







 该博客介绍了蛋白质三维结构相关知识。涵盖蛋白质结构层次,包括一至四级结构;阐述研究构象的方法,如X - 射线衍射法等;分析多肽链主链折叠限制、二级结构类型特点;还讲述蛋白质折叠过程、二硫键作用及体内折叠辅助因素,以及四级结构的特点和优越性。
该博客介绍了蛋白质三维结构相关知识。涵盖蛋白质结构层次,包括一至四级结构;阐述研究构象的方法,如X - 射线衍射法等;分析多肽链主链折叠限制、二级结构类型特点;还讲述蛋白质折叠过程、二硫键作用及体内折叠辅助因素,以及四级结构的特点和优越性。
6生物化学与分子生物学——蛋白质的三维结构
引论
蛋白质是由多肽链折叠而来的,具有特殊的空间结构和生物学功能
蛋白质的空间结构即蛋白质的构象,指蛋白质分子所有原子在三维空间(x,y,z)的位置
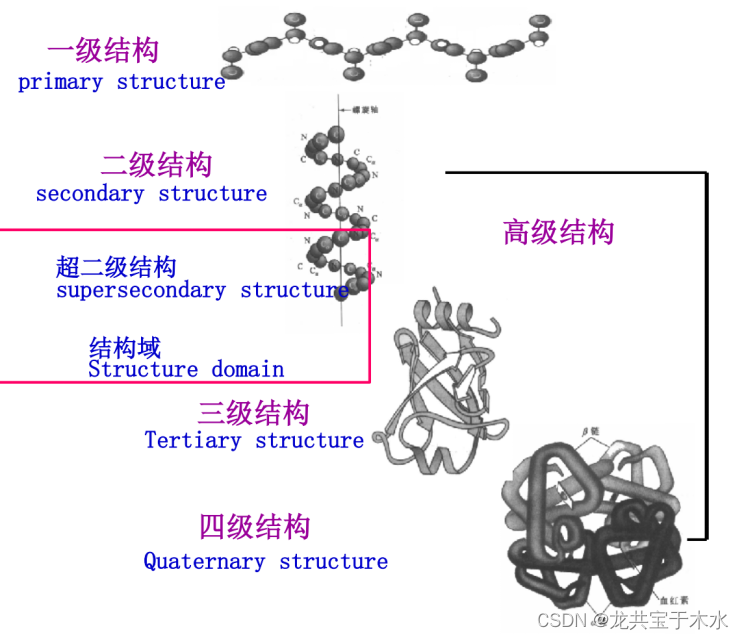
蛋白质结构的层次
1°结构:蛋白质多肽链中氨基酸残基的排列顺序
2°结构:蛋白质多肽链主链的折叠方式
3°结构:二级结构基础上蛋白质一条多肽链的折叠
4°结构:寡聚蛋白质各亚基的空间排布(并不是所有蛋白质都具有)
研究蛋白质构象的方法
X-射线衍射法(目前用得最多,冷冻电镜用来分析整合膜蛋白的立体三维结构)
核磁共振(NMR)法(用来分析水溶液中蛋白质三维结构-更接近生物体内的蛋白质)
圆二色谱(CD)法
荧光偏振
拉曼光谱
扫描隧道显微术(STM)
维持蛋白质三维结构的作用力
维持蛋白质三维结构的作用力主要是一些弱的相互作用(非共价力)
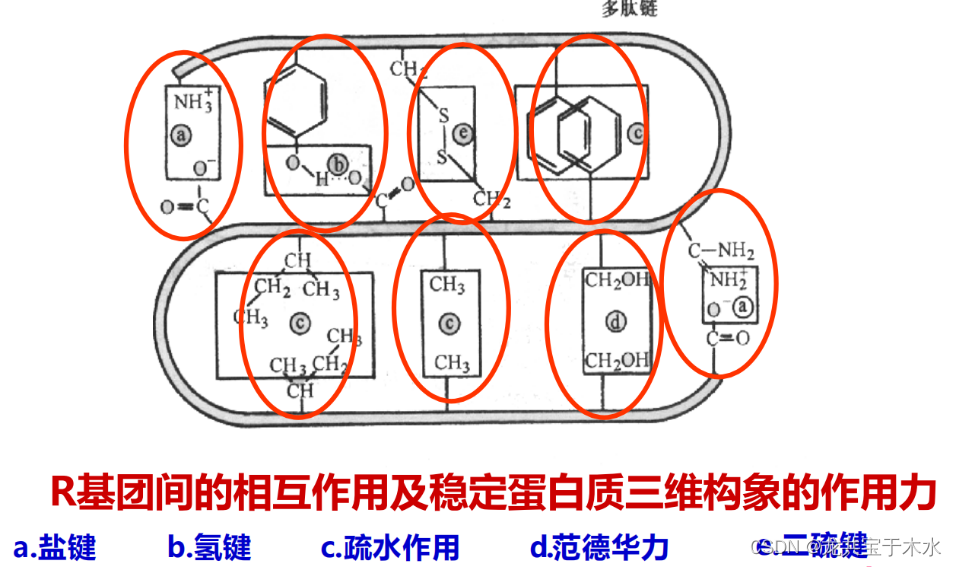
本章重要内容(参考)
稳定蛋白质三级结构的作用力
多肽主链折叠的空间限制(二面角)
蛋白质二级结构的类型和特点
主要纤维状蛋白质的结构特点
多肽链主链折叠的空间限制
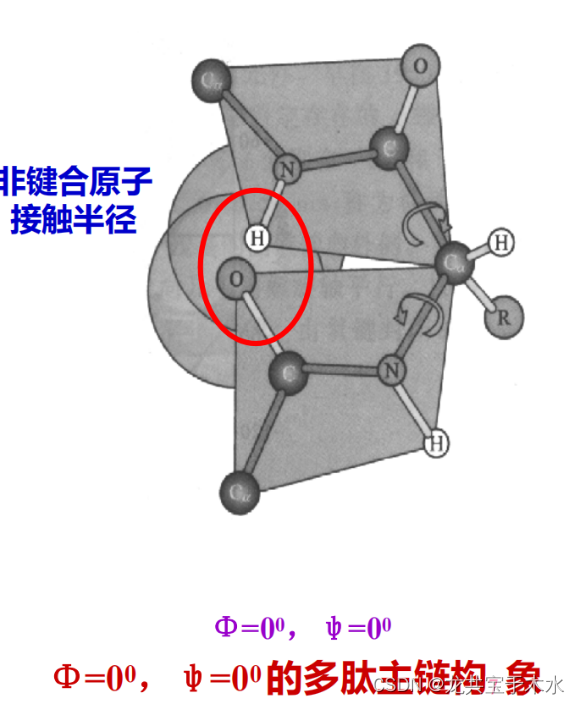
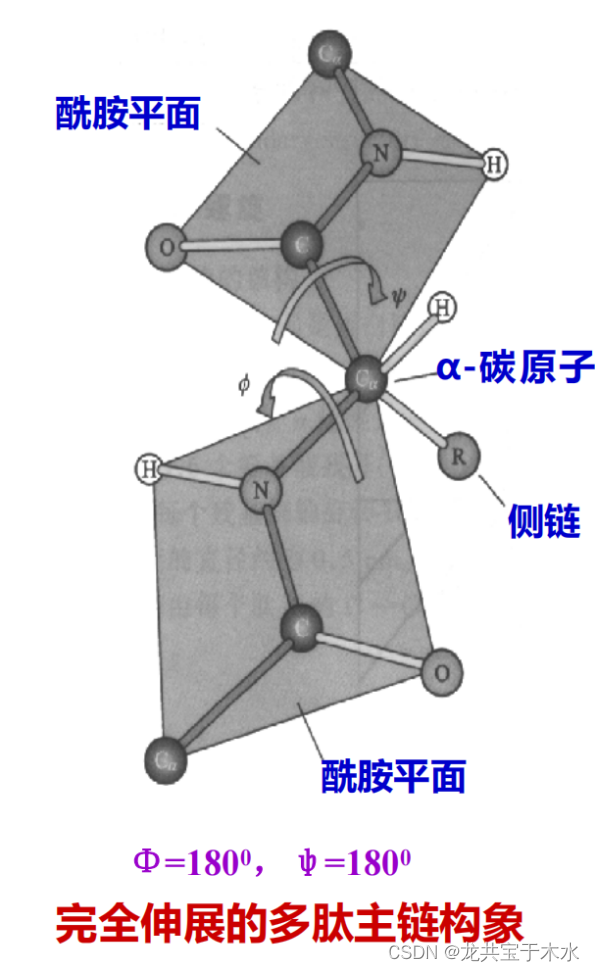
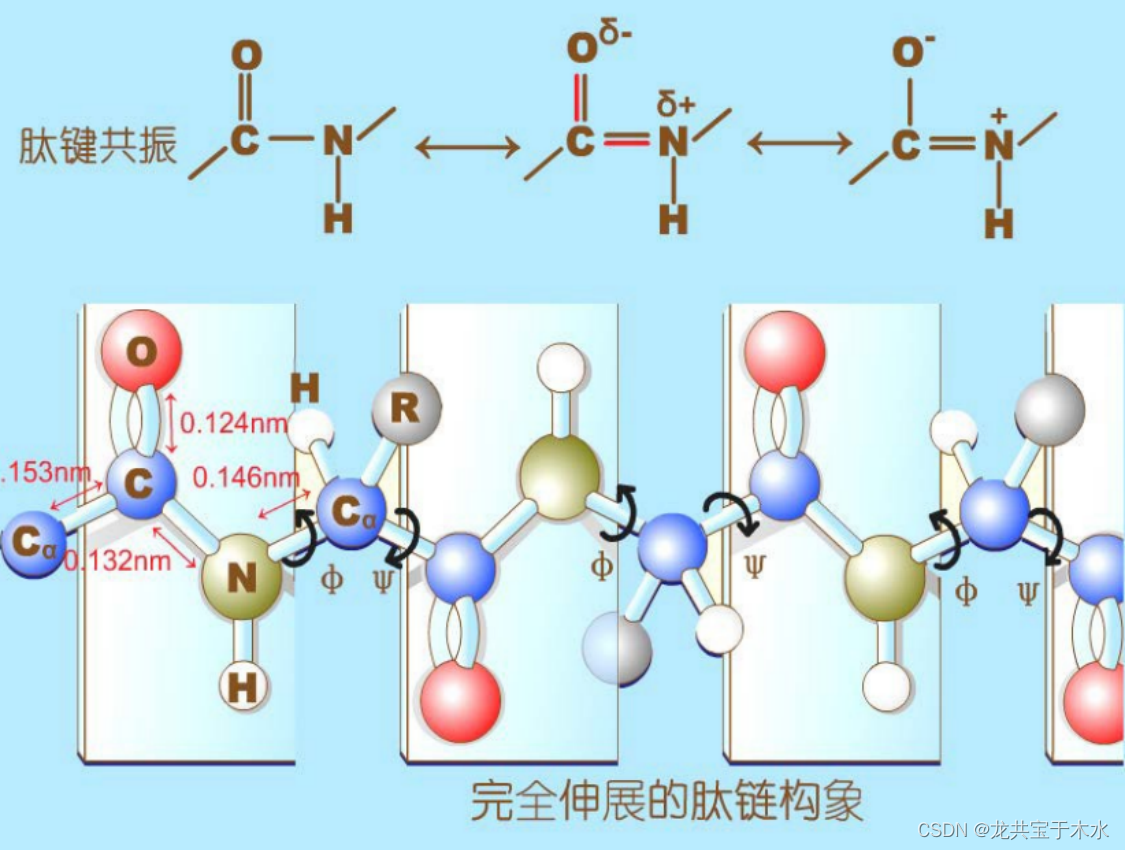
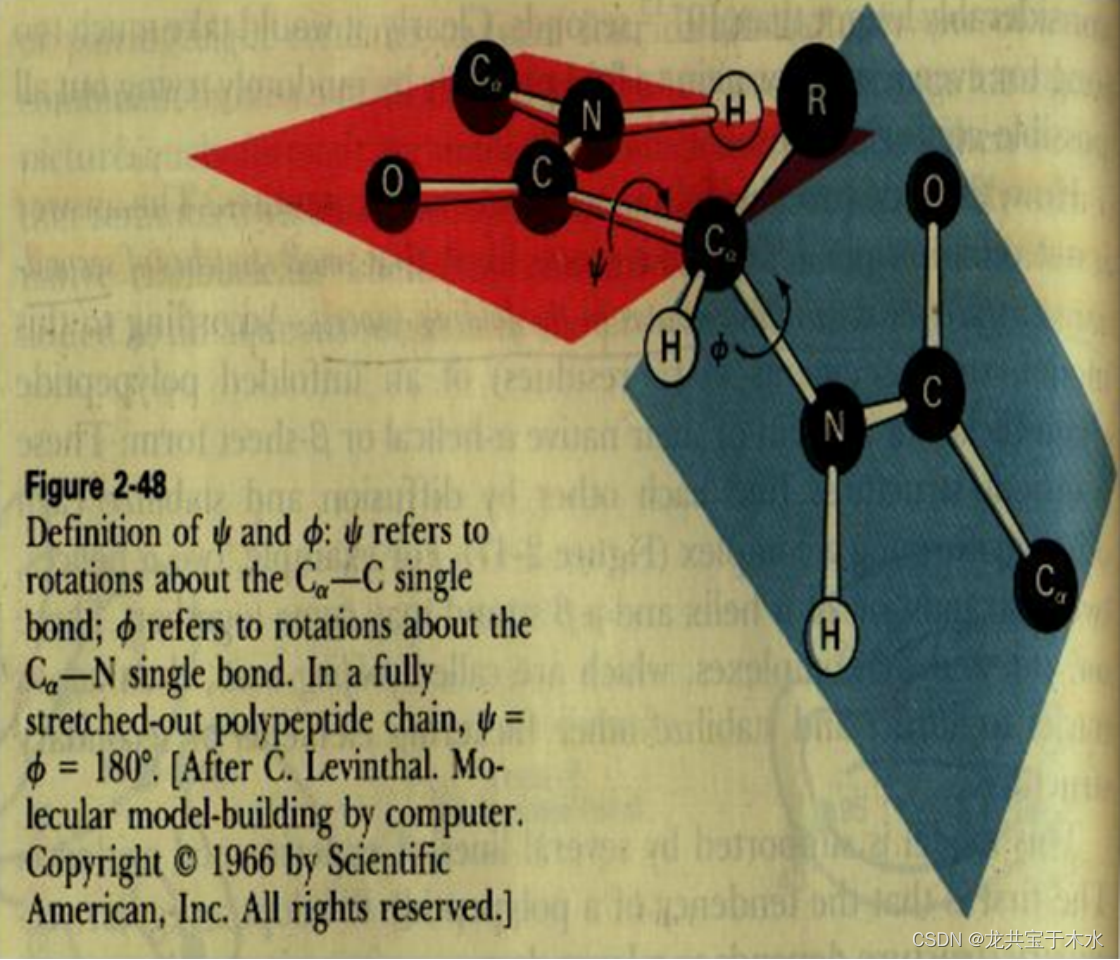
酰胺平面与α-碳原子的二面角(Φ和ψ)
两个氨基酸残基间的肽键由于共振相互作用导致其键本身是单键但具有部分双键的性质,所以组成肽键的这四个原子和两个α-C形成了一个刚性的酰胺平面,这个平面中只有α-C和羧基的C之间的单键能自由旋转
二面角:两相邻酰胺平面之间,能以共同的Cα为定点而旋转,绕Cα-N键旋转的角度称Φ角,绕C-Cα键旋转的角度称ψ角,Φ和ψ称作二面角,亦称构象角
实际上二面角的旋转是受到限制的:它的旋转会改变蛋白质多肽链中非共价键与原子之间的距离,当两个非共价键与原子之间的距离过近的时候,会产生一种斥力使得此构象不稳定
由此推导出二面角的旋转存在两种特殊情况
-
-
