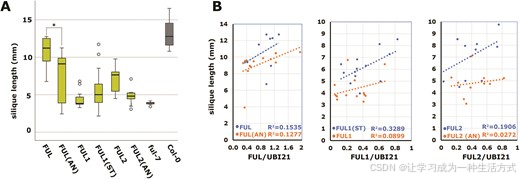
图 4. FUL、FUL1 和 FUL2 的生物活性依赖于它们的相互作用能力。(A) 在 ful-7 突变体背景下,不同 pFUL:(th)FUL:tFUL 转基因株系的平均角果长度。每个柱子代表至少 15 个不同转基因株系的数据。FUL 表示 pFUL:FUL:tFUL,FUL(AN) 表示 pFUL:FUL(AN):tFUL,依此类推。右侧的两个柱子表示野生型和 ful 突变体的对照。星号表示 FUL(AN) 和 FUL 株系之间的显著差异(P < 0.01),采用单因素方差分析(ANOVA)并进行 Tukey 事后检验确定。在此实验中,幼苗首先在平板上筛选,然后转移到岩棉上,与图 2C 中的实验相比,产生的植物角果略小。(B) 散点图显示了角果长度(y 轴)与相对表达水平(x 轴;ddCt (th)FUL/ddCtUBI21)之间的关系,展示了 (A) 中相同转基因株系的一个子集。每个线性趋势线的确定系数(R²)也包括在内。对于每种转基因组合,橙色点和相应的下方线表示 Ala58 Asn59 转基因株系,而蓝色点和相应的上方线表示 Ser58 Thr59 转基因株系。
识别出的多态性对 MADS 结构域蛋白-蛋白相互作用的特异性具有更广泛的重要性
对 FUL 相互作用特异性重要的多态性(Ser58Thr59/Ala58Asn59)位于 M- 和 I- 域的边界,这是先前与 MADS-MADS 二聚体特异性相关的区域(65)。为了评估该区域对相互作用特异性的普遍重要性,我们在 I 域中寻找了一些通常保守或特定类群的基序。I 区域从 1006 个不同植物物种中的总计 11966 个 MIKC 型蛋白序列中提取,并使用 MEME (48) 和 FIMO (49) 进行基序挖掘和计数。识别出了 18 个基序,主要是类群或谱系特异性的(补充图 S13)。有趣的是,最常见的、普遍保守的基序在开始时包含了位置 58 和 59(图 5A)。在这些位置,两个 Ser 残基最为常见,但包括 Ala58 和 Asn59 在内的几种替代残基也相对高频出现。该基序中的其他几个残基,如 Met62、Glu67、Arg68 和 Tyr69(具体位置取决于蛋白)表现出高度保守性。
为了获得该基序对蛋白-蛋白相互作用特异性普遍重要性的结构解释,我们使用 Alphafold2 (AF2) (69) 对 FUL、AG、SEP3、FUL1、FUL2、TAG1 和 TM3 的不同组合进行同二聚体和异二聚体的预测。此外,我们使用 PISA (54) 计算了所得结构的埋藏表面积和自由能(补充图 S14)。预测的结构与实验确定的 SEP3 DNA 结合域(64)和 SEP3 以及 SEP3-AG 的 K 结构域一致(70–72)。自由能计算预测 FUL、AG、TM3、FUL1 和 FUL2 的同二聚体比异二聚体更不稳定,而 SOC1 同二聚体得分较高,与其稳定的同二聚体形成一致。残基 58–59 (AN/ST) 位于 MADS 结构域的 C 端 β 链和 I 域的 N 端 α 螺旋之间的低概率灵活区域,预测不会在二聚化界面上与相邻单体直接接触(图 5B)。这表明,可能存在更微妙的变构效应,而这些效应在 AF2 模型中较难预测,对二聚体特异性的赋予具有重要作用。
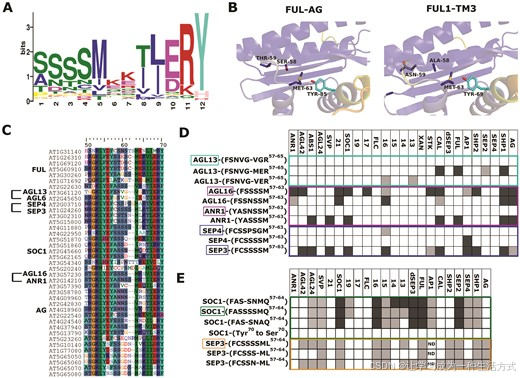
