






Genome-Wide Identification of BAHD Superfamily and Functional Characterization of Bornyl Acetyltransferases Involved in the Bornyl Acetate Biosynthesis in Wurfbainia villosa
阳春砂(Wurfbainia villosa)中BAHD超家族的全基因组鉴定及参与龙脑乙酸生物合成的龙脑乙酰转移酶功能特性研究
植物中的酰基转移酶:正是BAHD大展身手的好时机-文献精读65_bahd酰基转移酶-CSDN博客
Littorine生物合成糖基转移酶和酰基转移酶-文献精读39_trv2载体通用引物-CSDN博客
惕佫酰假托品合酶的发现-文献精读28_discovering a mitochondrion-localized bahd acyltra-CSDN博客

摘要
乙酸龙脑酯 (Bornyl acetate, BA) 是一种天然芳香单萜酯,具有广泛的药理和生物活性。龙脑乙酰转移酶 (Borneol acetyltransferase, BAT) 能催化龙脑和乙酰辅酶A (acetyl-CoA) 合成乙酸龙脑酯,是一种酒精乙酰转移酶,属于BAHD超家族酰基转移酶。然而,目前尚未对负责乙酸龙脑酯生物合成的BAT进行功能表征。阳春砂(Wurfbainia villosa,学名同义词:Amomum villosum)的种子富含乙酸龙脑酯。本研究通过使用PF02458(转移酶)和PF07247(AAT酶)作为隐马尔可夫模型 (HMM) 筛选BAHD基因,从阳春砂的基因组中鉴定了64个BAHD家族成员。这64个WvBAHD基因分布于14条染色体和9个未锚定的contig上,并聚类为6个分支,其中3个带有PF07247的WvBAHD基因形成了一个独立的新分支:分支VI。
我们选择了属于分支I-a、I-b和VI的12个候选基因进行克隆和体外功能表征,其中8个基因被鉴定为BAT基因,能够催化至少一种类型的龙脑乙酰化生成乙酸龙脑酯。所有8个WvBAT基因都可以利用(-)-龙脑作为底物,但只有5个WvBAT基因能够催化(+)-龙脑,而(+)-龙脑是阳春砂种子中的内源性龙脑底物;WvBAT3和WvBAT4对(+)-龙脑的催化效率显著高于其他基因。WvBAT基因的时间和空间表达模式显示,WvBAT3和WvBAT4是种子特异性表达基因,其表达水平与乙酸龙脑酯的积累呈正相关,这表明WvBAT3和WvBAT4可能是阳春砂种子中乙酸龙脑酯合成的两个关键BAT基因。
这是首次报道负责乙酸龙脑酯生物合成最后一步的BAT基因,这将为进一步研究乙酸龙脑酯的生物合成及其在其他植物或异源宿主中的代谢工程提供重要基础。
引言
天然产物一直是药理活性分子的重要来源,具有疗效显著、副作用少等优势。乙酸龙脑酯(Bornyl acetate, BA) 因其丰富的药理作用而备受关注。研究表明,BA具有较强的脂氧合酶抑制作用,从而能够减轻炎症/过敏反应、肿瘤及阿尔茨海默病,并减少由5-氟尿嘧啶引起的肠道黏膜炎症(Zhang et al., 2017;Cutillas et al., 2018)。此外,BA被证实对人类MCF-7、HT-29和H-1299细胞具有潜在的增殖抑制作用(Sajjadi et al., 2015)。更有研究报道,BA具有镇痛、抗氧化、美白、抗癌、抗肿瘤、抗流产及抗焦虑等作用,同时在挥发油中与其他芳香化合物协同作用,表现出抗菌、杀虫及麻醉等效果(Wang et al., 2011;Asghari et al., 2012;Ohtsubo et al., 2015;Perestrelo et al., 2016;Zhang et al., 2017;Ao et al., 2019)。
然而,BA仅分布于少数植物科中,包括姜科(Zingiberaceae)、松科(Pinaceae)、柏科(Cupressaceae)、芸香科(Rutaceae)、伞形科(Umbelliferae)、唇形科(Lamiaceae)和漆树科(Anacardiaceae)。其中,阳春砂(Wurfbainia villosa,学名同义词:Amomum villosum) 的种子富含BA,其种子精油中的BA含量为10.53%(占总单萜含量的50.35%),显著高于其他植物,如月桂树(Laurus nobilis,果实,4.40%)、香叶木姜子(Illicium pachyphyllum,果实,4.01%)和缬草(Valeriana jatamansi,根,0.6–1.5%)(Liu et al., 2012;Raina and Negi, 2015;Wang et al., 2018;Fidan et al., 2019;Chen et al., 2020b)。因此,阳春砂是解析BA生物合成途径的重要研究材料。阳春砂属于姜科植物,其芳香干果被称为砂仁(中药名:砂仁),是一种著名的传统中药,在中国尤其是南方地区被广泛用于治疗消化系统疾病及日常烹饪中。BA是阳春砂的主要药效成分和质量指标,其含量决定了阳春砂的质量(中国中医药管理委员会,1999;中国药典委员会,2015)。除了高水平的BA,阳春砂还含有其他低含量或痕量的萜类乙酸酯,如乙酸异龙脑酯(IBA)、乙酸橙花叔醇酯和乙酸檀香醇酯等,这些成分是阳春砂特有的香气和风味成分,并与其药理活性密切相关(Zhao et al., 2021b;补充表1)。
