








Bacterial coexistence driven by motility and spatial competition
DOI:https://doi.org/10.1038/s41586-020-2033-2
写在前面:
2020年2月19日发表在Nature的一篇文章。这是近期读过的最有意思的文章,没有之一。
生物多样性的形成与维系机制始终是生态学研究的重难点之一。本公众号长期致力于推送相关的研究成果,无论是理论研究还是实验研究,对此不再赘述。比较早的开展系统研究并提出standard理论的代表,应该就是达尔文了吧?及至后来,现代生态学之父哈钦森先生也很疑惑,动物的多样性为什么这么高?(详见哈钦森1959年的演讲文章:Homage to Santa Rosalia or why are there so many kinds of animals?--太长了,至今还没有翻译完,争取今年5月17之前弄完)
关于coexistence,比较经典的文献之一,就是Peter Chesson于2000年发表在Annual Review of Ecology and Systematics的一篇综述(也被这篇Nature文章所引用-参考文献24:Mechanisms of maintenance of species diversity)。所谓stable coexistence,如果能够长期实现,低密度或者低丰度的组分需要有机会翻身,而不是因为本来就少、然后很快被灭掉。也就是这篇Nature文章里提出的,需要等级置换hierarchy inversion。
Stable coexistence的形成主要包括两种途径:1)equalizing,均等机制(或者求同),即,不同的species将彼此间的平均适合度最小化;2)stabilizing,维稳机制(或者存异),即,不同的species努力增强negative的种内互作、而不是增强negative的种间互作。(啊,忽然想到,我们伟大的周总理,N多年前(1955年万隆会议)就提出,世界人民的友好共存是要“求同存异”的~你看,人,终归也只是一种动物而已)