-
-
- 当Φ和ψ都是0°,即二面角平分由这个α-C上的H与R基团之间形成的角时。但实际上,此种构象是不存在的,因为酰胺N上的H与羰基上的O之间距离过近,产生斥力
-
-
-
-
-
-
-
- 还有一种充分伸展的构象即Φ与ψ都是180°,此种构象理论上是能量最小,最稳定的一种状态
-
-
-
可允许的Φ与ψ值:Ramachandran构象图
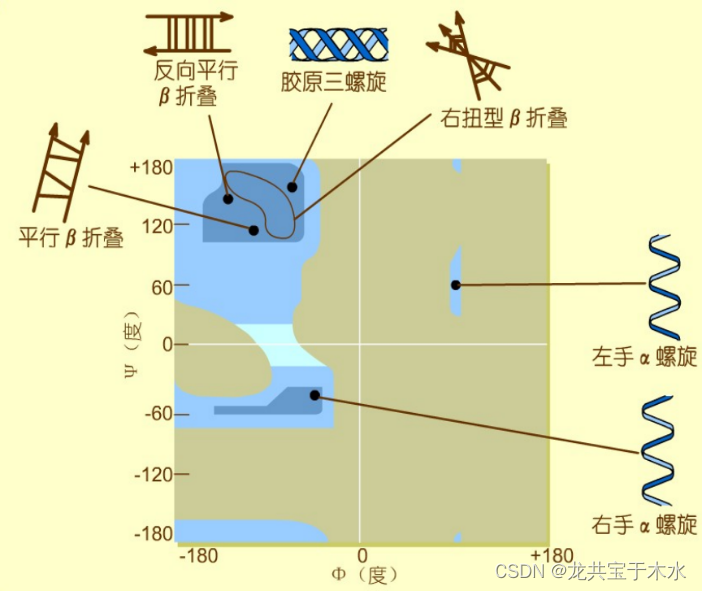
以横坐标为Φ的角度,纵坐标为ψ的角度,得到图画中不同颜色标注的部分,颜色最深的部分就认为是空间结构稳定的一种构象,颜色稍微淡一点的被认为可以形成这样的二面角(不稳定),通过此图可以确定多肽链中所有二面角的可能性,进而得到它的稳定空间结构
蛋白质的二级结构
定义
蛋白质的二级结构(secondary structure)指肽链主链不同区段通过自身的相互作用,形成氢键,沿某一主轴盘旋折叠而形成的由氢键维系的局部规则构象,是蛋白质结构的构象单元
主要有以下类型
-
-
-
-
- α-螺旋(α-helix)
-
-
-
-
-
-
-
- β-折叠(β-pleated sheet)
-
-
-
-
-
-
-
- β-转角(β-turn)
-
-
-
-
-
-
-
- 无规则卷曲(non-regular coil)
-
-
-
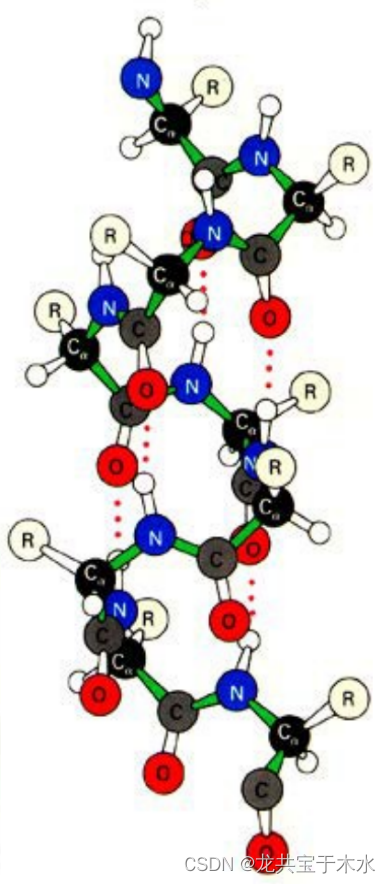
α-螺旋
特征
-
-
-
-
- 每隔3.6个残基螺旋上升一圈,螺距0.54nm
-
-
-
-
-
-
-
- 螺旋体中所有氨基酸残基R侧链都伸向外侧
-
-
-
-
-
-
-
- 每个氨基酸残基的>N-H与前面第三个氨基酸残基的>C=O形成氢键,肽链上所有的肽键都参与氢键的形成,取向几乎都平行于螺旋轴
-
-
-
-
-
-
-
- α-螺旋是蛋白质的一条多肽链或者多肽链的一个部分以假想的一个主轴为中心,以右手螺旋的方式形成的螺旋,此螺旋每隔3.6个氨基酸残基,就上升一圈,一圈的高度是0.54nm,形成螺旋的所有氨基酸残基的侧链基团都伸向这个螺旋的外侧。每个氨基酸残基的酰胺的H会和前面第三个氨基酸残基的羰基氧之间形成氢键(实际上多肽链中所有肽键都参与氢键的形成,而且氢键的取向与假想的中间轴是平行的)
-
-
-
形成原因
-
-
-
-
- 与AA组成和排列顺序直接相关
-
-
-
-
-
-
-
-
- 即需要氨基酸之间不能有强的静电作用,较大的R基团,同时还需要氨基酸有足够的分子大小形成稳定的二面角
-
-
-
-
-
-
-
-
- 丙氨酸:多聚丙氨酸在溶液中就很容易形成α-螺旋;异亮氨酸:侧链的分支很大,就不容易形成α-螺旋;多聚赖氨酸在pH为中性的溶液中就不能形成α-螺旋,因为此时赖氨酸带正电,电荷相互排斥,就不能能形成螺旋,但如果将多聚赖氨酸的溶液pH调成12时,此时不带电,即可形成螺旋
-
-
-