在植物中,所有萜类骨架均通过甲羟戊酸(MVA)途径和2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)途径合成,这些途径已被广泛研究(Vranová et al., 2013)。龙脑二磷酸合酶(BPPS) 是BA生物合成下游途径中的第一个关键酶,其催化单萜前体牻牛儿基二磷酸(GPP) 生成龙脑二磷酸,后者是龙脑的前体。BPPS已从少数植物中克隆和表征,如阳春砂的WvBPPS和肉桂的CbBPPS(Wang et al., 2018;Ma et al., 2021)。二磷酸在脱磷酸作用下转化为龙脑,而龙脑则通过酒精乙酰转移酶的乙酰化作用生成BA。酒精乙酰转移酶(AAT, EC 2.3.1.84) 可催化萜醇、芳香醇或脂肪醇生成植物中的芳香挥发性乙酸酯(图1;Beekwilder et al., 2004;Souleyre et al., 2005)。AAT已从草莓、香蕉、苹果、桃等果实以及玫瑰和薰衣草等花卉以及传统草药如九里香(Celastrus angulatus)、罗勒(Ocimum basilicum)等植物中克隆和鉴定(补充表2;Beekwilder et al., 2004;Souleyre et al., 2005;Zhang et al., 2010;Sarker and Mahmoud, 2015;Dhar et al., 2020;Yan et al., 2020)。
BA是通过龙脑的乙酰化生成的,而龙脑有三种光学异构体,包括左旋、右旋和外消旋体(Drienovská et al., 2020;Chánique et al., 2021)。相较之下,化学合成的龙脑含有四种不同的立体异构体,即(+)-龙脑、(-)-龙脑、(+)-异龙脑和(-)-异龙脑(Khine et al., 2020)。然而,阳春砂中负责BA及其他单萜乙酸酯合成的AAT尚未被表征。此外,负责催化不同类型龙脑生成BA的龙脑乙酰转移酶(BAT)基因迄今尚未有报道。
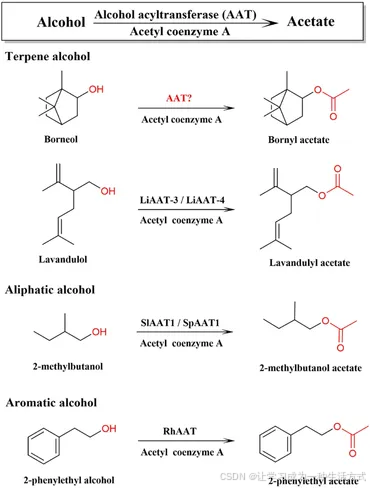
图1. 酒精乙酰转移酶 (AATs) 在乙酸酯生物合成途径中的作用
酒精乙酰转移酶(AATs)是BAHD酰基转移酶家族中的重要成员。BAHD家族以其前四个被生化表征的酶(BEAT、AHCT、HCBT和DAT)的首字母命名,并具有两个高度保守的基序:“HXXXD”和“DFGWG”(St-Pierre和De Luca, 2000)。迄今为止鉴定出的BAHD成员均为单体酶,其分子量范围为48至55 kDa(D’Auria, 2006)。AATs主要在积累大量挥发性酯的植物组织中高表达,例如腺毛、花朵(花瓣/柱头)或果实(果肉/花托),其中一些在叶或茎中表达(Beekwilder et al., 2004;Souleyre et al., 2005;Sarker和Mahmoud, 2015;Dhar et al., 2020;Yan et al., 2020)。果实中表达的A
酒精乙酰转移酶(AATs)是BAHD酰基转移酶家族的重要成员。BAHD家族的命名来源于该家族中首批被生化表征的四种酶(BEAT、AHCT、HCBT和DAT)的首字母,并且其显著特征是具有两个高度保守的基序:“HXXXD”和“DFGWG”(St-Pierre和De Luca, 2000)。迄今为止鉴定的BAHD家族成员均为单体酶,其分子量范围为48至55 kDa(D’Auria, 2006)。AATs主要在植物组织中高表达,这些组织通常积累大量挥发性酯,如腺毛、花朵(花瓣/柱头)或果实(果肉/花托),部分AATs在叶或茎中表达(Beekwilder et al., 2004;Souleyre et al., 2005;Sarker和Mahmoud, 2015;Dhar et al., 2020;Yan et al., 2020)。果实中的AATs表达通常受到果实成熟的诱导。AATs的底物范围广泛,包括醇类前体和乙酰辅酶A (acyl-CoA) 供体,一个AAT可能同时参与多种酯的合成和积累(Ma et al., 2005;Balbontín et al., 2010;Nimitkeatkai et al., 2011;Cumplido-Laso et al., 2012;Souleyre et al., 2014)。虽然AAT在体外可以利用更多种类的醇类底物,但酯化化合物的生物合成主要取决于植物内源性底物的可用性。阳春砂种子可能含有多种AAT,因为种子是积累BA及其他单萜乙酸酯的主要器官。
在本研究中,作为解析阳春砂BA生物合成途径的一部分,我们从阳春砂基因组数据库中鉴定了64个BAHD家族成员,并选择了12个候选WvAAT基因进行克隆和功能表征。通过生化实验,确定其中8个基因能够编码BAT,催化至少一种类型的龙脑转化为BA,其中两个基因进一步被证明是阳春砂种子中BA合成的关键BAT基因。
材料与方法
植物材料
