







 本文详细介绍了线粒体与叶绿体的功能,包括线粒体的形态、动态特征、超微结构、氧化磷酸化过程,以及叶绿体的结构、光合作用的原初反应和电子传递。同时探讨了线粒体与叶绿体的半自主性、DNA、蛋白质转运和起源。
本文详细介绍了线粒体与叶绿体的功能,包括线粒体的形态、动态特征、超微结构、氧化磷酸化过程,以及叶绿体的结构、光合作用的原初反应和电子传递。同时探讨了线粒体与叶绿体的半自主性、DNA、蛋白质转运和起源。
目录
ATP合酶的结合变构机制(binding change mechanism)
Proton leak mediated by the uncoupling proteins
电子传递链(electron transport chain)呼吸链(respiratory chain)
The mitochondrial electron-transport chain
氧化磷酸化(oxidative phosphorylation)
Light-dependent reactions of photosynthesis at the thylakoid membrane
1.电子传递(photosynthetic electron transfer chain)
第一节 线粒体与氧化磷酸化
一、线粒体的基本形态及动态特征
高度动态的细胞器,包括运动导致的位置和分布的变化以及融合和分裂介导的形态、体积与数目的变化等。
(一)线粒体的形态、分布和数目
- 呈颗粒或短线状;
- 分布与细胞内能量需求密切相关;
- 线粒体的数目呈动态变化并接受调控;与细胞类型相关,随着细胞分化而变化。
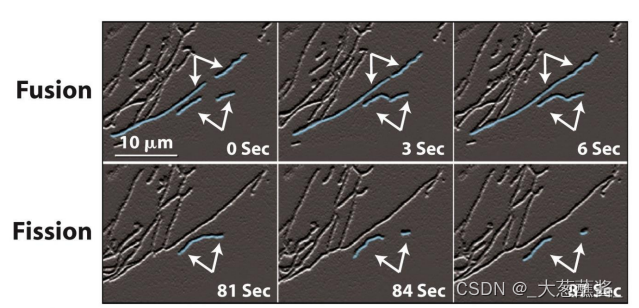
(二)线粒体的融合和分裂
线粒体融合和分裂时线粒体形态调控的基本方式,也是线粒体数目调控的基础。

1.线粒体融合和分裂的分子基础
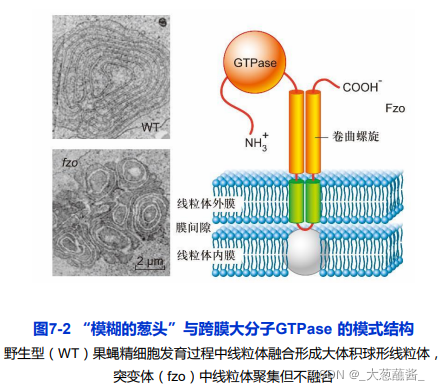
依赖于特定的基因和蛋白质的参与和调控;
果蝇精细胞线粒体融合基因Fzo(fuzzy onion)编码一个跨膜大分子GTPase,定位在线粒体外膜上
哺乳动物线粒体融合素(mitofusin), 编码线粒体融合素的Fzo 同源基因被称作 Mfn • 线粒体分裂必需的dynamin 类蛋白同样是 一类大分子GTPase
2.线粒体融合与分裂的结构动力基础
线粒体的融合和分裂装置(fusion and division apparatus)是参与线粒体融合或分裂的所有蛋白质在细胞内组装而成的功能单位。
线粒体分裂环(mitochondrial division ring):环绕线粒体的蛋白质缢缩结构