图 5. 鉴定出的多态性对 MADS 结构域蛋白-蛋白相互作用的特异性具有更广泛的重要性。(A) 在 I 区域中最丰富的基序,通过 MEME 基于 11966 个 MIKC 型蛋白鉴定。序列标志的第 1 和第 2 个位置分别对应 MADS 结构域蛋白的氨基酸位置 58 和 59。(B) FUL-AG 和 FUL1-TM3 的代表性 Alphafold2 二聚体,显示了对二聚体特异性重要的 AN 和 ST 残基的位置。蛋白质按概率用颜色编码,从蓝色(较高概率)到黄色(较低概率)。残基位于 M- 和 I- 域之间的环中,不直接参与二聚界面的形成。展示了对二聚体稳定性和/或特异性可能重要的甲硫氨酸和酪氨酸残基,伴侣单体的酪氨酸残基以青色显示。残基以棍状显示,并按原子颜色编码和标记。(C) 拟南芥 MADS 结构域转录因子序列的比对,展示了包含识别基序的蛋白部分。用于测试该基序重要性的蛋白已标出。(D, E) 用于测试 Met63/Tyr70/环假设的构建体。方框内标注的蛋白名称表示野生型序列基序;其他则为交换或修改。每列表示使用酵母双杂交测试的 MADS 结构域蛋白。详见补充图 S18–S23。深色框表示双向相互作用(AD 对 BD 和 BD 对 AD);浅色框表示仅单向相互作用;白色框表示无相互作用;ND = 未确定。SEP3 构建体在 (E) 中仅测试了一个方向,因为 BD-SEP3 自激活。使用未突变的 SEP3 时,我们使用缺少部分 C 末端的 dSEP3 版本,避免自激活。数字 21 表示 AGL21,19 表示 AGL19,依此类推。XAN=AGL12(XAL1)。
在 I 域二聚螺旋中,二聚体配对的两个单体间的主要相互作用由 Tyr70(单体 A)和 Met63(单体 B)形成。这是一种有利的甲硫氨酸-芳香族相互作用,在所有相互作用伙伴中保守,AG 除外。图 5B 显示 Thr59 不直接影响 Met-Tyr 配对,因此无法轻易解释其对二聚体特异性的影响。位置 58–59 处于邻近二聚螺旋(位置 58–62,邻近 Met63)的环区。由于环区结构预测难度较大,我们假设环中的氨基酸变化可能会对二聚界面的 Met–Tyr 相互作用产生微妙影响,从而使该相互作用不稳定或稳定,并允许仅特定的相互作用。AG 具有 Ala63 而非 Met63,可能解释了为什么 FUL-AG 需要 Thr59,而其他相互作用也能容忍 Ser59。有趣的是,SOC1 的环序列不同,少了一个氨基酸,可能通过更刚性的环或不同的环构象来补偿 Thr59/Asn59 FUL 突变,从而稳定二聚界面。由于 AF2 的结构预测尚不能明确预测 MADS 结构域转录因子的二聚体特异性和相互作用模式,我们决定实验性地测试环-Met-Tyr 假设。
在拟南芥 MIKC 型蛋白中,我们选择了环残基已发生分化并具有不同相互作用特征的旁系同源物/近缘同源物(55):AGL16/ANR1、AGL16/AGL13 和 SEP3/SEP4(图 5C)。设计了构建体,其中一个或几个氨基酸被交换为近缘同源物的残基(图 5D 和补充图 S15–S17)。在 AGL16/ANR1 配对中,交换氨基酸显著改变了相互作用能力,ANR1 获得了多种相互作用,而 AGL16 失去了大部分相互作用。突变 AGL13 蛋白也表现出相似的相互作用增强,特别是引入 Met63 替代 Val63 时。SEP4 在引入 SEP3 环中的氨基酸后并未表现出额外相互作用,但鉴于 SEP3 和 SEP4 的 K 结构域之间的氨基酸差异较大,这或许并不令人意外(补充图 S17)。