阳春砂采自中国广东省阳春市。从健康植株中采集叶片、根茎、花和果实,并冷冻保存于−80°C。对发育阶段不同的果实(分别为开花后30天、45天、60天、75天和90天)进行采集,并分离果皮和种子。
全基因组鉴定BAHD超家族
为了鉴定阳春砂中的BAHD候选基因,我们使用蛋白质基础逻辑比对工具(BLASTP)(E值阈值为1e-5),将拟南芥 (Arabidopsis thaliana) 和苹果 (Malus domestica) 的BAHD蛋白序列(Liu et al., 2020)与阳春砂基因组数据(未发表)进行比对。这些氨基酸序列来自Phytozome数据库。随后,根据Pfam数据库中BAHD结构域 (PF02458) 的隐马尔可夫模型 (HMM),我们使用Hmmsearch (3.2.1)(Potter et al., 2018)进一步鉴定阳春砂中的候选WvBAHD基因。我们使用Web CD-Search Tool和Web SMART工具确认通过上述两种方法鉴定的候选WvBAHD基因的结构域,并检查其是否具有“HXXXD”和“DFGWG”基序。此外,利用PF07247(酒精乙酰转移酶,AATase)补充筛选BAHD基因。通过MCSanX软件鉴定WvBAHD基因的共线性区域,并使用Circos生成圆形图结果(Wang et al., 2012)。氨基酸序列的比对和最大似然(ML)进化树(自举值为1,000)通过MUSCLE和IQ-TREE(TBtools中的工具)获得,进化树分支的分类参考Liu et al. (2020)。
参与乙酸龙脑酯生物合成的候选WvAAT基因的筛选
候选WvAAT基因通过其聚类的分支、未发表的RNA-seq数据中转录本每百万表达值(TPM)以及不同组织和果实发育阶段中与上游基因WvBPPS的关联表达情况直接筛选得到。为了预测WvAAT的底物偏好,我们使用ML方法对已报道的AAT进行多序列比对和系统发育分析(工具为MEGA X,Kumar et al., 2018)。系统发育分析中所用的其他植物AAT信息见补充表2。通过Jalview软件对候选基因的核苷酸和氨基酸序列进行多序列比较分析(Waterhouse et al., 2009)。保守基序和活性基序的预测使用在线工具MEME(Bailey和Elkan, 1994)。WvAAT候选基因的蛋白质结构、信号肽和转运肽通过在线平台TMHMM 2.0、SignalP和ChloroP 1.1 Server进行预测和分析(Emanuelsson et al., 1999;Krogh et al., 2001;Petersen et al., 2011)。根据序列比对和进化树的结果,进化树通过在线工具iTOL进行美化(Letunic和Bork, 2021)。
扩增全长WvAAT候选基因
总RNA和互补DNA(cDNA)的提取方法参考此前研究(Wang et al., 2018)。AAT候选基因的编码区通过使用适当引物(补充表3)和Primer STAR Max DNA聚合酶(Takara,中国)从阳春砂的cDNA中扩增获得。PCR反应条件如下:98°C预变性1分钟;98°C 10秒,50–60°C退火15秒,72°C延伸15秒,共30个循环;最后在72°C延伸5分钟。纯化后的PCR产物连接到pLB(Tiangen,中国)或007 vs.(TSINGKE,中国)克隆载体中,随后转化至大肠杆菌DH5α中并进行测序。
WvAAT重组蛋白的原核表达与纯化
候选WvAAT基因的全长开放阅读框(ORF)通过In-Fusion Cloning Kit(Takara,中国)连接到pET32a(+)表达载体中,随后转化至大肠杆菌Rosetta(DE3)*中。引物信息见补充表3。细胞在37°C培养至OD600达到0.4–0.6,然后加入终浓度为10 μM的*异丙基-β-D-硫代半乳糖苷(IPTG) 于16°C诱导16–20小时,培养基为Luria–Bertani (LB) 培养基,并添加50 μg/μL羧苄青霉素和25 μg/μL氯霉素。重组蛋白通过NI-NTA树脂(Qiagen,德国)纯化,并使用PD-10脱盐柱(GE Healthcare)透析去盐。
WvAAT候选基因的酶活性检测及产物分析
体外酶促反应在总体积250–350 μL的反应体系中进行,包含:20 mM磷酸氢钠缓冲液(pH 9.0),10%甘油,20–80 μg纯化蛋白,0.2 mM乙酰辅酶A(acetyl-CoA),以及0.02 mM萜醇类底物((+)-龙脑、(-)-龙脑、异龙脑、牻牛儿醇、橙花醇、α-萜品醇、(-)-薄荷醇、芳樟醇和橙花叔醇)。根据AAT酶促反应的研究(Croteau and Hooper, 1978;Sarker and Mahmoud, 2015)及初步实验结果,WvAAT的最优pH和温度分别为9.0和32°C。所有反应混合物在32°C孵育6小时,然后加入250 μL正己烷覆盖。反应混合物以12,000 rpm离心5分钟,分离相后用Agilent 7890B气相色谱仪(GC)和5977A惰性质谱检测器(MSD)(Agilent,美国)分析正己烷提取物。载气为氦气(1 mL/min),分离柱为HP-5MS柱(30 m × 250 μm × 0.25 μm膜厚)。GC的初始温度设为35°C保持2分钟,然后以每分钟12°C升温至300°C,并在300°C保持5分钟。对于手性化合物,使用CycloSil-B柱(30 m × 0.25 mm id, 0.25 μm膜厚),初始温度为50°C保持2分钟,然后以5°C/min升温至180°C,再以10°C/min升至230°C并保持2分钟。代谢物通过NIST14/Wiley275质谱库鉴定,同时使用标准品进一步确认。每种WvAAT基因的实验均重复三次。
