









A-to-I mRNA编辑是一种重要的基因表达调控方式,它通过将mRNA中的腺苷(A)转变为肌苷(I),从而可能改变蛋白质的编码信息。在动物中,这一过程由ADAR家族酶介导,然而在真菌中,由于缺乏ADARs的同源物,其背后的机制尚不明确。近年来,西北农林科技大学植保学院刘慧泉教授团队在一些丝状子囊菌中发现了一种有性生殖阶段特异的A-to-I mRNA编辑现象,提示可能存在一种新的、尚未被充分理解的编辑机制。

近日,刘慧泉教授团队在Nature Communications上在线发表了题为“Unveiling the A-to-I mRNA editing machinery and its regulation and evolution in fungi” 的研究论文,该研究以小麦赤霉病菌为研究材料,揭示了真菌中A-to-I mRNA编辑的酶复合体,并明确了其起源、进化和调控机制。研究结果对真菌病害防控和基因编辑工具开发具有重要意义。爱基百客为本文提供了RIP-seq技术支持。

-
【标题】Unveiling the A-to-I mRNA editing machinery and its regulation and evolution in fungi
-
【发表期刊】Nature Communications(IF:16.6)
-
【发表日期】2024年5月10日
-
【研究技术】酵母双杂交(Y2H),co-IP,DNA-seq,链特异性RNA-seq,RIP-seq(爱基百客均可提供服务)
-
【研究路线】
-
【研究内容】
1. FgTad2和FgTad3形成异源二聚体
Tad2和Tad3组成的异源二聚体是真核生物中保守的tRNA特异的腺苷脱氨酶,二者均含有CDA结构域,但Tad3中关键的催化残基谷氨酸被缬氨酸取代,失去活性(Fig.1a)。在禾谷镰刀菌有性发育1至8天的过程中,FgTAD2和FgTAD3显示出相似的表达趋势(Fig.1b)。酵母双杂交(Y2H)和免疫共沉淀(co-IP)实验都证实了FgTad2与FgTad3之间的相互作用(Fig.1c,d)。FgTad2被发现与FgTad3的C-末端区域(203-428aa)相互作用,而不与N-末端区域(1-202aa)相互作用。因此,FgTad2和FgTad3在禾谷镰刀菌中作为异源二聚体进行腺苷脱氨作用。