-
-
-
-
-
- 由于多聚丙氨酸的主链结构相对简单,相邻的主链氨基基团之间没有明显的碰撞或空间阻挡。 这种结构特点使得多聚丙氨酸的主链能够形成紧密的排列,从而有利于形成稳定的α螺旋结构。 在α螺旋中,多聚丙氨酸的主链呈螺旋状排列,形成了一个稳定的螺旋结构
-
-
-
-
-
-
-
-
- α-螺旋结构遇到脯氨酸Pro,因为脯氨酸是亚氨基酸,它的氨基形成了一个吡咯环,所以α-C和N之间会限制这个键的自由旋转;甘氨酸中的侧链只有一个H,分子量太小不易形成约束,也不容易形成α-螺旋
-
-
-
-
-
-
-
-
- 由于静电排斥不能形成链内氢键,从而不能形成稳定的α-螺旋,R基较小且不带电荷的氨基酸有利于α-螺旋的形成,R基越大,如异亮氨酸,不易形成α-螺旋,脯氨酸终止α-螺旋
-
-
-
-
-
-
-
-
-
- 甘氨酸 不利于形成α螺旋的原因是它的侧链太小,即自由度太大,与它有关的二面角变化太大,无法满足α螺旋形成所必需的一个条件——具有相对固定的 二面角
-
-
-
-
多态性
-
-
-
-
- 一般情况下α-螺旋都是右手螺旋,这是因为组成生物体结构的氨基酸大多是L型氨基酸(糖类大多是D型),这也就导致当其组成左手螺旋时,其侧链基团的第一个C(β-C)会和羰基氧过于接近产生排斥
-
-
-
-
-
-
-
- 在个别蛋白质的个别肽段中(一种嗜热菌),其蛋白酶Asp-Asn-Gly-Gly(226-229)存在左手螺旋
-
-
-
β-折叠
特征
-
-
-
-
- 多肽链一个充分伸展的一种构象。两条或多条伸展的多肽链(或一条多肽链的若干肽段)侧向积聚,通过相邻主链上的N-H与C=O之间有规则的氢键,形成锯齿状片层结构,即β-折叠片
-
-
-
-
-
-
-
- 氨基酸的R基团在片状平面(纸面)的上方或下方交替出现
-
-
-
类别
-
-
-
-
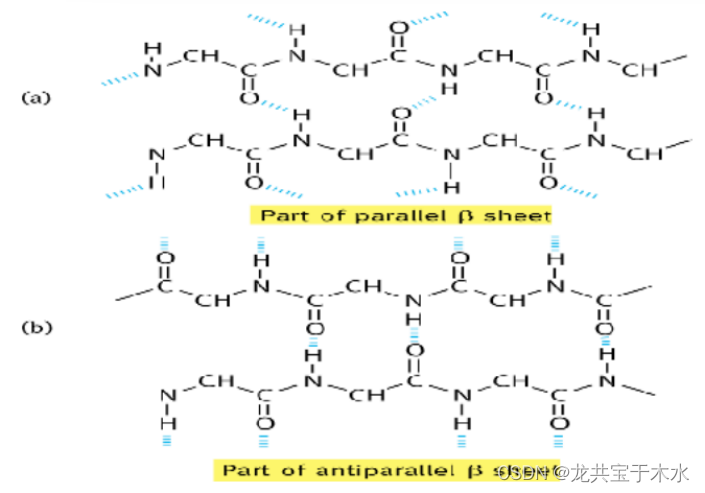
- 正向平行:两条多肽链中氨基都在一侧,羧基在另一侧
-
-
-
-
-
-
-
- 反向平行:羧基氨基扭过头相反排列
-
-
-
-
-
-
-
- 纤维状蛋白主要是反向平行
-
-
-
-
-
-
-
- 球状蛋白两种皆有
-
-
-
β-转角
很多蛋白质需要发挥功能需要进行折叠,特别是球状蛋白质,所以必须要在蛋白质的某些部位发生折叠才能形成这样的结构
特征
-
-
-
-
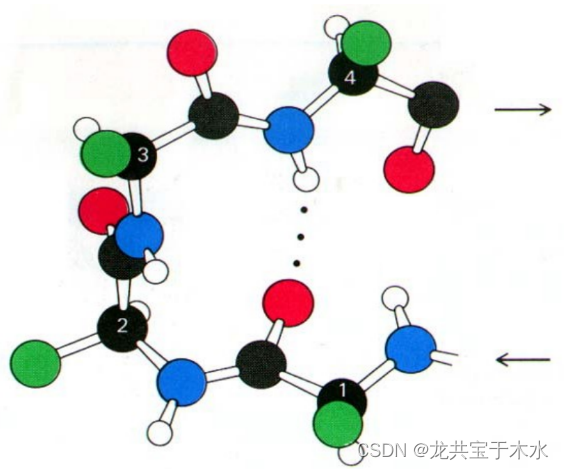
- 多肽链中氨基酸残基n的羰基上的氧与残基(n+3)的氮原子上的氢之间形成氢键,肽键回折180°
-
-
-
虽然脯氨酸和甘氨酸是α-折叠的破坏者,但它们却常常参与β-转角的组成