挥发性萜类的提取与分析
取约0.1 g发育不同时期的种子,用液氮研磨后加入1 mL正己烷提取,用超声波清洗器处理30分钟,并在40°C孵育1小时。样品以10,000 rpm离心15分钟,取上清液转移至新的2 mL离心管中。取1 mL正己烷提取物转移至1 mL小瓶中,用于GC-MS分析。分析条件与上述相同,使用CycloSil-B柱分离。初始温度设为35°C保持2分钟,然后以5°C/min升温至200°C,再以10°C/min升至230°C并保持2分钟。最后温度保持在240°C 5分钟。通过NIST14/Wiley275质谱库鉴定萜类化合物,主要萜类乙酸酯及其前体萜类醇用标准品进一步确认。每种器官的实验均进行三次生物学重复和三次技术重复。
WvBAT基因的实时荧光定量PCR分析
基于阳春砂的转录组数据与酯类数据的相关性分析,从中选择了8个WvBAT基因用于实时荧光定量PCR(qRT-PCR)。所有WvBAT基因的引物手动设计(荧光定量引物信息见补充表3)。qPCR使用2xTSINGKE® Master qRT-PCR Mix-SYBR (+ UDG, TSINGKE, 中国) 在CFX96实时PCR检测系统(Bio-Rad,美国)中进行。WvBAT的转录水平以内参基因TUA为标准,使用2–ΔΔCt法计算。所有实验均进行三次生物学重复和三次技术重复。
WvBAT基因启动子的分析
对启动子区域进行初步元件分析,提取WvBAT基因编码区上游2,000 bp的非编码区域(通过TBtools),并在PlantCare在线平台上分析是否存在与种子特异性表达相关的顺式作用元件(Lescot et al., 2002)。功能基因启动子的顺式作用元件分类参考Abdullah等(2018年)的研究。
结果
阳春砂 WvBAHD 基因家族与 WvAAT 候选基因的全基因组鉴定
通过使用 BAHD HMM 配置文件 (PF02458, 转移酶) 对阳春砂 (W. villosa) 的基因组数据库进行筛选,共鉴定出 61 个具有完整开放阅读框 (ORF) 和两个保守基序的 BAHD 基因。此外,利用 PF07247 (AATase) 作为 HMM 工具,筛选出另外 3 个基因(WvBAHD3、WvBAHD52 和 WvBAHD53)。AATase (PF07247) 属于蛋白家族 CL0149,这一家族名为 CoA-acyltrans,其特征同样为 HXXXD 基序,与转移酶家族 (PF02458) 一致。AATase 家族包含一些来自细菌和后生动物的 AAT,能够催化乙酰辅酶A (acetyl-CoA) 对异戊醇 (isoamyl alcohol) 进行酯化,这一功能与植物中的 AAT 类似(Minetoki et al., 1993;Zhu et al., 2015;Lin et al., 2016;Reyes-Sánchez et al., 2019)。因此,我们推测,通过 PF07247 (AATase) 筛选出的这 3 个基因应属于 BAHD 超家族。
总计鉴定出 64 个 WvBAHD 基因,它们被归为 BAHD 超家族的酰基转移酶成员(补充表 4),分布在 14 条染色体和 9 个未锚定的 contig 上(图 2A)。基因组共线性分析显示,发生了 15 次片段复制事件 (占 23.4%) 和 4 次串联重复事件 (占 6.3%),表明片段复制事件在 WvBAHD 基因家族扩展中起到了重要作用(图 2B)。跨膜功能预测结果表明,大多数 WvBAHD 基因定位于细胞质中(补充表 5)。
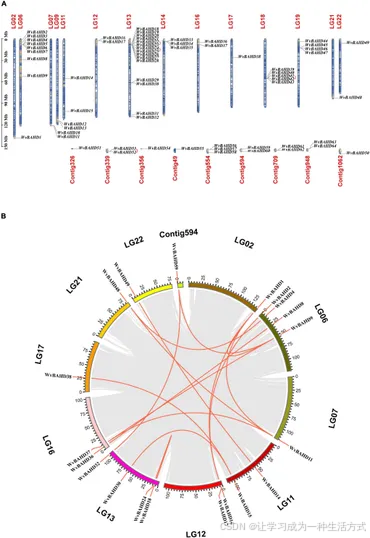
图 2. WvBAHD 基因家族的基因定位和共线性分析
(A) WvBAHD 基因在染色体上的定位和分布分析。串联重复的基因通过红线连接。 (B) 在 Circos 图中进行 WvBAHD 基因的染色体复制分析。红线表示同线性基因对。
为了确定 WvBAHD 基因家族的分类,我们利用来自阳春砂 (W. villosa)、拟南芥 (A. thaliana)、苹果 (M. domestica) 的 235 个 BAHD 基因和 14 个其他植物中鉴定的 AAT 基因的氨基酸序列构建了系统发育树。64 个预测的 WvBAHD 基因被分为 5 个分支:I-a(22 个基因)、I-b(15 个基因)、II-a(9 个基因)、II-b(14 个基因)、III-a(1 个基因)和 VI(3 个基因)(图 3A)。属于分支 I-a 的基因在拟南芥和苹果中参与芳香族醇和萜类醇的修饰,因此我们推测阳春砂中聚类到分支 I-a 的 WvBAHD 基因可能编码具有类似功能的蛋白。然而,鉴定的 AAT 也聚类到分支 I-b 和 IV,这与之前的研究不一致,后者报告分支 I-b 的成员与木质素单体中间体的生物合成相关(Hoffmann et al., 2003;Petrik et al., 2014)。这可能是由于系统发育树算法的差异,以及这些酶尚未被全面鉴定。