具体来说,Stabilizing这一方式,主要通过如下机制实现stable coexistence,例如资源分配、频次依赖性的捕食作用,以及其他依赖于群体密度和环境因子时空波动的若干机制。Equalizing这一方式,则主要是通过如下机制实现stable coexistence,即降低物种彼此间的适合度差异,这一结果实际上会削弱stabilizing相关机制的效应。
本文是近期发表在Nature的一篇文章,非常有意思,应该算是stabilizing模式形成的coexistence。以两株大肠杆菌为材料,研究了运动能力与空间竞争共同驱动下的细菌共存,并从多个水平对相关机制做了解析。本研究将两个菌株按不同比例混合共培养时,任一菌株在低丰度时,都会竞争超过另一菌株,即通过主动隔离和空间排除实现等级置换。具体来说,就是,运动速度快(生长速度慢)的小群体,会快速从起始位点向远处移动,从而比生长速度快(运动速度慢)的大群体更早地占据了远处的空间,并消耗掉了相应的资源,导致生长速度快的群体无法再向远处迁移;反之,生长速度快(运动速度慢)的小群体,会迅速消耗掉两群体接触区域的资源,从而驱逐运动速度快(生长速度慢)的大群体离开。所以,总的来说,就是对于存在“生长-迁移”权衡的细菌,它们通过这两个过程的反复增强作用,实现了有效的空间隔离和群体共存:即,生长速度快的群体,总是会倾向于占据原始位置的空间,而运动速度快的群体,则总是倾向于占据远离原始位置的空间。
文章简介:
1. 共培养的两个野生菌株存在竞争等级的置换
本研究使用了两株Escherichia coli的菌株,分别是A和B—分离自同一宿主的粪便样品。将A和B按照不同比例混合(比例为R0),接种到液体培养基和软的固体培养基,培养2.5~4天后,统计两群体新的比例R,计算比例的变化倍数:R/R0。结果发现,液体摇瓶培养条件下,无论起始R0为多少,R均增大约1000倍,这与两个菌株分别单独培养时的情况吻合;而在固体培养条件下,R0较低时,R值增大;R0较高时,R值减小(图1a-b)。即,这两个菌株,都能够在自身起始丰度较低的情况下,最终竞争超过对方。也就说,要实现共存,需要A和B在生长的过程中,完成等级置换,双方趋向于一个能够共存的比例;否则一方的灭绝,就会导致共存无法实现(图1c-d)。
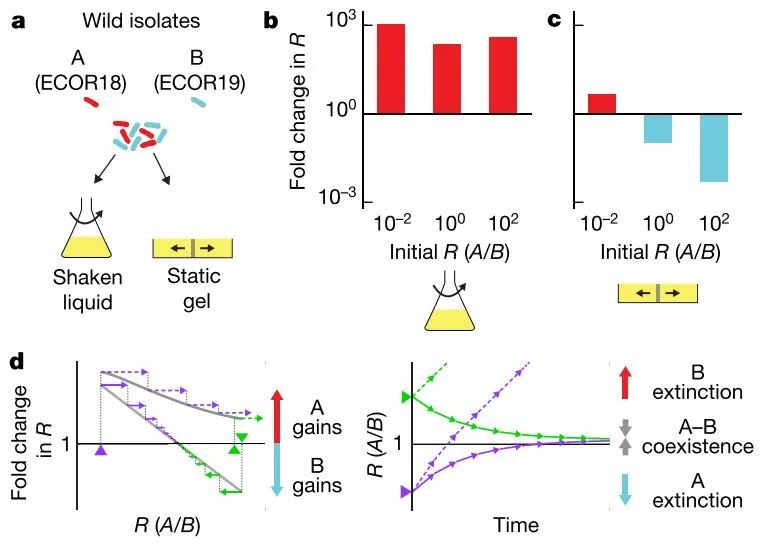
图1. 共培养的两个野生菌株存在竞争等级的置换
2. 生长-迁移权衡对空间隔离的贡献