β-转角的特定 构象 在一定程度上取决与他的组成 氨基酸 ,某些氨基酸如 脯氨酸 和 甘氨酸 经常存在其中,由于甘氨酸缺少 侧链 (只有一个H),在β-转角中能很好的调整其他 残基 的 空间阻碍 ,因此是 立体化学 上最合适的氨基酸;而脯氨酸具有环状结构和固定的角,因此在一定程度上迫使β-转角形成,促使多肽自身回折且这些回折有助于反平行β折叠片的形成
无规则卷曲(coil)
蛋白质的一级结构决定蛋白质的二级结构,无规则卷曲这样的结构在氨基酸的排列组合的时候就已经确定了会形成
虽然无规则卷曲相比其它二级结构来说“无规则”,但是这种结构往往是一个蛋白质分子重要的活性位点与酶活性中心
某些氨基酸在二级结构中出现的概率

纤维状蛋白
定义
是动物体的基本支架和外保护成分,呈纤维状,与多肽有规律的二级结构有关
α-角蛋白
定义
-
-
-
-
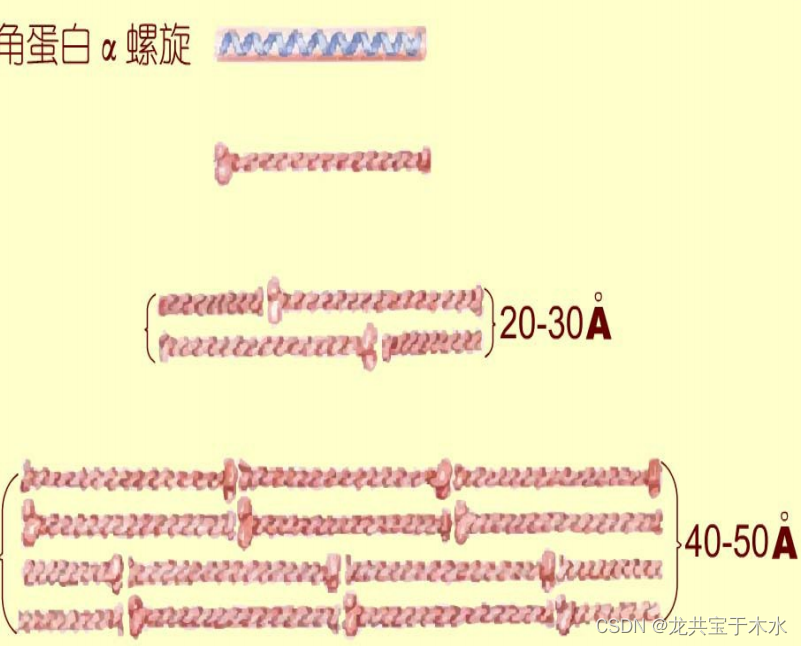
- α-角蛋白是头发的主要成分,α-角蛋白是一种沿长轴方向的α-螺旋,靠近氨基端和羧基端加粗
-
-
-
-
-
-
-
- 一对这样的螺旋彼此缠绕(左手螺旋)形成双链的螺旋化螺旋(coiled coils)
-
-
-
-
-
-
-
- 螺旋化螺旋继而结合成高度有序的结构
-
-
-
头发丝
-
-
-
-
- 两条α-螺旋形成的超螺旋,两条超螺旋形成原纤丝,四条原纤丝形成原初纤维,四条原初纤维形成中间丝(头发纤维的基本结构)
-
-
-

-
-
-
-
- 头发在加热的时候是可以伸长的:形成头发的蛋白质一级结构中半胱氨酸含量高(但没有指甲、牛角那样非常高),其侧链R基团比较大,其不适合形成β折叠,导致其容易α-螺旋结构可以被拉开
-
-
-
-
-
-
-
- 而指甲、牛角硬的软蛋白中蛋白质含有非常多的半胱氨酸,导致链间容易形成二硫键,进一步稳定了其空间结构
-
-
-
-
-
-
-
- 永久烫发的原理就和肽链间的二硫键的拆开还原形成有关(烫头发之前抹的药剂及二硫键的还原剂)
-
-
-
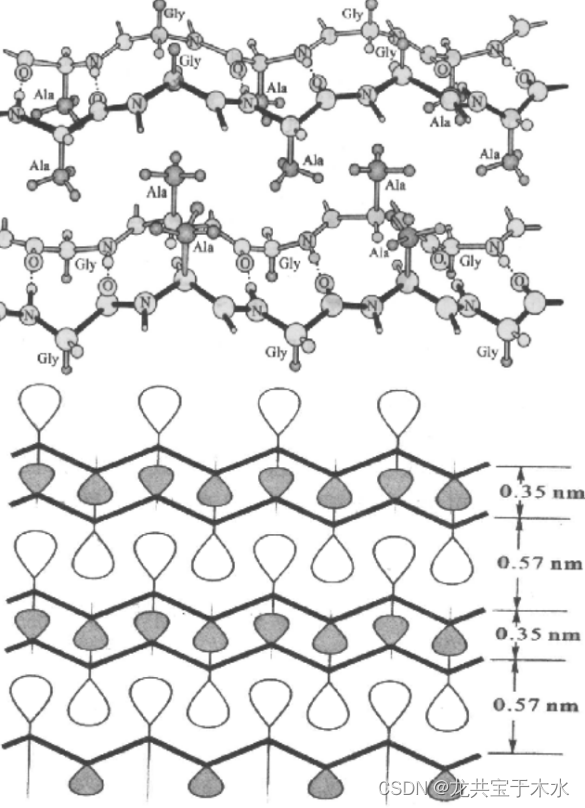
β-角蛋白
蝉丝、蜘蛛丝就属于此类(丝心蛋白fibroin)
特征
-
-
-
-
- 堆积的反向平行的β-折叠片的三维结构
-
-
-
-
-
-
-
- 富含Gly甘氨酸、Ala丙氨酸、Ser丝氨酸(小的氨基酸)
-
-
-
-
-
-
-
- 交替层中的Ala(或Ser)残基和Gly残基侧链(H原子)的连锁
-
-
-
-
-
-
-
- 结构就是甘氨酸在一侧,丝氨酸和丙氨酸在另一侧,像这样侧向紧密排列,形成齿状结构,使其抗牵拉作用很强(本身又是β-折叠片这样充分伸展的结构导致其更不能在拉长)
-
-
-
胶原蛋白(collagen)
定义
-
-
-
-
- 是脊椎和无脊椎动物体内含量最丰富的蛋白,能使肌腱、软骨、牙、皮和血管等结缔组织具有机械强度
-
-
-
三股胶原蛋白分子(左手螺旋)交替缠绕形成原胶原(右手螺旋);原胶原分子交替排列形成胶原原纤维,每一股原胶原分子在末端均会膨大,在电镜下出现明暗相间的条带