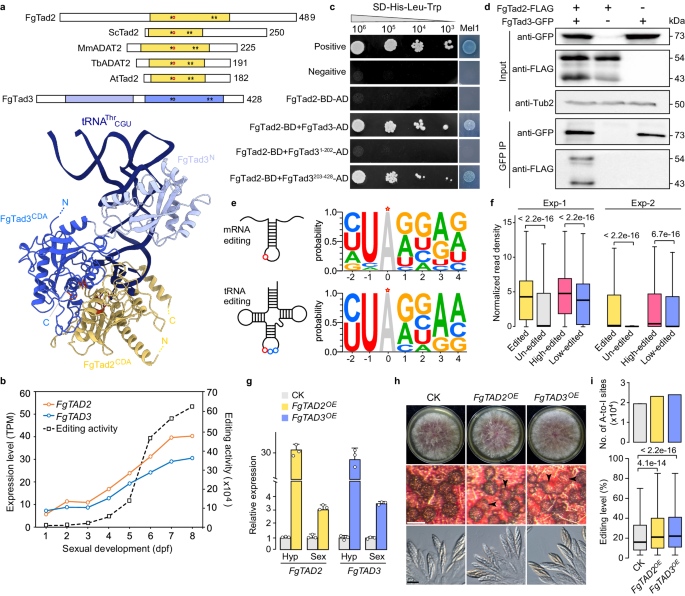
Fig1. FgTad2-FgTad3复合物对tRNA和mRNA的A-to-I编辑
2. FgTad2-FgTad3优先结合被高度编辑的基因的mRNA
禾谷镰刀菌中FgTAD2和FgTAD3的表达水平与性发育过程中的A-to-I mRNA编辑活性正相关(Fig.1b)。通过分析,发现所有鉴定的A-to-I mRNA编辑位点和tRNA的A34位点具有相似的碱基偏好性(Fig.1e)。利用RIP-seq技术,证明了FgTad2-FgTad3复合体倾向于结合被高度编辑的基因的mRNA(Fig.1f),暗示其在真菌的mRNA编辑中发挥作用。
3. 过表达FgTAD2 或 FgTAD3 会促进mRNA编辑
为验证FgTad2-FgTad3在mRNA编辑中的作用,研究者通过RP27启动子在特定位点插入额外拷贝,实现了FgTAD2和FgTAD3的过表达。qRT-PCR分析显示,过表达转化体在菌丝体和子囊壳中的表达水平分别增加了20倍和3倍以上(Fig.1g)。尽管子囊壳外观透明,但转化子仍能正常生长并产生子囊和子囊孢子(Fig.1h)。链特异性RNA-seq测序分析表明,两种转化子有性发育7天的子囊壳中A-to-I mRNA编辑位点的数量和中位数编辑水平都有显著的增加(Fig.1i)。这些结果表明,过表达FgTAD2或FgTAD3可以增强性生殖过程中的mRNA编辑。
4. FgTAD2 和 FgTAD3 均在有性阶段特异性表达一种S-转录本
FgTAD2和FgTAD3均通过选择性转录起始产生两种主要的转录异构体,长转录本(L)在整个过程中组成型表达,而短转录本(S)只在有性阶段表达,且S-转录本在有性发育后期成为主导,可能作为mRNA编辑的主要因子(Fig.2a,b)。根据RNA-seq数据,FgTAD2的S-转录本的转录起始位点定位于L-转录本的开放阅读框(ORF)区域内(Fig.2a)。通过5'RACE技术,研究者确定了FgTAD2两种转录异构体的转录起始位点:L-转录本起始位点在起始密码子上游,而S-转录本起始位点在起始密码子下游,且仅在性生殖阶段表达(Fig.2c)。