为什么会出现上述现象呢?本研究发现,将A和B各自单独培养在固体培养基时,4天之内,A和B均会长满平板;而当混合培养时,点样在平板中间位置时,A会留在接种圆点,并且形成较小的菌落,B则会在远离A的、边缘区域,形成较大菌落(图2a)。当把两菌点样在平板边缘位置时,A会留在边缘位置,B则依然出现远离A的区域(现在等于是大约到了平板中间位置)。其他一些配对菌株也发现类似现象。
已知,驱动细菌互斥的一个重要因素可能是细菌分泌的抑制物,但是本研究并没有检测到类似的抑制物。然而,研究人员发现,比较有意思的是,出现上述现象的配对菌株,往往存在生长和迁移的权衡,即,一方具有生长优势时,另一方往往具有迁移优势(图2b)。实验确实也发现,不存在这样的权衡或者较弱的存在时,菌株对就没有呈现出类似的空间隔离。
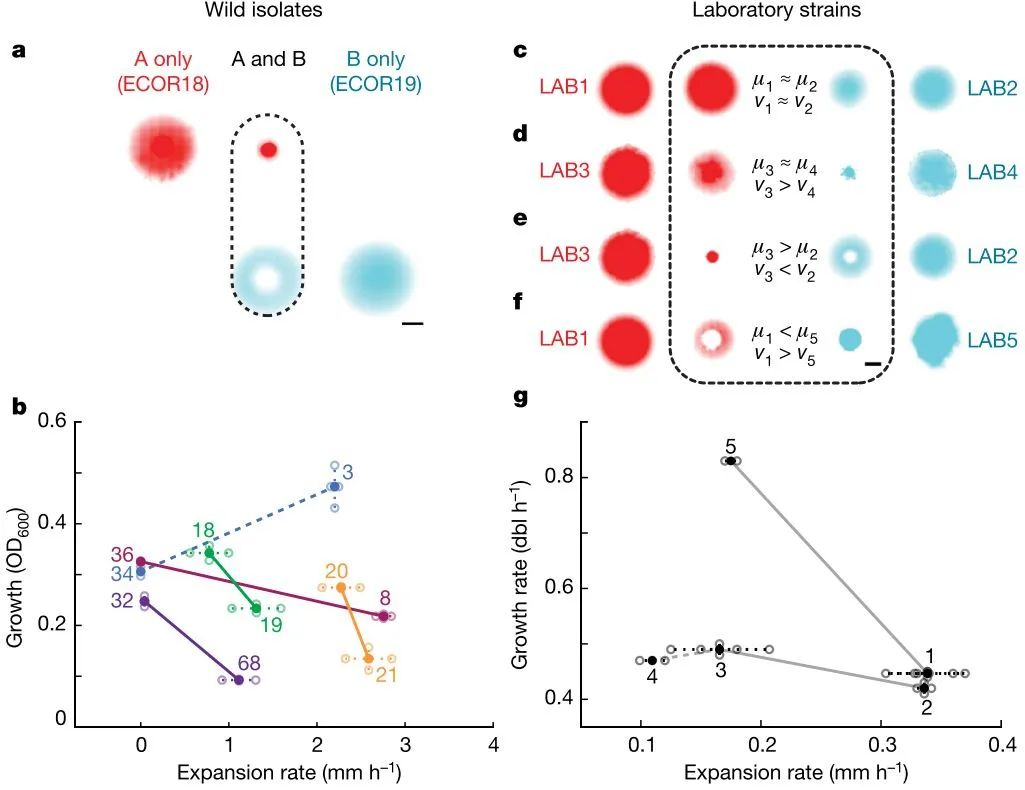
图2. 生长-迁移权衡与空间排除
为了进一步直接验证生长-迁移权衡的重要性,研究人员在实验室构建了突变体菌株,改变菌株的生长速度和运动速度。结果发现,当突变体菌株的生长和运动速度相当或者接近时,空间排除现象没有出现(图2c,2g);当运动速度差异大、生长速度接近时,出现了边缘排除现象、没有出现中心排除现象(图2d,2g);当生长速度和运动速度均差异明显、存在权衡时,同时出现了边缘排除和中心排除现象(图2e,2f,2g)。
3. 菌落发展动力学解释生长-迁移权衡的作用原理
对存在生长-迁移权衡的菌株,研究人员仔细研究了其菌落发展动态(图3a),发现其菌落模式可以用Keller-Sigel细菌运动模型来预测(图3b)。研究人员结合Monod生长方程、营养消耗等,对上述模型进行了延伸,得到了Monod-Keller-Sigel模型,可以更准确地预测群落的发展变化。
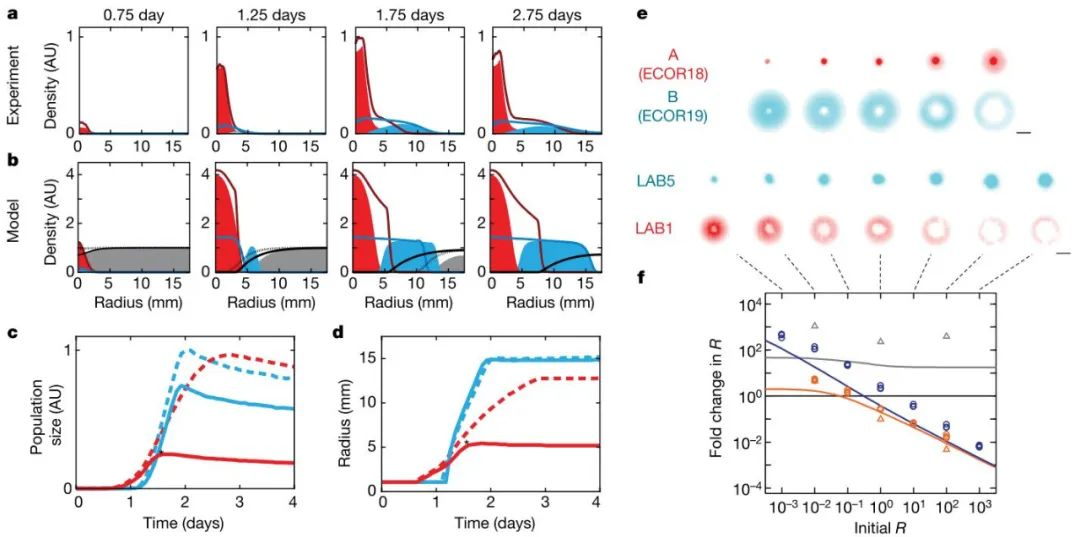
图3. 竞争性迁移的围追堵截效应
通过对菌落变化的动态过程分析发现,一方面,运动速度快的菌株会限制运动速度慢的菌株留在点样原始位置:运动速度快的菌株在前进的过程中,会消耗掉沿途的资源,大约在1.6天左右完全消耗掉,从而阻断了运动速度慢的菌株、使其不能向外扩散(图3)。另一方面,运动速度慢的菌株,因其生长速度快,也在主动上占据了原始位置,限制了生长速度慢(幸好运动速度快)的菌株留在这里、驱使其赶紧向远处迁移。总的来说,与以往的预设空间生态位不同,本研究发现的这两株菌株,其生态位的形成属于后发性质的。