线粒体的融合与分裂是一个动态过程,需要特定的动力学机制保证。介导线粒体融合与分裂的分子力学机制被称为线粒体的融合与分裂装置(fusion and division apparatus)。
是指参与线粒体融合和分裂的所有蛋白质在细胞内组装而成的功能单位。
相比之下线粒体融合装置较为简单而分裂装置较为复杂。线粒体分裂环(FtsZ环)的形成参与线粒体的分裂。
2. 线粒体融合与分裂的结构动力基础
线粒体分裂环(mitochondrial division ring)
分裂的三个阶段:早期;中期;后期。
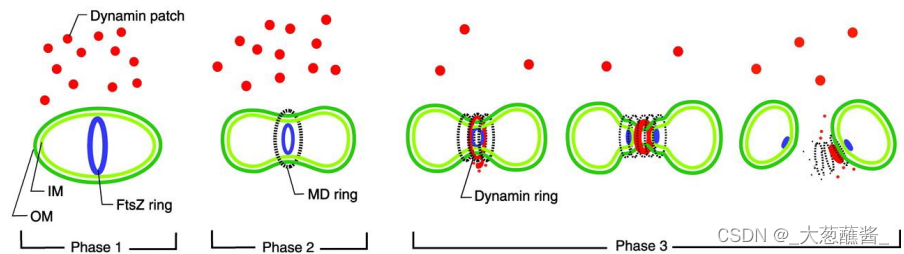
3. 线粒体融合与分裂的生物学意义
线粒体基质中含有高水平的氧化自由基,容易导致DNA损伤。线粒体可以通过不断融合和分裂来平衡这种损伤,以保证部分受损的线粒体可以正常工作。
线粒体的融合与分裂显然也是线粒体大小、数目及分布调控的基础。
植物体细胞中的线粒体数目远大于细胞中线粒体DNA的拷贝数。植物细胞中的线粒体遗传信息在线粒体之间呈现出显著的不均等分布,需要依赖频繁的线粒体融合和分裂实现遗传信息的共享。
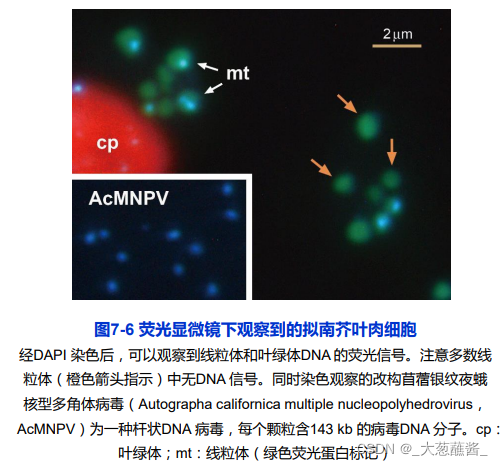
融合与分裂:会帮助细胞调整代谢状态?
细胞处于营养缺失状态时,线粒体延伸自身长度,使其产能效率提高以维持ATP产生并保证细胞活力。