此外,三个 AATase-WvBAHD 基因独立聚类为一个新分支,并命名为分支 VI。因此,我们推测 WvAAT 候选基因的范围从分支 I-a 扩展到分支 I-b 和分支 VI。为了验证上述聚类与保守基序之间的相关性,我们利用 MEME 检测了阳春砂 WvBAHD 基因的八个保守基序。Motif1 和 Motif2 分别对应保守基序 HXXXD 和 DFGWG(图 3B 和补充图 1、2)。相同分支中保守基序的类型和分布相似性进一步支持了系统发育树的分类。
在分支 VI 的成员中,HXXXD 基序中的组氨酸残基被苏氨酸或丝氨酸取代,同时 DFGWG 基序中的四个残基也发生了取代,这导致分支 VI 的成员与其他 BAHD 和已知 AAT 成员具有更远的进化关系(图 3A 和图 4)。
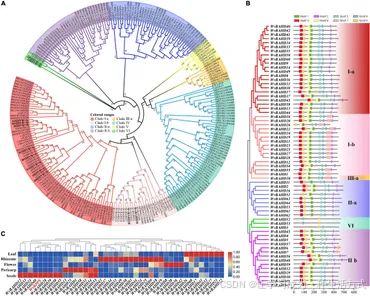
图3. (A) 阳春砂 (Wurfbainia villosa)、拟南芥 (Arabidopsis thaliana)、苹果 (Malus domestica) 的 BAHD 基因和 14 个已鉴定的 AAT 基因的系统发育分析。系统发育树通过最大似然法构建,重复 1,000 次,聚类分支分类依据 Liu 等人(2020)的研究。WvAAT 候选基因以黑色粗体字体标注。 (B) WvBAHD 基因的保守基序分析。保守基序的氨基酸序列图例见补充图 1。 (C) 分支 I 中 WvBAHDs 和 WvBPPS 的转录组差异表达分析。果皮和种子样本来源于开花后 60 天 (60-DAF) 的果实。
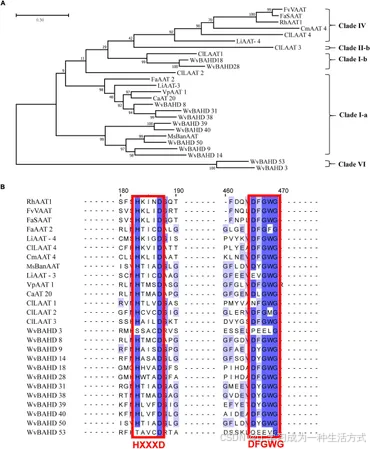
图4. (A) 12 个 WvAAT 候选基因和 14 个其他植物中已鉴定的 AAT 基因的系统发育分析。系统发育树通过最大似然法构建,重复 1,000 次,聚类分支分类依据 Liu 等人(2020)的研究。 (B) WvAAT 基因和已鉴定 AAT 蛋白中 HXXXD 和 DFGWG 基序的序列比对。序列相似性阈值为 30% 或以上,保守基序用红色高亮标注。补充表 2 包含每个 AAT 序列的 GenBank 登录号。
我们将分支 I 和 VI 的基因表达谱与龙脑生物合成的关键基因 WvBPPS 的表达进行了比较,发现 4 个基因(WvBAHD14、WvBAHD40、WvBAHD39、WvBAHD50;TPM >10)在 60-DAF 种子中与 WvBPPS 共表达,表明这些基因可能是参与龙脑乙酰化的关键基因(图 3C)。进一步分析显示,分支 I-a 中的 WvBAHD8、WvBAHD38 和 WvBAHD31 与已报道的 AAT(如 CaAT20、LiAAT3、VpAAT1 和 FaAAT2)具有较高的相似性(>45%);WvBAHD9、WvBAHD14、WvBAHD39、WvBAHD40 和 WvBAHD50 与 MsBanAAT 聚为一个紧密分支;在分支 I-b 中,WvBAHD18 和 WvBAHD28 与柠檬 (Citrus limon) 的 AAT 基因 ClLAAT1 的相似性约为 40%(图 4)。这些已报道的 AAT 基因在体外对牻牛儿醇和橙花醇显示出酶促亲和力(Aharoni et al., 2000;补充表 2),因此,我们推测这些 WvBAHD 基因可能具有与这些 AAT 类似的酶活性。
最终,从分支 I 中选择了 10 个基因(WvBAHD8、WvBAHD9、WvBAHD14、WvBAHD18、WvBAHD28、WvBAHD31、WvBAHD38、WvBAHD39、WvBAHD40 和 WvBAHD50),以及从分支 VI 中选择了 2 个基因(WvBAHD53 和 WvBAHD3),这些基因均在种子中表达(补充表 6),作为 WvAAT 候选基因用于克隆和功能表征。
8 个 WvAAT 基因被鉴定为龙脑乙酰转移酶 (BAT)
通过从 E. coli 表达菌株中提取和纯化重组蛋白(补充图 3)进行体外酶促反应实验。实验结果表明,8 个 WvAAT 重组蛋白(WvBAHD8、WvBAHD14、WvBAHD28、WvBAHD39、WvBAHD40、WvBAHD50、WvBAHD3 和 WvBAHD53)能够催化牻牛儿醇和橙花醇这两种易被催化的一元醇,分别生成牻牛儿乙酸酯和橙花乙酸酯。尽管可溶性蛋白含量较低,但这些结果验证了这 8 个候选基因具有 AAT 酶活性,并且这种方法在 AAT 活性表征中是有效的(Shalit et al., 2003;Sarker 和 Mahmoud, 2015;补充图 4)。其余 4 个候选基因在酶促反应中未检测到牻牛儿乙酸酯或橙花乙酸酯,也不能催化龙脑类底物。