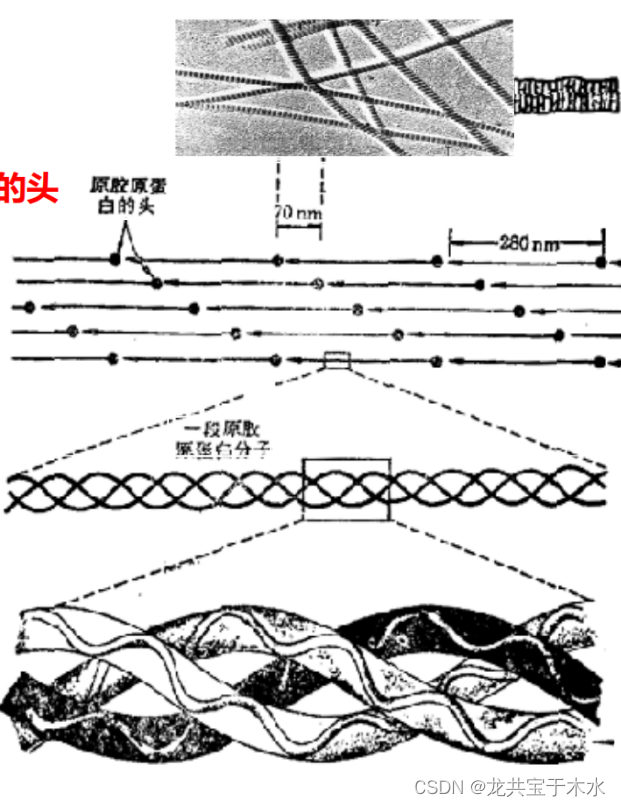
胶原原纤维可通过分子内或分子间的共价交联进一步加强它的强度
-
-
-
-
- 分子内:通过氨基端的赖氨酸残基,其ε-氨基在赖氨醯胺氧化酶作用下,被氧化成一种醛的衍生物,再聚合称为醛醇
-
-
-
-
-
-
-
- 分子间:通过分子间羧基端或者氨基端的赖氨酸以及吡啶啉交联形成锁链素(交联体)得到进一步稳定和增强
-
-
-
-
-
-
-
- 对于人体而言,随着年龄的增加,胶原纤维的交联度会逐渐上升,造成肌腱失去弹性、骨头变脆、眼镜晶体变模糊
-
-
-
超二级结构和结构域
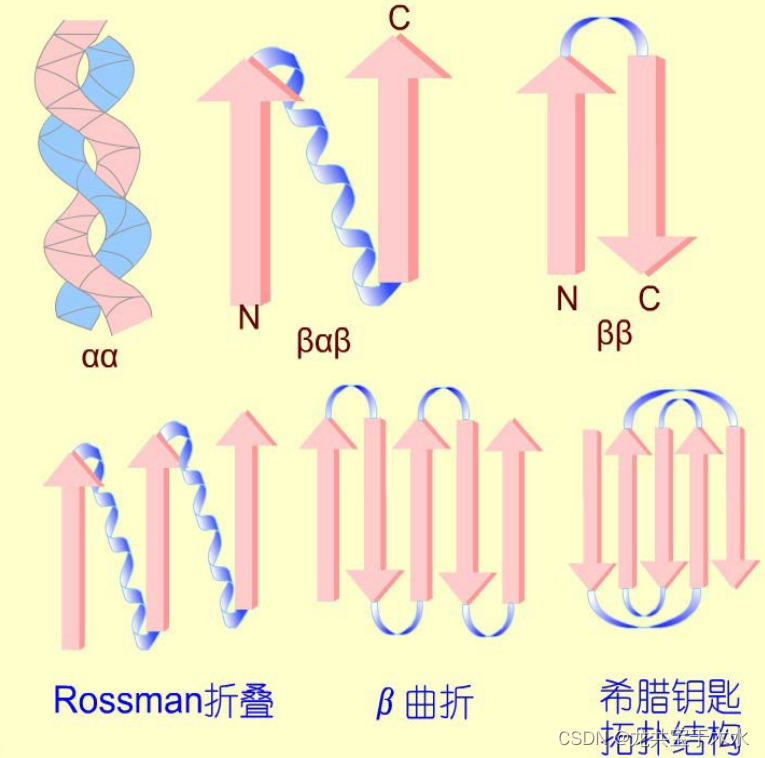
超二级结构(super-secondary structure)
概念
-
-
-
-
- 在蛋白质分子中,特别是球状蛋白质中,由若干相邻的二级结构单元(即α-螺旋、β-折叠片和β-转角等)彼此相互作用组合在一起,形成有规则、在空间上能辨认的二级结构组合体,充当三级结构的构建单元,称超二级结构或模体(motif)或折叠花样(folding motif)
-
-
-
类型
-
-
-
-
- α α
-
-
-
-
-
-
-
- β α β
-
-
-
-
-
-
-
- β β
-
-
-
结构域(structure domain)
概念
-
-
-
-
- 结构域的概念有三种不同而又相互联系的涵义:即独立的结构单位、独立的功能单位和独立的折叠单位
-
-
-
结构域运动
-
-
-
-
- 结构域本身都是紧密装配的,结构域之间通过松散的肽键形成牢固而又柔韧的连接,为域间较大幅度的相对运动提供了可能,这种结构调整与其整体功能的行使密切相关(特别是酶的活性改变)
-
-
-
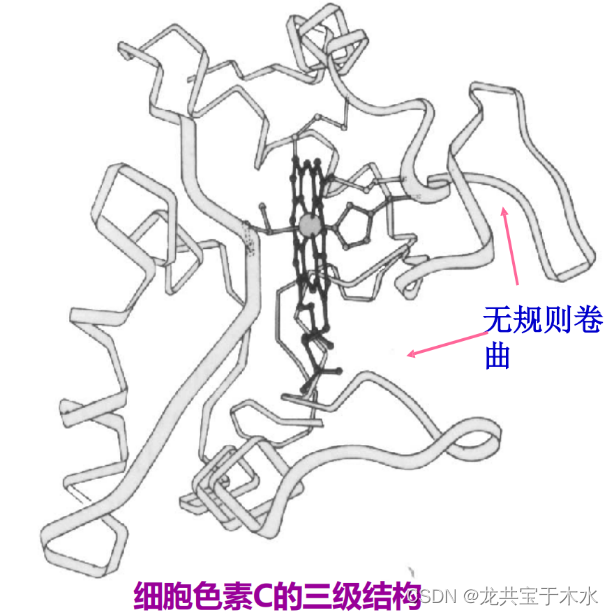
球状蛋白质与三级结构
三级结构(tertiary structure)
是指球状蛋白质的多肽链在二级结构的基础上,通过侧链基团的相互作用进一步卷曲折叠,借助次级键维系使α-螺旋、β-折叠、β-转角和无规则卷曲等二级结构相互配置而形成特定的构象。
三级结构的形成使肽链中所有的原子都达到空间上的重新排布
特征
-
-
-
-
- 含多种二级结构单元
-
-
-
-
-
-
-
- 有明显的折叠层次
-
-
-
-
-
-
-
- 是紧密的球状或椭圆状实体
-
-
-
-
-
-
-
- 分子表面如果存在空穴就可能是酶的活性部位
-
-
-
-
-
-
-
- 疏水侧链埋藏在分子内部,亲水侧链暴露在分子表面(结果就是蛋白质能够溶于水)
-
-
-
全α-结构(反平行α-螺旋)蛋白质
三级结构域中包含的二级结构以α-螺旋为主,一般情况下α-螺旋反向平行
最常见的即由四股α-螺旋通过稍微左手螺旋的方式以一种环的结构相连,每一股排列的方向,氨基羧基方向相反