细胞营养过剩时,线粒体分裂变得活跃,导致ATP产量下降,以避免能量浪费。

二、线粒体的超微结构
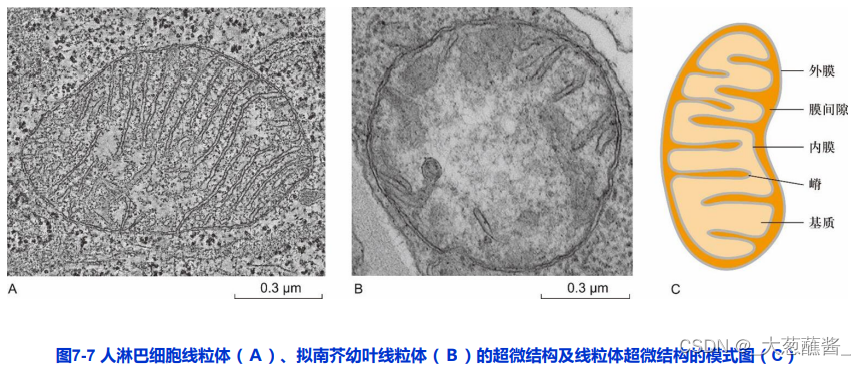
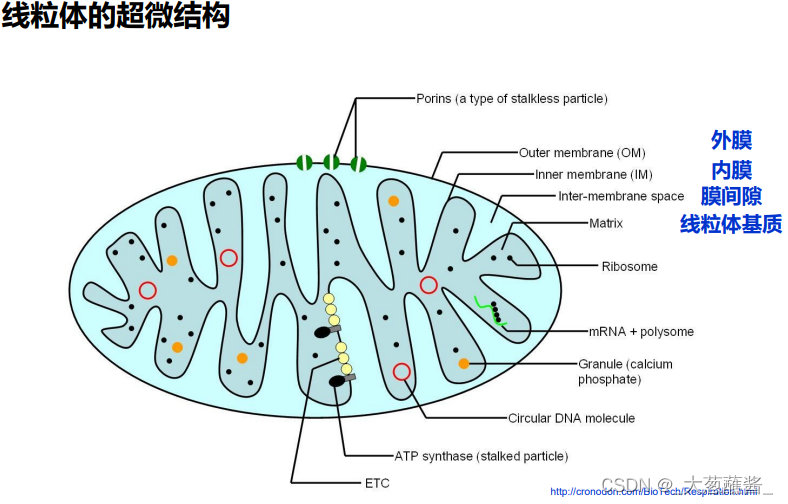
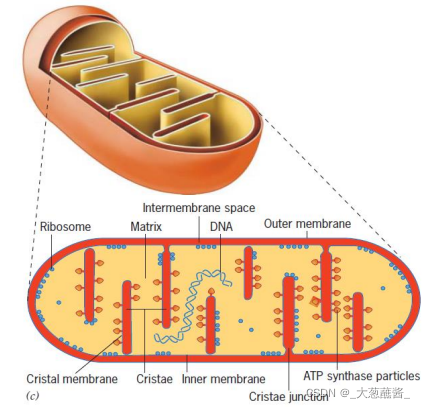
外膜(outer membrane)
- 通透性高;孔蛋白;标志酶是单胺氧化酶;
内膜(inner membrane)
- 不透性;嵴;ATP合成酶;标志酶是细胞色素氧化酶;氧化磷酸化的关键场所;
膜间隙(inter membrane space)
- 标志酶是腺苷酸激酶;
线粒体基质(matrix)
- 催化线粒体重要生化反应;标志酶是苹果酸脱氢酶;含DNA、RNA、核糖体及转录、翻译必需重要分子。
ATP 合酶(ATP synthase)
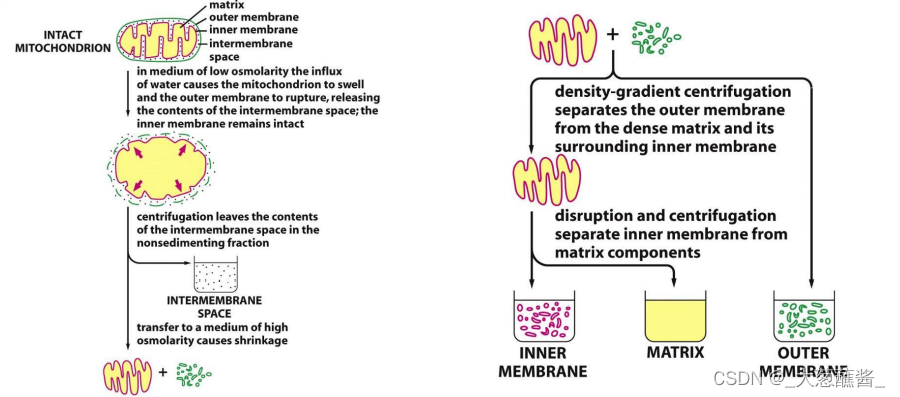
线粒体组分分离的方法
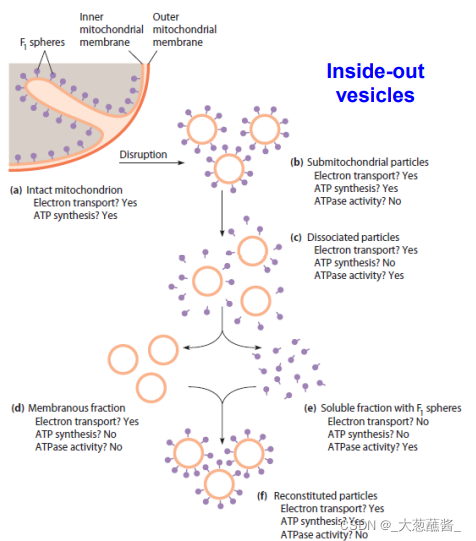
亚线粒体小泡
- 用超声波破碎线粒体,形成颗粒朝外的小膜泡,称亚线粒体小泡,具有正常的电子传递和磷酸化功能。