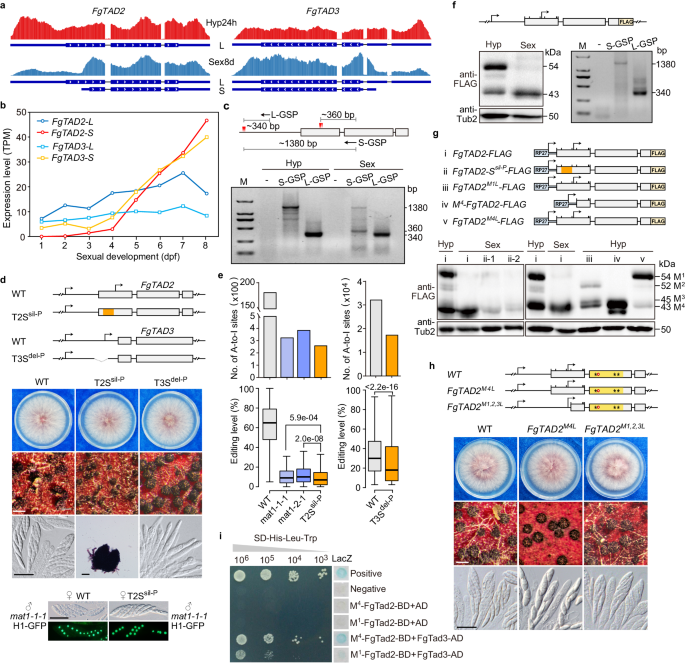
Fig2.FgTad2-FgTad3的转录和翻译调控
5. S-转录本在性发育和mRNA编辑中的作用
研究者通过在FgTAD2的推定启动子区域引入同义突变,生成了T2Ssil-P突变株,该突变株营养生长正常,但产生的子囊壳较小且缺乏子囊和子囊孢子, L-转录本表达不受影响(Fig.2d)。T2Ssil-P突变株与另一突变株交配时,能产生正常的子囊和子囊孢子,说明FgTAD2 S-转录本主要在可育组织中发挥作用。通过链特异性RNA-seq测序分析,发现T2Ssil-P突变株的mRNA编辑位点数量和编辑水平降低,突显了FgTAD2 S-转录本在mRNA编辑中的作用(Fig.2e)。
对于FgTAD3,通过删除其启动子区域,生成了T3Sdel-P突变株,该突变株子孢子形成有缺陷,L-转录本的表达没有明显变化,但在子囊中检测不到S-转录本(Fig.2d)。与野生型相比,T3Sdel-P突变株的mRNA编辑位点数量和编辑水平显著降低(Fig.2e),这表明FgTAD3 S-转录本在mRNA编辑中的重要性。
6. FgTAD2 S-转录本起始密码子与L-转录本的M4密码子对齐
研究者通过在FgTAD2基因上引入同义突变并构建融合蛋白表达系统,发现有性发育7天的子囊壳中主要表达来自FgTAD2 S-转录本的蛋白(Fig.2f,g)。进一步实验表明,S-转录本的起始密码子与L-转录本的第四个阅读框AUG密码子(M4)相对应。在M4密码子引入A-to-C突变后,FgTAD2M4L突变体在子囊孢子形成上出现缺陷(Fig.2h),表明M4对S-转录本的功能至关重要,可能与S-转录本的翻译效率有关。
7. FgTAD2 L-转录本在菌丝中表达两种蛋白质亚型