蚯蚓的血红蛋白;细菌中分离的铁蛋白;细胞色素b1

α、β结构(平行或混合型β-折叠片)蛋白质
单饶平行β桶
-
-
-
-
- 催化两种磷酸丙糖相互异构反应的酶:磷酸丙糖异构酶即是这样的结构
-
-
-
-
-
-
-
- 一般是由8个平行的β-折叠片排列形成一个闭合的桶装的结构,桶存在明显的层次,里面是β-折叠片,外面是α-螺旋进行保护
-
-
-
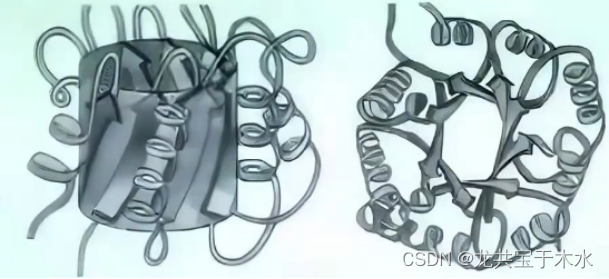
双饶平行β片
-
-
-
-
- 一般是由4个或者到9个折叠成一种开放状结构,从顶上看类似马鞍的结构
-
-
-
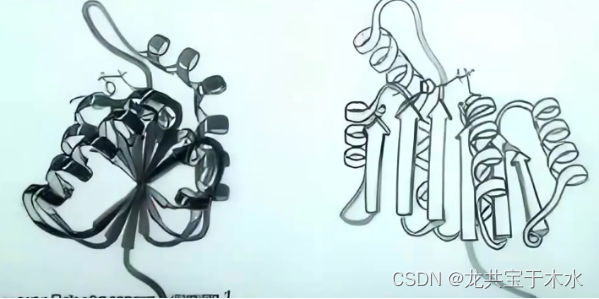
全β-结构(反平行β-折叠片)蛋白质
-
-
-
-
- 把疏水残基安排在一侧,亲水残基安排在另一侧
-
-
-
-
-
-
-
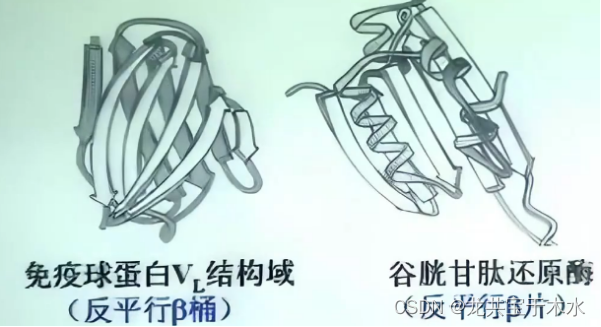
- 反平行β桶-免疫球蛋白VL结构域(轻链结构域)
-
-
-
-
-
-
-
- 反平行β片-谷胱甘肽还原酶
-
-
-
富含金属或二硫键(小而不规则)蛋白质
-
-
-
-
- 胰岛素
-
-
-
-
-
-
-
- 二节荠蛋白
-
-
-
-
-
-
-
- 高氧还势铁蛋白
-
-
-
-
-
-
-
- 铁氧还蛋白
-
-
-
全β-结构(反平行β-折叠片)蛋白质
反平行β-折叠片把疏水残基安排在一侧,亲水残基安排在另一侧
富含金属或二硫键(小的不规则)蛋白质
此类蛋白质三维结构没有明显规律性,二级结构的种类也很少,但其本身通过富含的金属离子络合或者二硫键来稳定其三维结构

蛋白质的折叠
定义
多肽链的折叠形成了二级结构和三级结构的各种花式
氨基酸序列(一级结构)决定了蛋白质的三维(高级)结构
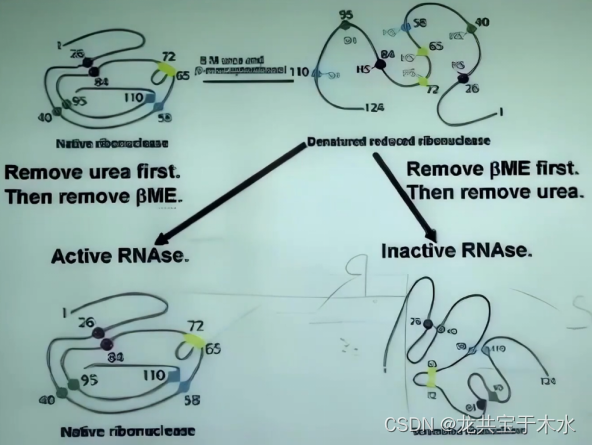
核糖核酸酶的变性与复性试验
1961年(20世纪60年代),克里斯蒂安·B·安芬森(Christian B. Anfinsen)做了一个有趣的实验,即牛胰核糖核酸酶的变性复性实验,发现去折叠蛋白或者变性蛋白质在实验条件下可以重新折叠到原来的结构,根据其研究表明,蛋白质在加热或者放置于某些化学环境中而变性,三维结构解体,而当环境恢复原本的状态时,蛋白质可于不到一秒的时间折叠至原来的立体结构,并且无论实验多少次,蛋白质都仅此一种立体结构
先除去尿素再除去还原剂,酶能够恢复原来的活性状态,顺序反过来就不行(尿素能破坏氢键,导致蛋白质分子结构松弛,使蛋白质变性,还原剂可以保护蛋白质结构不受尿素影响)
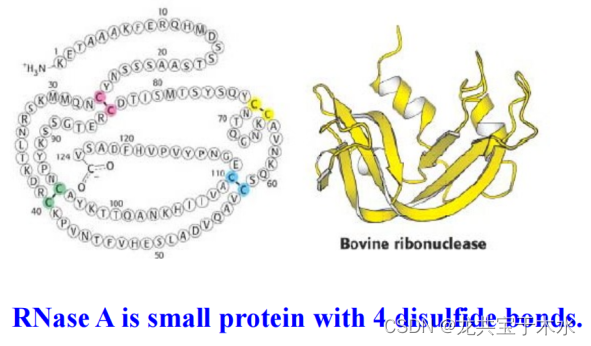
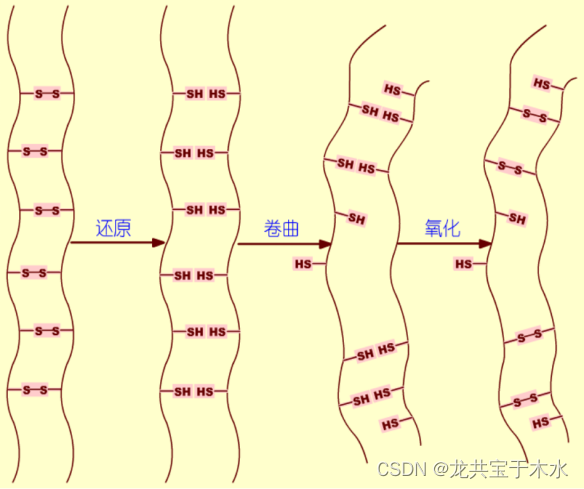
二硫键在稳定蛋白质构象中的作用
二硫键对多肽的正确折叠并不是必要的,但对稳定正确折叠具有重要作用