4. 从打破共存的角度反向验证生长-迁移权衡的重要性
既然上述分析证实了生长-迁移的权衡及其对群体丰度的负依赖性,驱动力群体的最终共存,可以推测,如果打破这样的平衡,共存应该也会消失。举例来说,如果提高运动速度快的群体的丰度,可能会导致其在向远方移动时、没有足够的空间来供养自己,最终失去对相应空间的占据;而生长速度快的群体,当其丰度较高时,可能会导致其在生长时没有足够的资源支撑,最终也无法较好地占据相应的空间。模型预测和实验结果均证实,当对群体大小、接种密度等进行改变后,确实会导致共存的崩溃(图4)。
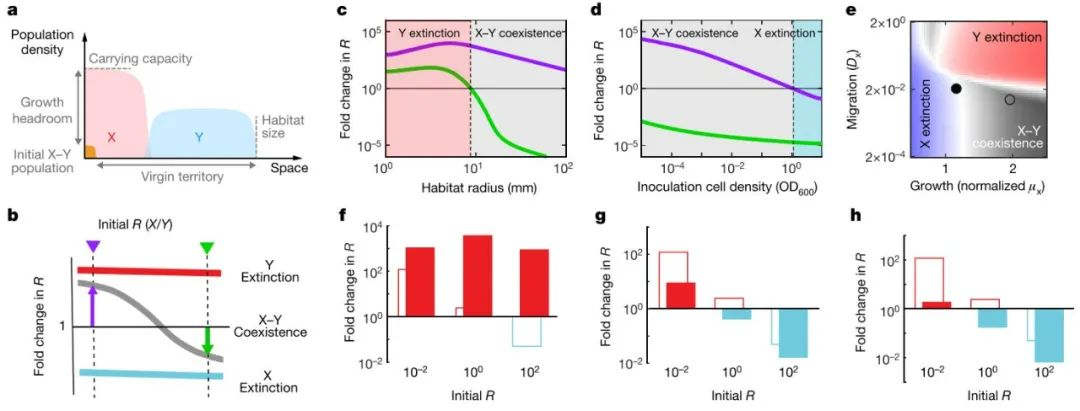
图4. 共存的稳定性及其界限
5. 基因组学证据揭示菌株运动能力差异的根本原因
为了进一步揭示,两个菌株运动能力差异的本源,研究人员对菌株继续了基因组测序。结果发现,基因组水平的一些变异可能可以提供一些解释。A和B菌株的很多个运动相关的基因上,检测到50多个氨基酸替换。三维的单细胞追踪揭示,不同菌株的游泳速度存在很大的差异,并且与其群体的迁移速度具有很好的相关性。
6. 小结
细菌的运动对细菌来说是非常重要的能力与特征,相关的研究非常多(记得以前在生态中心时,有幸听过刘双江老师的一次报告,就是关于细菌运动的,讲得特别有意思)。然而,本文发现,运动能力对于细菌的多样性维持,可能也具有非常重要的作用。这一发现,无论是对于细菌的群落多样性研究、还是其他生态系统内生物多样性的研究,无论是理论研究、还是实验研究,都具有很重要的参考意义。
猜你喜欢
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
文献阅读 热心肠 SemanticScholar Geenmedical
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份,另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。

学习16S扩增子、宏基因组科研思路和分析实战,关注“宏基因组”
 点击阅读原文
点击阅读原文

















 1407
1407

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









