







目录
以下内容是关于文献《The Human Transcription Factors》的笔记。
这篇综述讲述了如何鉴定TF与如何对TF(Transcription Factors)的功能进行表征(characterize)。
首先对文献的每一节进行一下梳理:
- How Transcription Factors Are Identified(如何鉴定转录因子)
- Determining TF DNA-Binding Motifs(确定TF的DNA结合motif)
- TF Cooperativity and Interactions with Nucleosomes(TF的协调性和TF与核小体的互作)
- TF Effector Functions(TF效应子功能)
- The Human TF Repertoire(人类转录因子库或集合)
- Sequence Specificities of the Human TFs(人类TFs的序列特异性)
- Conservation and Evolution of Human TFs(人类TFs的保守和进化)
- Expression of Human TFs across Tissues and Cell Types(人类TFs在组织和细胞类型中的表达)
- Human TFs in Genetics and Disease(人类遗传和疾病的TFs)
- Perspective: Learning to Read the Genome(视角:学习读懂基因组)
下面对每一节的内容做一下笔记记录。
一、How Transcription Factors Are Identified(如何鉴定转录因子)
- 真核生物的主要TF家族最初于1980s被鉴定,这些主要TF有:C2H2-zinc finger, Homeodomain, basic helix-loop-helix (bHLH), basic leucine zipper (bZIP), and nuclear hormone receptor (NHR)。
- 这些TF的结合位点通常是由DNase印迹或mobility shift等方法来鉴定的。新的DNA结合蛋白继续通过实验方法被鉴定,这些实验方法包括one-hybrid assays,DNA affinity purification-mass spectrometry,protein microarrays。
- 现如今,大部分已知和推定的TFs是通过与先前确定的DNA结合域(DBD)的序列同源性来鉴定的,这种方法也被用来鉴定TF。
- 当仅基于与DBD的同源性匹配推断TF的功能时,必须小心,因为并非所有这些域都必定结合特异的DNA序列。
二、Determining TF DNA-Binding Motifs(确定TF的DNA结合motif)
本节主要介绍了表征TF结合位点的motif与鉴定TF motif的方法。
1. Motif与PWM及其PWM的限制
motif通常展示为序列logo的形式,如下图就是转录因子AP1的motif:
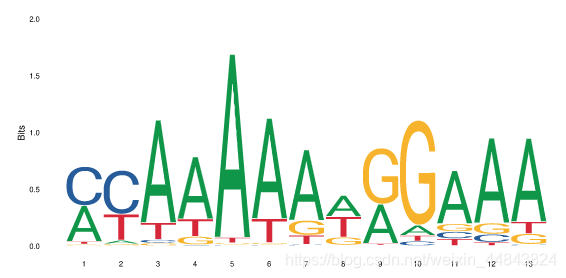
这种序列logo通常用PWM(position weight matrix,位置权重矩阵)来表示。大多数情况下,这些logo可以反映该TF对一些DNA序列的强偏好性,即更偏好与该序列结合。
当然,也有一些PWM不能捕获所导致的complications(并发症),主要原因有:
- motif的各个位置的碱基之间存在依赖;
- 该TF有多个结合模式,例如蛋白质的不同物理构型导致分离的不同基序;
- cooperative interactions(互作)可能会影响TF的结合位点;
- DNA甲基化可以影响结合;
为了解决这些问题,一些更复杂的模型也相继被开发出来。然而,在多数情况下,这些复杂模型相比于PWM的提升很小或几乎检测不到,因此PWM仍然是用于TF结合分析的最普遍使用的模型。
2. TF的序列偏好和结合位点
TF的序列偏好和结合位点可以通过多种体外和体内技术进行评估,表1列出了最流行的方法与该方法的特点。作为相对结合亲和力的预测因子,大量序列的motif可以从定量亲和力测量中最准确地获得,最好是使用纯化的蛋白质和DNA。
3. ChIP-seq在检测TF结合位点方面的作用与限制
这一部分主要讲了ChIP-seq作为一种半定量的衡量方法,其在检测TF结合位点方面的限制,然后讲了先验知识可以帮助TF motif的鉴定,并继续介绍了ChIP-seq的一些限制已经在理论上通过高精度方法(比如ChIP-exo)被解决了,但目前的例子还很少。
ChIP-seq在检测TF结合位点方面的限制主要有4点:
- 结合受染色质状态的影响(许多TF几乎完全结合在开放染色质)以及基因组序列含量的偏差;
- ChIP-seq可以检测到间接结合,这可能导致识别除ChIPped以外的其他蛋白质的motif(ChIPped需要了解一下)
- 由于使用了交联剂,ChIP无法确定平衡结合;
- ChIP数据高度依赖于抗体质量,而许多抗体会交叉反应,并且ChIP-grade抗体不适用于许多TF;
三、TF Cooperativity and Interactions with Nucleosomes(TF的协调性和TF与核小体的互作)
1. 引言
多细胞动物的TFs必须在DNA结合和效应子功能上共同工作以获得所需的特异性(specificity);
已知的TF进行合作的方式有很多种,包括在结合DNA时互相帮助(cooperative binding)或通过不同的机制影响染色质状态或转录(synergistic regulation);TFs也可以作为同型二聚体(homodimer)(如bZIPs, bHLHs)、三聚体形式(热激因子)或高阶结构结合。TF的相互作用本质上与增强子功能和“逻辑”有关。
该文献接下来主要考虑cooperative binding(后文称为协同结合)是如何获得的,因为它与TF功能密切相关。
2. 协同结合的两种方式
协同结合可以通过几种方法发生,最容易理解的方式是通过PPI(protein-protein interactions)来实现协同结合。当两个(或多个)相互作用的蛋白质以兼容的间距和方向结合DNA时,这种方式可以赋予TFs额外的稳定性。
最近的研究表明,DNA介导的协同结合(另一种协同结合方法)在TF功能中也起着重要的作用。在这种方式下,一个TF的结合会在某种程度上影响DNA的形状,从而促进第二个TF的结合。
3. TF与DNA结合的机制
为了能够与核小体DNA结合,TFs必须与核小体竞争,或与核小体或核小体DNA互作以访问这些位点。TFs天生便可以互相协作以与核小体竞争,此外一些TFs可以发动核小体的置换或至少改变它们的构象,最有可能通过招募ATP依赖的染色质重塑者(chromatin remodelers)和其他TFs来实现。
这些TF的活性也可能取决于它们结合核小体DNA的能力,这可能受核小体上结合位点的旋转定位的影响(如the Yamanaka factors POU5F1, SOX2, KLF4, andMYC)。
另一个有趣的观察结果是,不同的染色质重塑者对特定的DNA序列和/或核小体构象具有偏好(Rippe等,2007),这表明核小体和核小体定位机制都赋予TF作用额外的DNA序列特异性。
四、TF Effector Functions(TF效应子功能)
五、The Human TF Repertoire(人类转录因子库或集合)
六、Sequence Specificities of the Human TFs(人类TFs的序列特异性)
- Conservation and Evolution of Human TFs(人类TFs的保守和进化)
- Expression of Human TFs across Tissues and Cell Types(人类TFs在组织和细胞类型中的表达)
- Human TFs in Genetics and Disease(人类遗传和疾病的TFs)
- Perspective: Learning to Read the Genome(视角:学习读懂基因组)

















 765
765

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