蛋白质的二硫键位点的多样性造成了可以有多种二硫键组合,但只有一种二硫键结构形成的蛋白质才具有生物活性
胰蛋白酶抑制剂(BPTI)是一种小分子蛋白质,图示为二硫键的存在对BPTI稳定性的影响
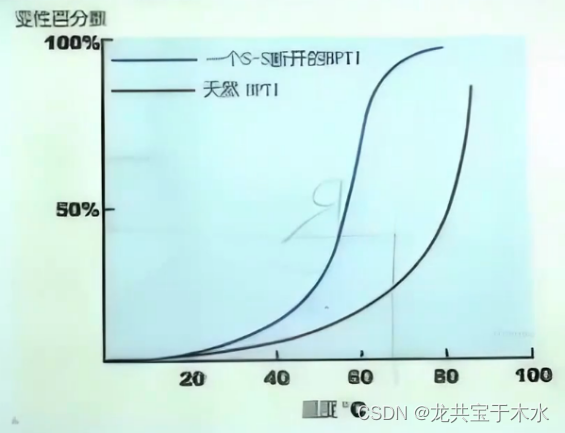
工业生产中,通过人为的方式改变酶蛋白质的一些氨基酸残基,加入一些能够形成二硫键的半胱氨酸残基,提高其对温度的稳定性
蛋白质折叠的热力学与动力学

蛋白质在水溶液中的折叠是热力学有利的过程:即蛋白质的折叠方向朝自由能最小的方向
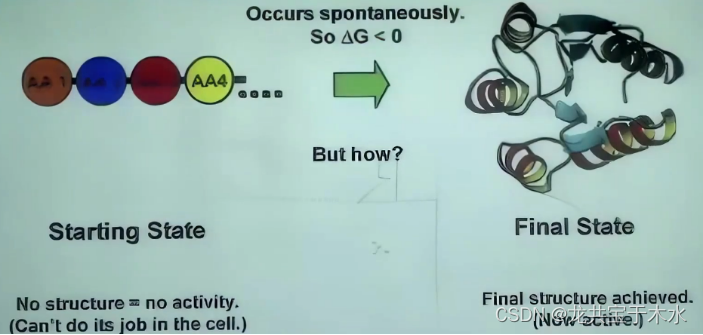
对于为什么蛋白质这种从无序状态到有序状态还能是热力学有力的过程,是因为:蛋白质溶液系统的熵增是疏水相互作用的主要动力
-
-
-
-
- 当疏水化合物残基进入水中时,它周围的水分子将排列成刚性的有序结构(笼型结构,这种排列比固体水-冰中水分子的排列还要有规律)
-
-
-
-
-
-
-
- 水分子这种由无序向有序的变化是对蛋白质的折叠的又一个不利的过程,但蛋白质维持稳定的疏水基团有避开水分,自身聚集在一起的倾向,这种相反的过程(疏水相互作用)排列有序的水分子笼型结构将被破坏。水分子被排入自由水中
-
-
-
-
-
-
-
- 最终的结果——水的混乱度增加即熵增加。所以,疏水相互作用是熵驱动的自发过程,导致蛋白质能够在水溶液中折叠成相对稳定的三维结构
-
-
-
Levinthal Paradox

-
-
-
-
- 如果用穷举法的思维理解蛋白质的折叠——蛋白质可以尝试所有可能的构象并最终找到正确的折叠:即最低能量的折叠。那么对于长度为例如 100 个氨基酸的蛋白质,每个氨基酸3种状态(二面角),它将会有3的100次方即10的47次方种构象,假设这条多肽链每秒能尝试10的12次方构象,它仍然需要10的35次方秒(宇宙大爆炸到今天有120亿年才相当于10的20次方秒)
-
-
-
-
-
-
-
- 实际上蛋白质的折叠过程只需要1~2秒,蛋白质在折叠过程中并不是一个一个去尝试可能的构象,而是沿着折叠通路,这种蛋白质折叠的中间状态称为“熔融的球状”
-
-
-
体内蛋白质的折叠有异构酶和分子伴侣参与
新生蛋白质中正确的二硫键配对受蛋白质二硫键异构酶(protein disulfide isomerase, PDI)的催化
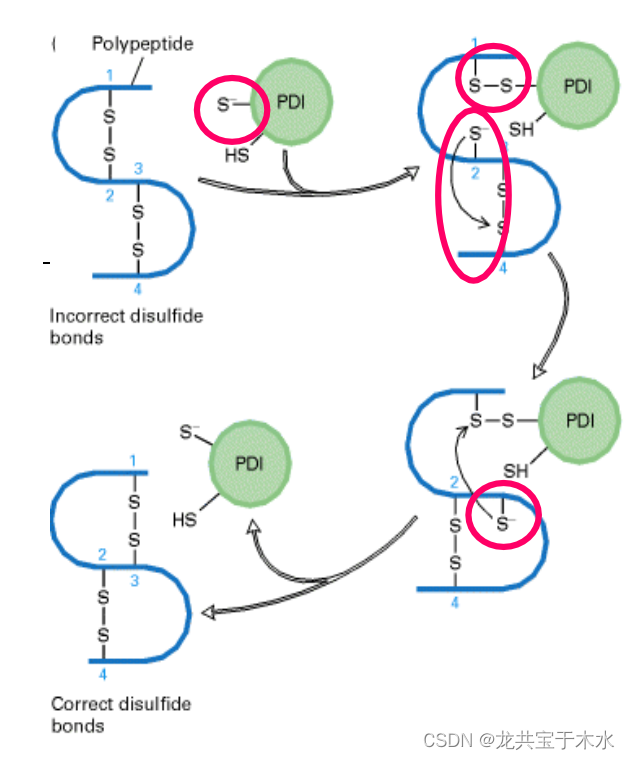
蛋白质中的肽键几乎总是反式的,但X-pro是例外。肽基脯氨酰异构酶(peptidyl prolyl isomerase)可催化顺-反异构化
-
-
-
-
- 脯氨酸是亚氨基酸,这导致它在形成肽链的时候常常以顺式连接,顺式是不稳定的结构,而这个酶可以催化,使其连接变成反式
-
-
-
分子伴侣(molecular chaperone)是一类与部分折叠的或不正确折叠的多肽结合,以简化正确折叠途径并提供折叠微环境的蛋白质