为进一步确定上述 8 个 WvAAT 重组蛋白的 BAT 活性,使用龙脑类底物(包括 (-)-龙脑、(+)-龙脑和异龙脑)分析催化产物。结果表明,这些 WvAAT 均能催化 (-)-龙脑生成 (-)-乙酸龙脑酯,但只有 5 个 WvAAT(WvBAHD8、WvBAHD14、WvBAHD39、WvBAHD40 和 WvBAHD3)能够催化 (+)-龙脑生成 (+)-乙酸龙脑酯(图 5A、B)。由此得出结论,这 5 个 WvAAT 能够同时催化 (-)-龙脑和 (+)-龙脑,且对不同光学异构体的龙脑没有严格的底物特异性,而 WvBAHD28、WvBAHD50 和 WvBAHD53 仅能催化 (-)-龙脑。此外,这 5 个能够催化 (-)-龙脑和 (+)-龙脑的 WvAAT 中,WvBAHD8、WvBAHD39、WvBAHD40 和 WvBAHD3 还能够催化异龙脑生成异龙脑乙酸酯 (IBA)(图 5C)。
除了龙脑类底物外,还使用了更多不同类型的底物进行酶促反应实验。结果显示,这些 WvAAT 无法催化三元醇(如 (-)-薄荷醇、芳樟醇或橙花叔醇)生成相应的乙酸酯产物,这与 LiAAT3 和 LiAAT4 的研究结果一致(Sarker 和 Mahmoud, 2015)。由于这些 WvAAT 均能将至少一种类型的龙脑转化为乙酸龙脑酯,证明它们具有 BAT 活性,因此分别将其命名为 WvBAT1–8(表 1 和补充表 7)。
为比较不同光学异构体龙脑底物的催化活性,对 5 个能够催化 (-)-龙脑和 (+)-龙脑的 WvBAT 进行等量底物反应,并分析产物峰面积。结果表明,这些 WvBAT 对 (-)-龙脑的催化能力优于 (+)-龙脑;特别是 WvBAT3(WvBAHD39)对 (+)-龙脑表现出最佳催化效率,其次为 WvBAT4(WvBAHD40)(图 5D)。此外,WvBAT3 是唯一能够催化 α-萜品醇生成 α-萜品醇乙酸酯的酶(表 1 和补充图 5)。
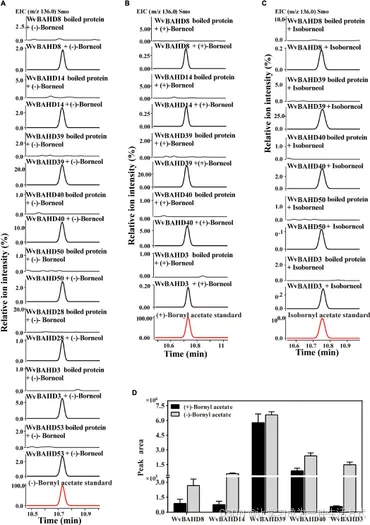
图5. WvBAHD基因的功能表征
(A–C) 使用 (−)-龙脑、(+)-龙脑和异龙脑作为底物时,各 WvBAHD 基因在体外反应中产生的产物的气相色谱–质谱 (GC-MS) 色谱图。 (D) 在相同底物浓度(酶浓度因 WvBAHD 基因不同而异)的条件下,以 (−)-龙脑、(+)-龙脑和异龙脑为底物时,各 WvBAHD 产物的 GC-MS 色谱峰面积(EIC, m/z 136.0)。
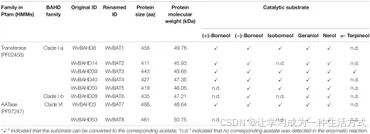
表 1. 阳春砂(*Wurfbainia villosa*)中表征的 WvBAT 基因信息。
阳春砂种子中龙脑及乙酸龙脑酯的光学构型
正如前文所述,大多数 WvBAT 在体外表现出对 (−)-龙脑更高的催化能力。(−)-龙脑和 (+)-龙脑是光学异构体,且二者均为植物中的天然代谢物。然而,目前尚未通过手性柱明确阳春砂种子中内源性龙脑及 WvBPPS 催化产物的光学构型(Wang et al., 2018)。因此,我们使用配备 CycloSil-B 柱的 GC-MS 对阳春砂种子中的龙脑及 WvBPPS 酶促反应产物的手性构型进行了鉴定。
结果显示,在阳春砂成熟种子(90-DAF)中未检测到 (−)-龙脑,但检测到大量的 (+)-龙脑和少量的异龙脑(图 6A)。这一结果在不同发育阶段的种子挥发物提取物(30-DAF 至 75-DAF)中也得到了验证,并与 WvBPPS 催化产物经磷酸酶处理后产生 (+)-龙脑的观察结果一致(图 6B)。此外,(+)-乙酸龙脑酯 (BA) 的含量随着种子发育中 (+)-龙脑的增加而升高,并在 90-DAF 达到峰值;(+)-龙脑的含量明显高于异龙脑,这与 (+)-BA 的含量显著高于异龙脑乙酸酯 (IBA) 的数据相符(图 6C)。这些结果表明,(+)-龙脑是阳春砂种子中的主要内源性龙脑类底物。
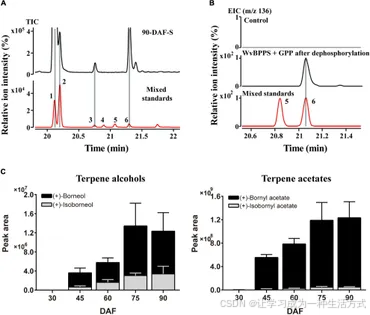
图6.(A) 阳春砂 (Wurfbainia villosa) 90-DAF 种子中挥发性萜类及其混合标准品的 GC-MS 色谱图。 (B) WvBPPS 蛋白经脱磷酸反应后生成的产物的 GC-MS 色谱图,与混合龙脑标准品的对比及煮沸蛋白处理的阴性对照。色谱峰对应化合物:1. 乙酸龙脑酯;2. 乙酸异龙脑酯;3. (+)-异龙脑;4. (−)-异龙脑;5. (−)-龙脑;6. (+)-龙脑。 (C) 阳春砂不同发育阶段种子中 (+)-龙脑、(+)-异龙脑及其相应酯类产物 (+)-乙酸龙脑酯和 (+)-乙酸异龙脑酯的 GC-MS 色谱峰面积(EIC, m/z 136.0)。
阳春砂中 WvBAT 基因表达水平与酯类积累的相关性
为了分析 WvBAT 基因的时间和空间表达模式,我们对叶片、根茎、花及 60-DAF 果实(果实分离为果皮和种子)以及种子在五个不同发育阶段的样本进行了 qRT-PCR 分析。结果显示,WvBAT 基因的相对表达水平与 RNA 测序 (RNA-seq) 数据中的 TPM(每百万转录本数)表达值基本一致,除了 WvBAT5 外;WvBAT5 实际上在根茎中的表达水平高于种子。
WvBAT3、WvBAT4 和 WvBAT6 的表达模式为种子特异性,与 WvBPPS 的表达模式相似。尽管 WvBAT6 在种子中特异表达,但其编码的酶无法催化 (+)-龙脑和异龙脑,表明它可能参与种子中其他酯类的生物合成。WvBAT1 在花中特异性表达,暗示其可能参与花中酯类的生物合成。此外,WvBAT7 在所有器官中均有表达,在根茎中的表达水平最高,表明其可能参与根茎中的乙酸龙脑酯 (BA) 合成,尽管根茎中的 BA 含量较低(Wang et al., 2018;图 7A)。