- 用细胞色素c的抗体能够一直完整的线粒体氧化磷酸化,但不能抑制亚线粒体颗粒的氧化磷酸化。
三、氧化磷酸化
细胞通过氧化磷酸化进行能量转换,其内膜上的ATP合酶、电子传递及膜本身的理化性质为氧化磷酸化提供了必要的保障。
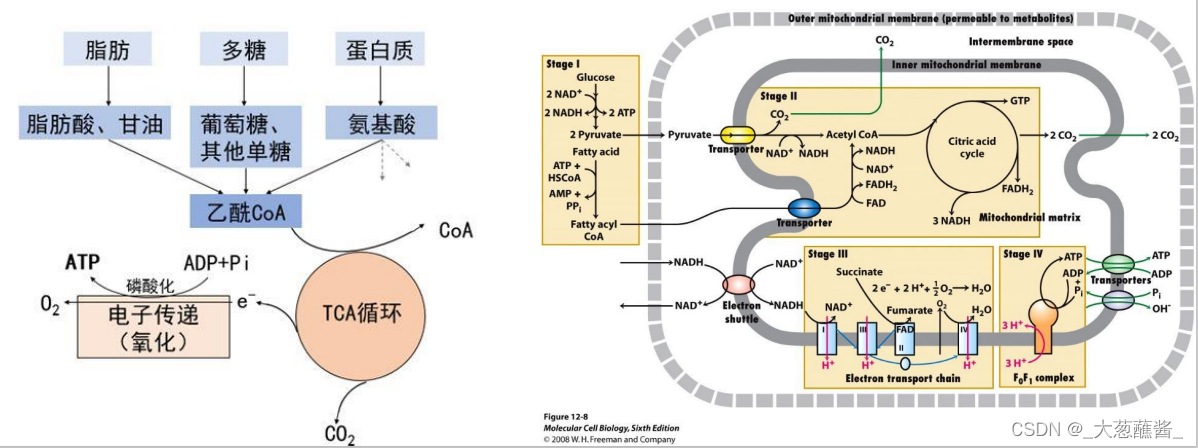
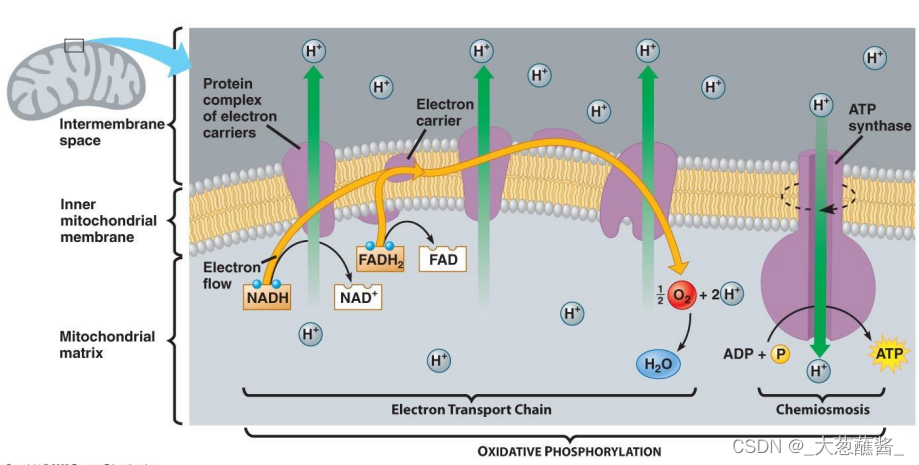
• 内膜的不透性;电子传递链;质子驱动力;ATP合酶
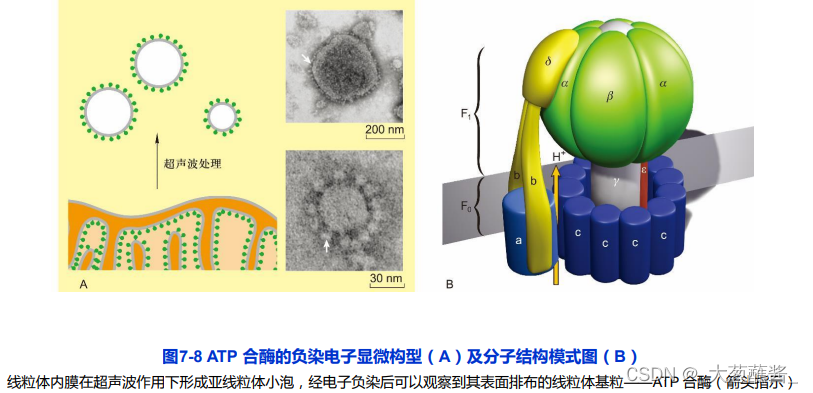
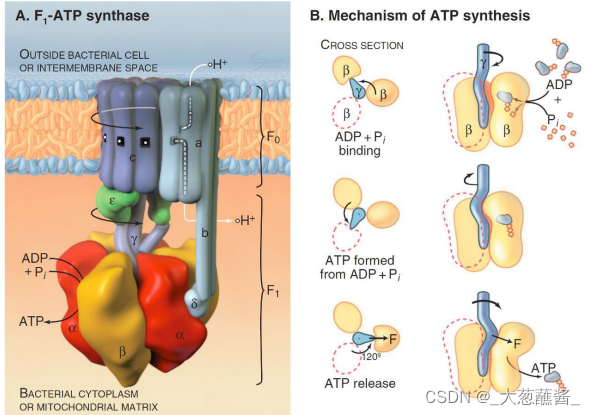
(一)ATP合酶
由球形的头部和基部组成,头部朝向线粒体基质;
头部称为偶联因子1(couping factor 1,F1),组分为 α3 β3γεδ。
基部称为偶联因子0(coupling factor 0,F0)。F0 是一个疏水性蛋白复合体,嵌合于线粒体内膜,由a、 b、c 3 种亚基按照ab2c10~12 的比例组成跨内膜的质子通道。
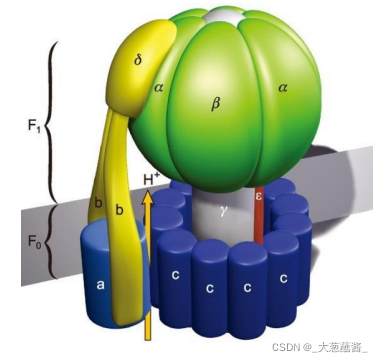
- 转子(rotor):γ与ε亚基组成;
- 定子(stator):a亚基、b亚基及F1的δ亚基共同组成。
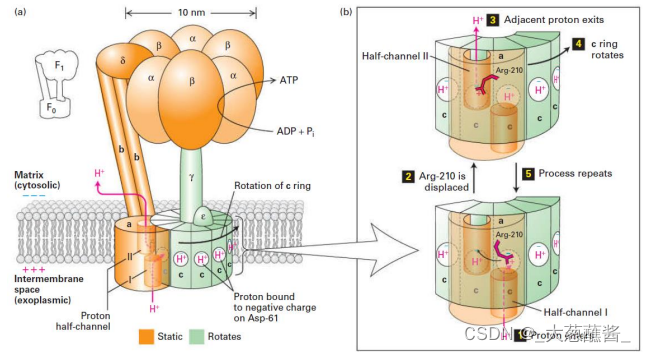