菌丝体中L-转录本表达产生两种蛋白(约43kDa和54kDa),推测分别由M1和M4密码子启动翻译。通过突变实验,证实了L转录本通过这两种密码子选择性翻译产生不同的蛋白异构体(Fig.2f,g)。M1-FgTad2蛋白比M4-FgTad2蛋白多一个112个氨基酸的N端延伸,但该延伸对菌丝生长和性生殖并非必需(Fig.2h)。酵母双杂交实验表明,M4-FgTad2与FgTad3的结合能力更强,暗示M1-FgTad2的N端延伸可能削弱FgTad2-FgTad3复合体的相互作用(Fig.2i)。
8. 有性阶段特异性蛋白翻译后修饰对于FgTad3功能至关重要
FgTAD3的两种转录异构体共享相同的ORF,只有5'非翻译区(UTR)长度不同。在子囊中表达FgTAD3-GFP融合蛋白时,检测到两个较小的蛋白条带,而非在菌丝中检测到的预期的73kDa蛋白(Fig.3a)。通过质谱分析发现了FgTad3在子囊壳中独有的蛋白乙酰化(acK198)、非典型磷酸化(pE8和pE241)和甲基化修饰位点(meE17)(Fig.3b)。通过突变这些位点生成低修饰突变体发现,K198R和E241A突变体在子囊孢子形成上受损,表明这些修饰对FgTad3功能至关重要(Fig.3c,d)。模拟乙酰化和磷酸化的K198Q和E241D突变体则表现出生长缺陷和子囊壳形成受损(Fig.3c),暗示这些位点如果组成型修饰可能对生长和发育有负面影响。
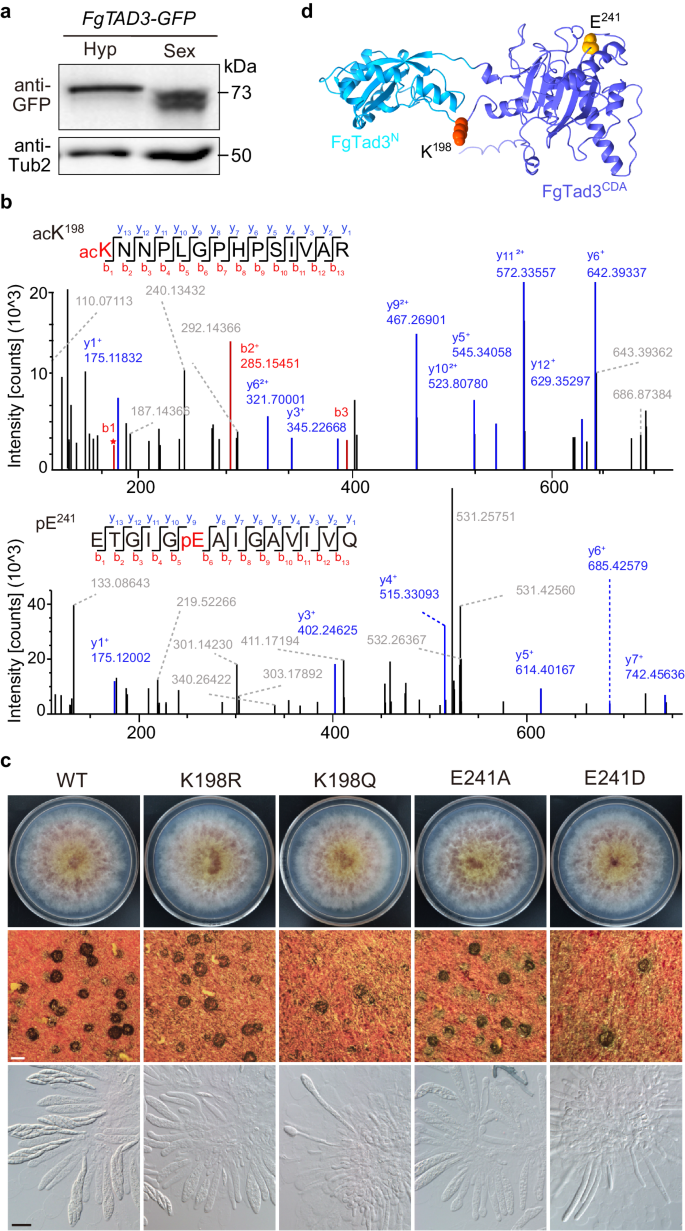
Fig3.FgTad3的翻译后修饰
9. 鉴定A-to-I mRNA编辑的激活因子
在禾谷镰刀菌F. graminearum和粗糙脉孢菌N. crassa中均存在有性阶段特异性的A-to-I mRNA编辑现象,因此,两种真菌应该都存在mRNA编辑的激活因子。研究者通过分析已发表的RNA-seq数据,发现了34个在两种真菌有性发育期特异性表达的同源基因(SSC基因)。其中26个SSC基因的缺失突变株中,有6个表现有性发育缺陷。特别是ssc23和ssc20突变株,分别在有性发育早期阶段显示出无法产生子囊和形成较小子囊壳的缺陷(Fig.4a-c)。对60小时有性组织的RNA-seq分析显示, ssc23突变株中仍然存在A-to-I mRNA编辑位点,而ssc20突变株中不能检测到可靠的编辑位点,表明SSC20对A-to-I mRNA编辑至关重要,因此被重新命名为AME1(mRNA编辑的激活因子)。