为进一步测试环长度以及 Met63 和 Tyr70 的重要性,我们在 SEP3 和 SOC1 蛋白中生成了额外的突变构建体(补充图 S18–S19)。在 SOC1 中通过引入一个氨基酸(Ser60)延长环明显影响了几个特定相互作用,例如 BD-SOC1_S59-62 与 AD-AGL13 和 AD-AGL14 的相互作用丧失,且与 AD-SEP2 的相互作用减弱。有趣的是,具有常规短环长度但 Ala63 替代 Met63 的 SOC1 版本影响了相同的相互作用,且增加了 AD-SOC1_Ala63 与 AGL19 的相互作用丧失(图 5E 和补充图 S19–S23)。因此,延长环长度或替换 Met63 都会降低 SOC1 与特定相互作用伙伴的结合亲和力。将 Tyr70 更换为 Ser70 对相互作用能力产生了最显著的影响,几乎消除了所有相互作用,表明 Tyr70 确实是 MADS 结构域蛋白二聚化的关键残基(图 5E)。
我们构建了两种不同的 SEP3 版本,以测试在 SEP3 中缩短环是否会增强相互作用,观察到 AD-SEP3_S59S60N61(与 SOC1 环序列完全相同)增加了与 BD-SEP3、BD-SEP4 和 BD-CAL 的相互作用。然而,克隆 AD-SEP3_S59S60S61 未获得这些相互作用,反而失去了一些,表明在 SOC1 中,Asn61 除了短环外也很重要(图 5E 和补充图 S19–S23)。
总的来说,我们利用番茄功能细化的共同同源基因 FUL1 和 FUL2,确定了对蛋白-蛋白相互作用特异性至关重要的残基,并进一步研究这些残基,识别出一个对 MADS 结构域蛋白相互作用特异性广泛重要的蛋白基序。在二聚界面处 Met–Tyr 相互作用的稳定性取决于相邻环的组成,该环仅允许有限的或多种二聚化伙伴。
讨论
我们利用番茄 FUL 共同同源基因 FUL1 和 FUL2,它们在一定程度上已经发生了功能细化,从而深入了解拟南芥 FUL 在蛋白质水平上的不同功能。我们发现,FUL 在花序或角果中形成的组织特异性复合体不同,其结合的靶基因集合也有所差异。通过将番茄 FUL1 和 FUL2 序列中的多态性与其相互作用模式的变化关联,我们鉴定出一个决定其与 AG 和 SEP 相互作用能力的 2 个氨基酸基序,但并未影响其与 SOC1 的相互作用。通过鉴定该基序,我们进一步证实了 MADS–MADS 相互作用特异性的最重要区域是 I 域(此前已有研究提出)(63,64),并提出了该基序如何通过位于 M- 和 I 域边界的结构影响相互作用亲和力的假设。我们展示了该基序对其他几种 MADS–MADS 相互作用也很重要,但这并不是特异性的唯一决定因素。因此,未来的研究需要补充更多细节,以获得对相互作用特异性决定因素的更全面理解。在多功能的拟南芥 FUL 蛋白中修改该 2 个氨基酸基序会特异性地降低其在果实中的功能,从而在植物体内至少部分地实现了特定功能的修饰。然而,生物学效应比预期的要温和,这表明将体外实验结果转化为复杂的植物体内情境仍具挑战性。然而,即使是相对适度的定量差异在作物育种中也很有价值,而启动子区的修饰已成功用于实现预期的定量效果(15,16)。在蛋白质水平上解析功能的能力也可能具有重要价值,代表了一种可行的方法来应用蛋白质工程改善作物。
除了开发新方法外,FUL 的花序和角果功能的分离还提供了关于复合体特异性靶基因调控的新见解。像拟南芥 FUL 这样的多功能蛋白具有广泛的表达模式,可以与多种其他蛋白相互作用,以履行其在不同植物组织中的功能。然而,在每种细胞类型的背景下,仅存在一部分相互作用伙伴,并且根据可以形成的细胞特异性复合体,不同的靶基因集合会被调控。MADS 结构域蛋白根据细胞类型特异性复合体调控不同靶基因集合的特性构成了著名的花器官发育 ABCE 模型的基础,但由于所有 MADS 结构域蛋白都结合 CArG-box,靶基因集合如何区分仍然是一个未解之谜(20,73,74)。