图7. WvBAT基因的表达模式
(A) WvBAT基因在不同组织中的相对表达水平。果皮和种子样本取自开花后60天(60-DAF)的果实。转录本每百万读数(TPM)值来自RNA测序(RNA-seq)数据。 (B) 能够催化(+)-龙脑的WvBAT基因在种子不同发育阶段的表达水平对比。 (C) 能够催化(+)-龙脑的WvBAT基因在45-DAF和60-DAF种子中的表达水平对比。数据表示均值±标准差(n=3)。
正如前文所述,(+)-龙脑是种子中的主要内源性底物,且乙酸龙脑酯 (BA) 在种子中显著积累,尤其在45-DAF阶段开始出现(图6C)。只有四个WvBAT基因(WvBAT2、WvBAT3、WvBAT4和WvBAT7)既能催化(+)-龙脑,又在种子中表达。为确定负责BA生物合成的主要BAT基因,我们分析了这些基因在种子不同发育阶段的表达模式,并比较了它们在45-DAF和60-DAF种子中的表达水平。结果显示,WvBAT2和WvBAT7的表达水平在45-DAF达到峰值,但它们的表达量远低于WvBAT4(图7B、C)。值得注意的是,种子特异性表达的WvBAT3和WvBAT4在30-DAF时未表达,但在45-DAF阶段开始高表达,这与WvBPPS在45-DAF阶段的最高表达水平以及种子中(+)-龙脑和(+)-BA的初始积累水平一致(Wang et al., 2018;图7B)。此外,WvBAT4在45-DAF和60-DAF种子中的表达量均为最高,而WvBAT3在60-DAF种子中的表达量也高于其他两个WvBAT基因(图7C)。考虑到WvBAT3(WvBAHD39)在(+)-龙脑上的催化效率最佳(图5D),我们推测WvBAT3和WvBAT4是负责BA合成的关键酶。此外,异龙脑乙酸酯 (IBA) 在种子中的初始积累也从45-DAF开始增加(图6C),这表明能够催化(+)-异龙脑的WvBAT3、WvBAT4和WvBAT7也参与了阳春砂种子中的IBA合成。
WvBAT基因启动子的分析
为了明确WvBAT基因在阳春砂中的潜在生物学功能,对WvBAT1–8基因启动子区域进行了顺式作用元件的分析。共鉴定出308个顺式作用元件,并将其分为三类:植物激素响应、胁迫响应和植物生长与发育(图8A、B)。在与植物激素响应相关的顺式作用调控元件中,比例最高的是与脱落酸(ABA)相关的ABRE元件,占22%,其次是MYC(21%)、TGACG基序(10%)和CGTCA基序(10%),后面三者与甲基茉莉酸 (MeJA) 相关。几乎一半的顺式作用元件与胁迫响应相关(161/304),其中最常见的四种顺式作用元件是与光响应相关的box 4(14%)、G-box(11%)和GT1基序(9%),以及与水胁迫相关的MYB(9%)(图8C)。这些结果表明,WvBAT基因可能受MeJA和ABA的诱导或抑制,并参与植物对多种非生物胁迫的响应。
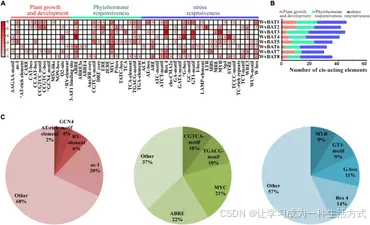
图8. (A) WvBAT 基因中顺式作用元件的数量,以红色的强度和网格中的数字表示。与种子特异性表达相关的顺式作用元件以红色星号标注。 (B) 柱状图显示与植物生长发育、植物激素响应或胁迫响应相关的顺式作用元件数量。 (C) 饼状图显示每个结构类别中不同顺式作用元件的比例。
我们之前观察到,WvBPPS 的启动子包含种子特异性表达元件 GCN4 基序,与种子中龙脑的主要积累一致(Zhao et al., 2021a)。与种子表达相关的顺式作用元件包括与胚乳元件相关的 GCN4 基序(Onodera et al., 2001)、RY 元件和参与种子特异性调控的 AT-rich 元件(Bäumlein et al., 1992;Ellerström et al., 1996),以及 G-box(Izawa et al., 1993, 1994)。正如预期,除 WvBAT5 外,这些 WvBAT 启动子均含有与种子表达相关的顺式作用元件。例如,WvBAT7 的启动子包含两个 GCN4 基序,而 WvBAT1、WvBAT2、WvBAT4、WvBAT6 和 WvBAT7 的启动子包含 G-box。
讨论
用于植物中 BAHD 酰基转移酶基因鉴定的新 Pfam 隐马尔可夫模型 PF07247
目前关于 BAHD 超家族酰基转移酶成员筛选的 HMM 研究较少。PF02458 是转移酶家族的 HMM,属于 CL0149 家族,具有特征性 HXXXD 基序。通过比较 Pfam 数据库中已报道的 BAHD 家族成员的 HMM,我们获得了 PF02458,与以往研究的结果一致(Liu et al., 2020;Kumar et al., 2021)。我们利用 PF02458 从阳春砂的基因组数据中鉴定出了 61 个 WvBAHD 基因,这些基因具有“HXXXD”和“DFGWG”基序,与 BAHD 家族的特征一致。