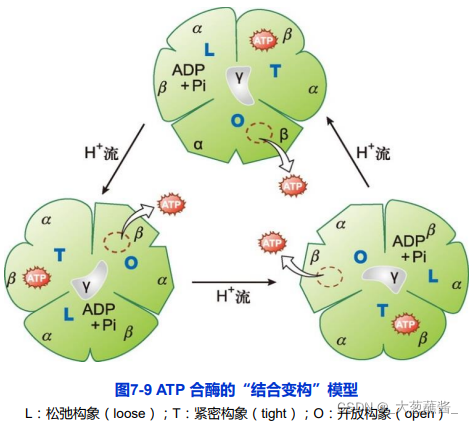
ATP合酶的结合变构机制(binding change mechanism)
• L:松弛构象(loose) • T:紧密构象(tight) • O:开放构象(open)
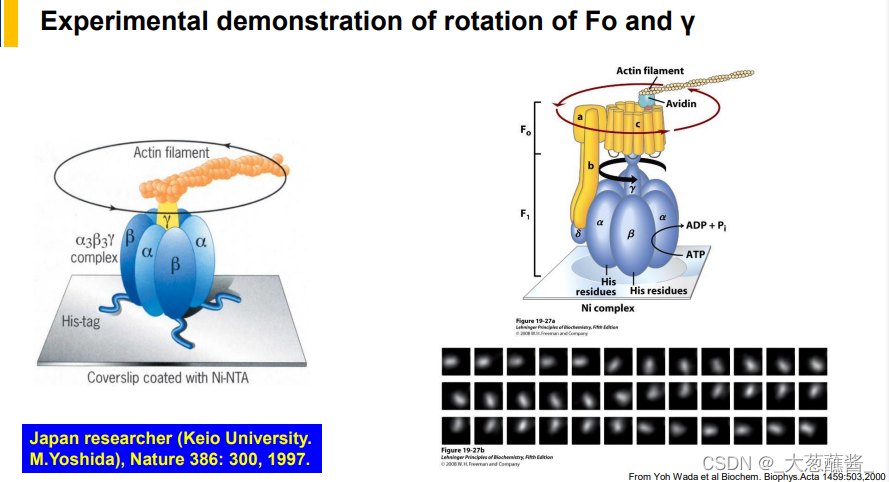
(二)质子驱动力(proton motive force)
- 膜间隙与基质间的质子浓度梯度的形成与保持是线粒体合成ATP的基本前体;
- 线粒体内膜上的电子传递为膜间隙和基质之间的质子梯度提供了保证;
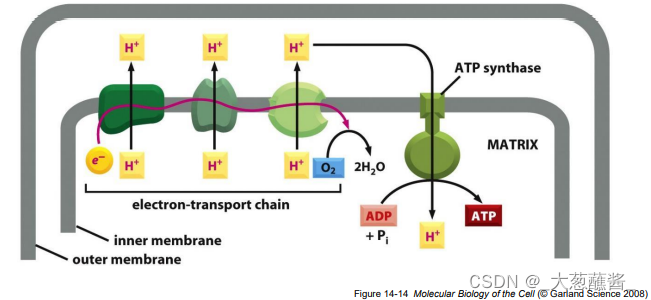
• H+ 跨膜电位差 • 质子浓度梯度(pH 差)
线粒体产能(ATP)原理
- 电子传递过程中,内膜上的电子传递复合物将基质中的质子转运至膜间隙,形成ATP合酶工作所需的质子梯度;
- 能量转换实质上是将质子驱动力转换为ATP分子中的高能磷酸键; <
 最低0.47元/天 解锁文章
最低0.47元/天 解锁文章


















 7861
7861

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