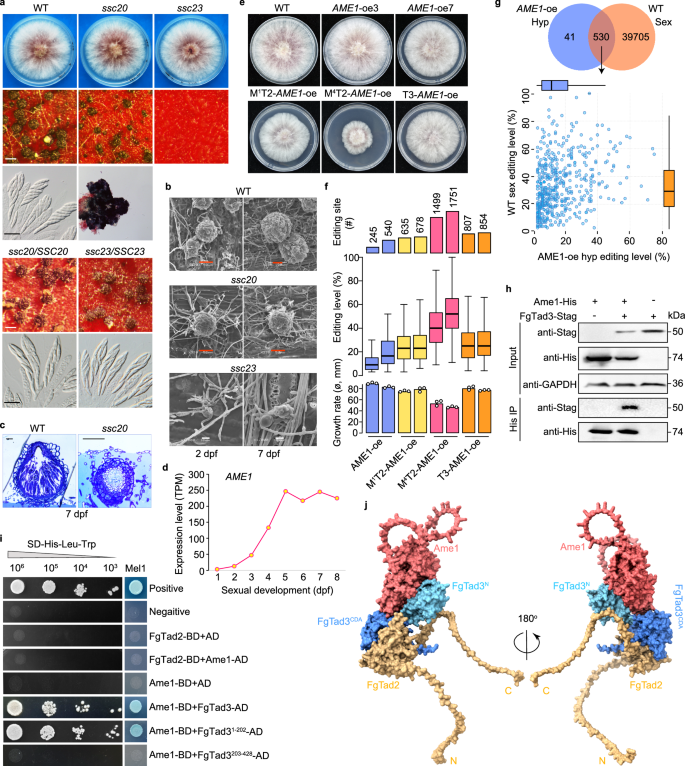
Fig4.有性阶段特异性的Ame1对于有性生殖和A-to-I mRNA编辑至关重要
10. 在菌丝中人工诱导表达AME1后产生广泛的A-to-I mRNA编辑
在有性发育过程中,AME1表达随时间增加并在5天后趋于稳定,与mRNA编辑活性的趋势相吻合(Fig.4d)。通过将AME1的天然启动子替换为RP27启动子,产生的AME1组成型表达转化子在菌落生长上呈现多样性,有的生长正常,有的略慢(Fig.4e)。在这些转化子的菌丝体中检测到大量A-to-I mRNA编辑位点(Fig.4f),证实了AME1在mRNA编辑中的重要作用。超过90%的编辑位点与子囊壳中自然发生的编辑位点重合(Fig.4g),但编辑水平较低。进一步的过表达实验显示,所有转化子都表现出生长缺陷,尤其是M4T2-AME1-oe转化子,其生长速率降低了约50%(Fig.4e)。不同转化子中检测到的编辑位点的数量与其生长缺陷的严重程度相关(Fig.4f),表明mRNA编辑在营养生长期间有害。结果还显示M4-FgTad2具有更高的编辑活性,并与FgTad3有更强的相互作用。
11. Ame1与FgTad3的N端域相互作用
研究者通过免疫共沉淀实验(co-IP)探讨Ame1与FgTad2-FgTad3的关系,发现共表达这两种蛋白对生长有害。在大肠杆菌中确认了Ame1与FgTad2-FgTad3复合体的体内关联(Fig.4h)。酵母双杂交实验表明Ame1仅与FgTad3直接相互作用,并且只与FgTad3的N端域互作(Fig.4i)。通过AlphaFold-Multimer预测,揭示了M4-FgTad2、FgTad3和Ame1形成的三元复合体结构,其中Ame1与FgTad3的N端域结合,可能影响复合体的底物识别和特异性(Fig.4j)。
12. Ame1及其关键残基在mRNA编辑中的演化