在本研究中,除了 PF02458,我们还使用了另一个 Pfam HMM PF07247(AATase 家族的 HMM),它同样属于 CL0149,用于鉴定阳春砂的 BAHD 家族成员。结果筛选出了三个被注释为 AATase 的 WvBAHD 基因(WvBAHD3、WvBAHD52 和 WvBAHD53)。然而,目前尚未有与 PF07247 类似的 BAHD 成员 HMM 报道。事实上,“AATase”通常用来描述来自细菌、后生动物和真菌(如酵母菌 Saccharomyces 和 Kluyveromyces)的 AAT(Zhu et al., 2015;Reyes-Sánchez et al., 2019)。为了进一步确认这三个 AATase-WvBAHD 是否属于 BAHD 酰基转移酶家族,我们基于 D’Auria 的分类系统(将 BAHD 酰基转移酶划分为六个主要组)(D’Auria, 2006;Sarker 和 Mahmoud, 2015),利用 64 个 WvBAHD 和其他物种中已鉴定的 BAHD(补充表 8),构建了另一条系统发育树。
出乎意料的是,这三个 AATase-WvBAHD 全部聚类到第 V 分支,该分支主要由参与挥发性酯类生物合成的 AAT 组成(补充图 6)。然而,它们与其他植物中已鉴定的 AAT 并未紧密聚类,这可能是由于 BAHD 酰基转移酶家族中保守基序 DFGWG 的氨基酸残基发生了取代。值得注意的是,WvBAHD3 和 WvBAHD53 确实具有 AAT 活性,且均能乙酰化龙脑、牻牛儿醇和橙花醇(表 1)。因此,本研究验证了“PF07247/AATase”对于植物中 BAHD 超家族成员筛选的可行性,从而通过新增 AAT 成员进一步丰富了 BAHD 家族。
首篇关于龙脑乙酰转移酶(BAT)参与龙脑乙酸酯(BA)最后一步生物合成的研究
龙脑乙酸酯是一种芳香单萜酯,具有多种药理和生物活性,但只有少数植物能积累显著量的 BA。BPPS 是催化龙脑前体合成的酶,也是 BA 下游生物合成途径的第一步,这种酶已从其他植物中鉴定,包括鼠尾草 (Salvia officinalis, SoBPPS)、甘草薄荷 (Lippia dulcis, LdBPPS)、薰衣草 (Lavandula angustifolia, LaBPPS) 和桂皮 (Cinnamomum burmannii, CbBPPS)(Wise et al., 1998;Despinasse et al., 2017;Hurd et al., 2017;Ma et al., 2021)。除了甘草薄荷,这些植物都含有 BA(Al-Dhubiab, 2012;Arceusz et al., 2013)。然而,催化龙脑乙酰化生成 BA 的 BAT 基因尚未被报道。在本研究中,我们基于 BAHD 家族的鉴定,从阳春砂 (Wurfbainia villosa) 中克隆并鉴定了 8 个 WvBAT 基因。这是首篇关于 BAT 负责 BA 最后一步生物合成的研究报告。
这些 WvBAT 基因能够催化至少一种类型的龙脑生成 BA,并且它们是底物广谱酶(substrate-promiscuous enzymes),类似于其他 AATs。AAT 是一种底物广谱酶,能够催化不同类型的醇和酰辅酶A(acyl-CoA)生成芳香挥发性酯类化合物(Beekwilder et al., 2004;Souleyre et al., 2005)。然而,酰基化产物的类型和含量取决于植物中内源底物的种类和含量。例如,薄荷中合成的乙酸酯类型并不取决于 AAT 的特异性,而是取决于薄荷油中萜醇的可用性(Croteau and Hooper, 1978)。对于阳春砂,其种子挥发油中含有高水平的 (+)-龙脑,较低水平的异龙脑,及微量的 α-萜品醇。因此,(+)-龙脑是阳春砂种子中 WvBAT 的主要内源底物。
两个 WvBAT 基因(WvBAT3 和 WvBAT4)能够以更高的效率催化 (+)-龙脑,并且它们在种子中的表达与 BA 的积累呈正相关,可能是协作合成阳春砂种子中 BA 的关键酶。这一推测与此前的研究一致,即植物中含有多种 AAT,这些 AAT 可能协作合成单一酯类(Cumplido-Laso et al., 2012;Sarker 和 Mahmoud, 2015)。
综上所述,基于本研究中对 WvBAT 的生化表征以及先前的文献(Wise et al., 1998;Despinasse et al., 2017;Wang et al., 2018),我们阐明了阳春砂中 BA 的下游生物合成途径(图9)。本研究将有助于更深入地理解阳春砂中 BA 和其他乙酸酯的生物合成,并为进一步研究 BA 的生物合成及在其他植物或异源宿主中的代谢工程奠定基础。
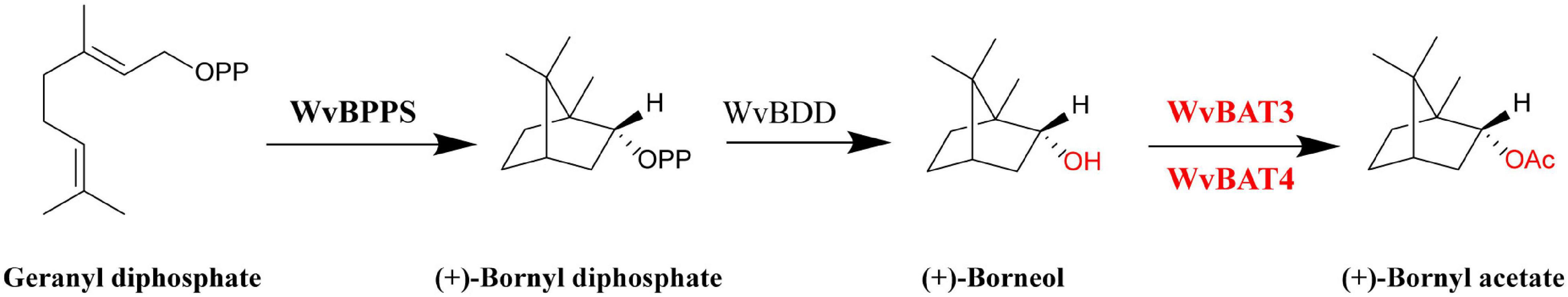
图9. 阳春砂 (*Wurfbainia villosa*) 中龙脑乙酸酯 (BA) 的生物合成途径
BPPS: 龙脑二磷酸合酶;BDD: 龙脑二磷酸磷酸酶;BAT: 龙脑酰基转移酶。标红的 WvBAT 被鉴定为参与种子中龙脑乙酸酯 (BA) 合成的关键酶。


















 2952
2952

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