最近的研究表明,CArG-box 中的 CC 和 GG 核苷酸对二聚体与 DNA 在大沟中的最佳接触至关重要,而序列中间的 A 链决定小沟宽度,这对于 MADS 蛋白的结合也具有决定性作用(22,23)。已证明 SEP3 具有较高亲和力结合带有 A 链和窄小沟的 DNA,这一特性依赖于 MADS 结构域蛋白 N 端的高度保守的精氨酸残基,该残基延伸至小沟(22)。AG 缺乏这一精氨酸可能表明它对 DNA 形状有其他需求。AG 结合的 CArG-box 5′ 和 3′ 侧的 A/T 伸长区的重要性可能与此相关(图 1H)。除了 DNA 碱基和形状识别外,四聚体结合两个 CArG-box 的亲和力可能取决于它们的方向和距离(22–24)。最后,复合体特异性共因子的募集可以影响 DNA 结合和靶基因调控。植物体内的 MADS 结构域复合体的下拉实验表明,几种共因子,包括共抑制剂/激活剂和染色质重塑因子,可能与 MADS 结构域复合体相关联(17)。关于 MADS 结构域蛋白的更具变异性的 C 端结构域的功能仍知之甚少。该结构域不参与 DNA 结合,但可能与多种共因子相互作用,从而特异性激活或抑制靶基因表达,进而促成复合体特异性调控(23,74)。拟南芥 FUL 在角果中的功能性优于番茄 FUL2,可能也由于其 C 端特异性相互作用(此外还有其同二聚化能力)。总之,不同 MADS 结构域复合体的靶基因特异性取决于多个特征的组合,我们的 DAP-seq 方法进一步揭示了 DNA 结合基序中的复合体特异性变异,这些特征有助于碱基和形状识别。
虽然 FUL–FUL、FUL–SOC1 和 FUL–AG–SEP 复合体的靶基因大多是重叠的,但可能是特定靶基因决定了组织特异性,可能在植物体内取决于相互作用亲和力和表达水平。例如,在拟南芥分生组织中,SOC1 在过渡至开花时的表达量高于 FUL(TraVa 数据库 (75)),并且由于 FUL–SOC1 复合体非常稳定(图 1A 和补充图 S7),FUL–FUL 复合体可能在该组织中不存在。然而,SOC1 不在雌蕊/角果中表达,可能是存在于花分生组织中心的 FUL–FUL 复合体(从第三阶段开始),在某个阶段特异性抑制 SPT/HEC 基因,此时其他高亲和力的相互作用伙伴(如 SEP3)较少。拟南芥角果的分布模式受到严格调控,以确保脱离区的发育(77,78),而番茄果实不需要这种模式,因为它不需要通过破裂来释放种子。番茄 FUL1 和 FUL2 无法同二聚可能与这种模式的需求丧失有关。然而,番茄 FUL2 在果实生长早期具有功能,而 FUL1 则没有(33)。FUL2 的这一功能可能取决于其与 TAG1 的相互作用,而 TAG1 在此阶段表达(番茄表达图谱 (79)),而 FUL1 仅在果实成熟开始时表达,这与其无法与 TAG1 相互作用一致。有趣的是,FUL1 和 FUL2 都在果实成熟开始时与 SEP3 的同源物 RIPENING INHIBITOR (RIN) 相互作用(80),而 FUL1 无法与拟南芥 SEP3 相互作用。因此,FUL1 和 RIN 的共同进化确保了它们的特异性相互作用,FUL1 的功能细化程度低于在拟南芥中的异源实验所预测的。将 MADS 结构域蛋白的蛋白序列差异与蛋白-蛋白相互作用谱和功能特征的差异联系起来仍是一个挑战,但我们发现 Met/Tyr/环基序作为 MADS 结构域蛋白-蛋白相互作用特异性的更通用的因素,将有助于未来对 MADS 结构域家族的研究。


















 2万+
2万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