Ame1是一种含有DUF726结构域的蛋白质,属于α/β水解酶超家族,具有催化三联体S-D-H(Fig.5a)。研究表明,Ame1的D589残基对其与FgTad3的相互作用至关重要,但催化三联体并非其mRNA编辑活性所必需(Fig.5b,c)。Ame1同源基因在子囊菌中广泛分布,但在酵母菌中已丢失(Fig.5a)。有意思的是,Ame1同源基因在Sordariomycetes真菌祖先上发生了基因负责,其中一个拷贝在Sordariomycetes中经历了加速进化,进化成了具有mRNA编辑活性的Ame1(Fig.5d)。通过同源物替换实验,发现AME1不能被来自Leotiomycetes的SsAME1替代,且SsAme1与FgTad3和SsTad3不发生相互作用(Fig.5c)。此外,三个氨基酸位点(Fig.5e)特别是(MV500-501)对Ame1在Sordariomycetes中的功能创新至关重要(Fig.5d,f),这些位点的变异对Ame1与FgTad3的相互作用和mRNA编辑功能有重要影响。研究人员发现了三个氨基酸位点(Fig.5e),这些位点可能对Sordariomycetes中Ame1的新功能产生至关重要。通过突变实验,发现MV500-501位点对Ame1的mRNA编辑功能至关重要(Fig.5d)。结构模型和酵母双杂交实验表明,这些位点位于Ame1与FgTad3相互作用的界面上(Fig.5f)。在Sordariomycetes中,KN到MV的替换是Ame1进化出与Tad3相互作用和mRNA编辑能力的关键。
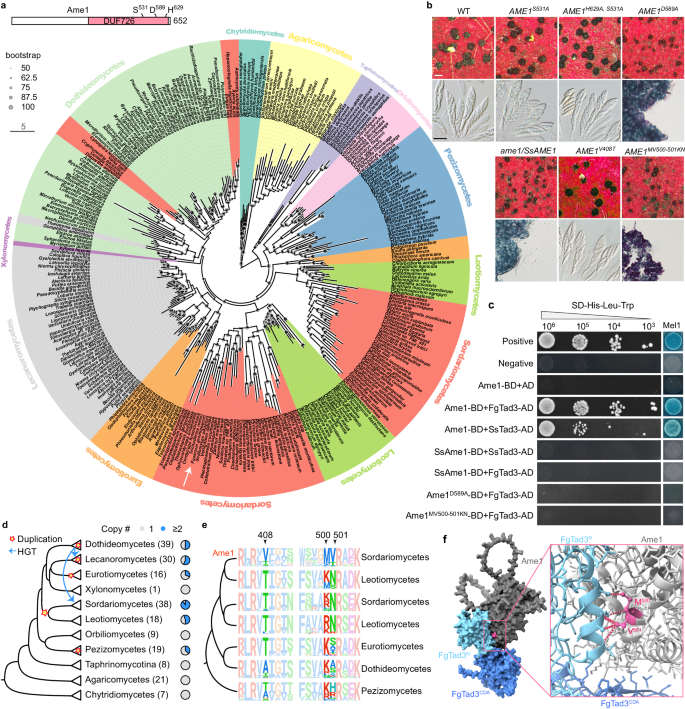
Fig5.Ame1在丝孢菌门中的进化起源及其关键氨基酸残基对mRNA编辑的作用
13. FgTad3中的氨基酸残基对于mRNA编辑至关重要
研究人员利用RIP(重复序列诱导的点突变)机制在FgTad3基因中诱发突变,以确定对mRNA编辑至关重要的氨基酸残基。通过反向插入FgTAD3编码区产生重复序列,诱发突变而不影响原基因转录(Fig.6a)。结果显示,尽管转基因体生长正常,有性生殖却出现了异常子囊孢子(Fig.6b)。350个子囊孢子后代中,有33个生长正常但有性发育有缺陷,其中M120I突变最为常见,影响了9个子代。FgTAD3M120I突变体形成小子囊壳,里面没有子囊和子囊孢子(Fig.6b),且在7天成熟的子囊壳中仅鉴定出40个A-to-I mRNA编辑位点,证实了M120在mRNA编辑中的关键作用。此外,V82I突变可能对M120I突变有补偿作用。酵母双杂交实验表明,FgTad3的某些突变体减弱了与Ame1的相互作用(Fig.6c),而E546和R548两个位点在Ame1与FgTad3的相互作用中起着关键作用(Fig.6e)。