-
-
-
-
- Hsp70(Heat shock protein当某一细胞突然经过高温处理后,此类蛋白质会大量地在细胞中生成)家族:与伸展多肽疏水区结合,防止肽链不恰当聚集,保护已热变性和正在合成的蛋白,或关闭那些必须以伸展状态跨膜运输的多肽的折叠(有些蛋白质需要以线性-依次沿着细胞器-的方式运输)
-
-
-
-
-
-
-
- 陪伴蛋白(chaperonin, or Hsp60,一组分子量大概在60k道尔顿的蛋白质,最初是经过热处理后发现,后来发现不经过热处理细胞也合成此类蛋白质):是许多不能自发折叠蛋白折叠所必需的
-
-
-
-
-
-
-
- 需要注意的是,在体外蛋白质的折叠可以是自发的,但在体内还是需要酶、分子伴侣或者是特殊的蛋白质来辅助折叠
-
-
-
-
-
-
-
- 在医药生产中,以酵母菌或者大肠杆菌作为生物反应器生产蛋白质,将蛋白质从细胞内庞杂的蛋白质中抽提出来后,需要用分子伴侣来帮助其复性
-
-
-
蛋白质的四级结构
引言
四级结构(quaternary structure)指由相同或不同的称作亚基(subunit)的亚单位按照一定排布方式缔合而成的蛋白质结构,亚基之间常常以非共价键(血红蛋白、肌动蛋白)连接,有时也会以二硫键(抗体、糖蛋白)等共价键进行连接
四级结构的蛋白质中每个球状蛋白称为亚基,有时也称为单体(monomer)
一个亚基组成的蛋白质如核糖核酸酶称单体蛋白质
由两个或两个以上亚基组成的蛋白质统称为寡聚蛋白质或多聚蛋白质或多亚基蛋白质
-
-
-
-
- 同多聚蛋白质(homomultimeric)
-
-
-
-
-
-
-
- 杂多聚蛋白质(heteromultimeric)
-
-
-
-
-
-
-
- 对称的寡聚蛋白质可视为由两个或多个不对称的相同结构成分组成,称为原聚体或原体(protomer)
-
-
-
维持四级结构稳定的作用力:疏水键、离子键、氢键、范德华力
亚基本身都具有球状三级结构,一般只包含一条多肽链,也有的由二条或二条以上由二硫键连接的肽键组成
四级缔合在结构和功能上的优越性
增强结构稳定性
-
-
-
-
- 四级缔合之后其表面积和体积的比值比亚基分开后要小得多,而一般情况下,蛋白质表面和水的相互作用是不利于蛋白质稳定的,所以降低此比值有利于蛋白质稳定性
-
-
-
提高遗传经济性和效率
-
-
-
-
- 减少基因链的长度,原来需要多个基因片段的蛋白质如果其亚基是一种即可只需要一段基因链即可编码
-
-
-
使催化剂基团汇集在一起
具有协同性和别构效应
-
-
-
-
- 调节酶的活性和催化效应,很多酶也是多亚基的结构
-
-
-


















 5155
5155

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言