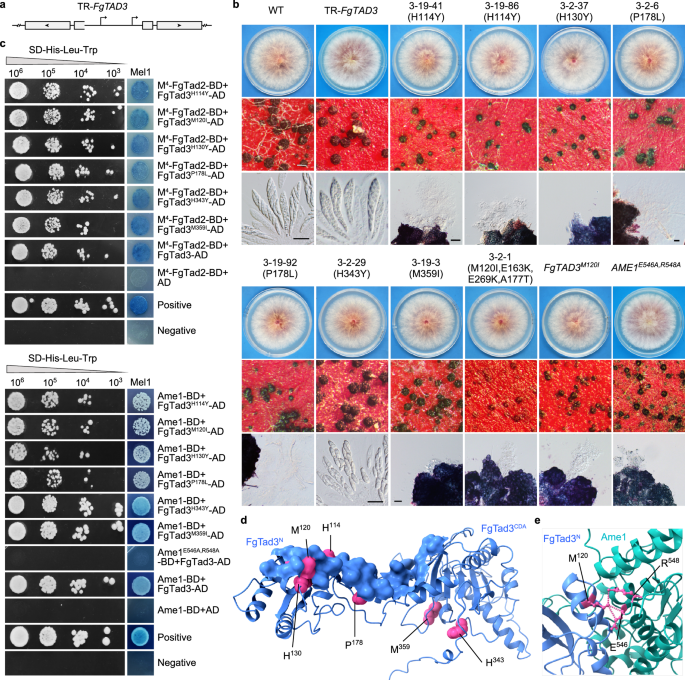
Fig6.FgTad3中对于有性生殖和与Ame1的相互作用至关重要的氨基酸残基
14. 该复合物可编辑酵母、细菌和人源细胞中的mRNA
研究人员在异源系统中测试了FgTad2-FgTad3-Ame1复合体的mRNA编辑活性(Fig.7a)。在酿酒酵母和人源细胞中,该复合体能高效地在目标位点进行A-to-I编辑,且FgTAD2中的E121位点对编辑至关重要。此外,该复合体在大肠杆菌中也显示出编辑活性。通过DNA-seq和RNA-seq分析,研究人员在两种转化株中鉴定出大量A-to-I编辑位点(Fig.7b),其中细菌BL21菌株中的编辑水平显著高于酵母菌株(Fig.7c)。与之前在禾谷镰刀菌F. graminearum中的报道一致,在INVSc1和BL21中观察到相似的核苷酸偏好和编辑位点在发夹环结构中的富集(Fig.7d,e)。这些结果表明FgTad2-FgTad3-Ame1复合体在不同细胞中具有广泛的A-to-I mRNA编辑活性。
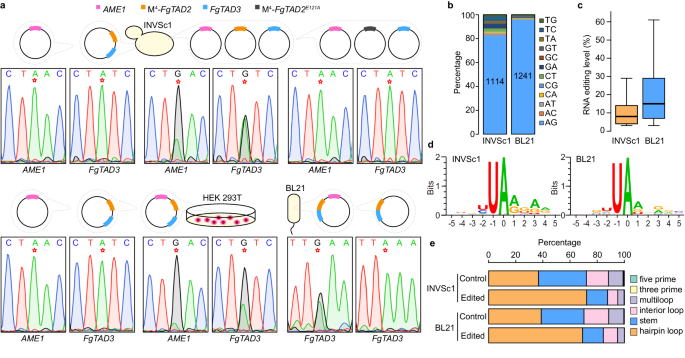
Fig7.异源系统中FgTad2-FgTad3-Ame1复合物的 A-to-I mRNA编辑活性
总 结
本文展示了真菌中进化产生的Ame1使tRNA特异的腺苷脱氨酶FgTad2-FgTad3能够进行A-to-I mRNA编辑。研究表明,FgTad2-FgTad3-Ame1复合物能够高效地编辑酵母、细菌和人源细胞的mRNA,这对于基因编辑技术在治疗和农业中的发展具有重要意义。
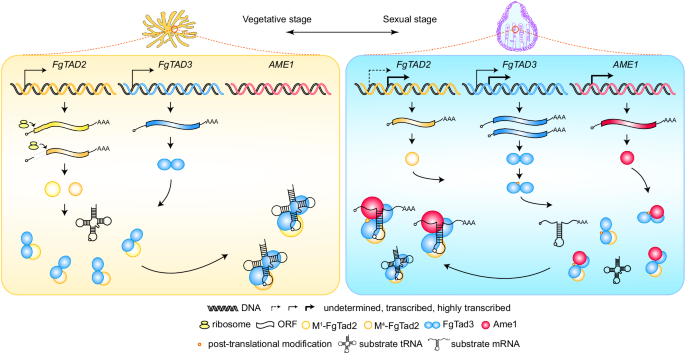
Fig8.A-to-I mRNA编辑机制模型















 1001
1001

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









