









 本文分享了一项关于组蛋白H3K18la修饰的研究,揭示了其在标记活性启动子和组织特异性增强子中的作用。研究采用CUT&Tag技术,在小鼠和人组织中检测了H3K18la的全基因组分布。
本文分享了一项关于组蛋白H3K18la修饰的研究,揭示了其在标记活性启动子和组织特异性增强子中的作用。研究采用CUT&Tag技术,在小鼠和人组织中检测了H3K18la的全基因组分布。
此前,在推文《组蛋白ChIP-seq研究思路及应用案例分享》中我们分享了组蛋白ChIP-seq的一些研究思路。文章中提到了几种新型组蛋白修饰:丙酰化、丁酰化、琥珀酰化、丙二酰化、戊二酰化、巴豆酰化和乳酸化等,其中组蛋白乳酸化修饰是2019年新发现的,一经发表就登上了Nature[1],该研究将细胞代谢与表观遗传调控联系起来了。
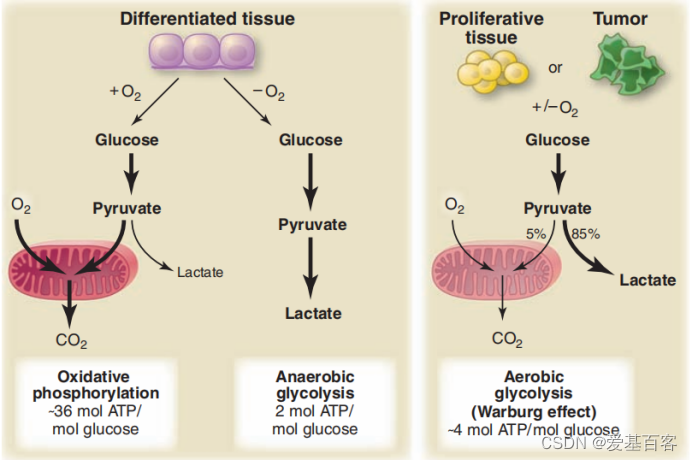
Warburg 效应|Matthew G. Vander Heiden et al. Science. 2009
提到乳酸,就不得不提Warburg 效应(有氧糖酵解),它是癌症的标志之一。在有氧条件下,癌细胞倾向于将葡萄糖转化成乳酸来产生能量,与正常细胞相比,葡萄糖代谢的乳酸更多。因此,肿瘤中的组蛋白乳酸化很可能是异常的。另外,组织中微环境的乳酸水平升高是慢性炎症的一个众所周知的特征。因此,乳酸既是能量底物,又是信号分子。
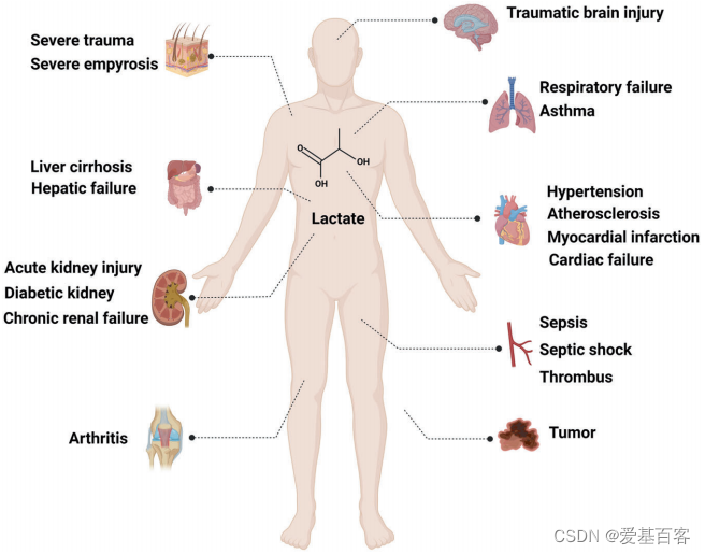
Li, X., Yang, Y., Zhang, B. et al.Lactate metabolism in human health and disease. Sig Transduct Target Ther 7, 372 (2022).
乳酸参与调控肿瘤、心血管系统、呼吸系统、消化系统、泌尿系统等疾病,乳酸在疾病的临床诊断和预后中起着重要的作用[2]。组蛋白乳酸化修饰发现时间短,乳酸又参与调控不少生物学功能。所以,组蛋白乳酸化修饰可以说处在高分文章的红利期,同时它也是今年的国自然热点。
研究组蛋白乳酸化修饰可以考虑ChIP-seq和CUT&Tag技术,爱基百客在这两个产品上都有丰富的项目经验,尤其是王牌产品ChIP-seq(常规修饰甲基化、乙酰化及非常规修饰巴豆酰化等,可以关注往期的项目文章解读)。
本期和大家分享一篇研究组蛋白乳酸化H3K18la修饰的文献,帮大家了解下组蛋白乳酸化修饰的研究思路。

发表期刊:Genome Biol
影响因子:17.906
研究技术:CUT&Tag
2022年10月瑞士苏黎世联邦理工学院营养和代谢表观遗传学实验室团队在Genome Biology(IF: 17.906)期刊发表题为“H3K18 lactylation marks tissue-specifc active enhancers”的研究论文。该研究在小鼠和人的多个组织或细胞中检测了H3K18la全基因组分布,并将其与其他研究的较为成熟的组蛋白修饰进行比较分析,结果发现H3K18la不仅可以标记活性启动子,还可以标记组织特异的活性增强子。
研究背景
在人类、小鼠、植物、真菌和寄生虫样本中已经报道了超过30个组蛋白乳酸化位点。H3组蛋白18位的赖氨酸残基的乳酸化已经进行了一些深入研究,研究显示其在基因启动子上高度富集,并与癌细胞和巨噬细胞中的相关基因的活跃表达有关。但迄今为止,大多数的Kla的报道集中在总Kla水平的变化,但全基因组H3K18la分布及其与其他组蛋白修饰和基因表达的关系报道较少。
该研究发现H3K18la存在于人和小鼠大范围不同代谢状态的细胞类型和组织中。此外,H3K18la不仅富集在启动子上,还富集于组织特异的活性增强子上;并且类似于但不是拷贝H3K27ac修饰的基因组定位。
研究方法
CUT&Tag(爱基百客可提供)
研究思路
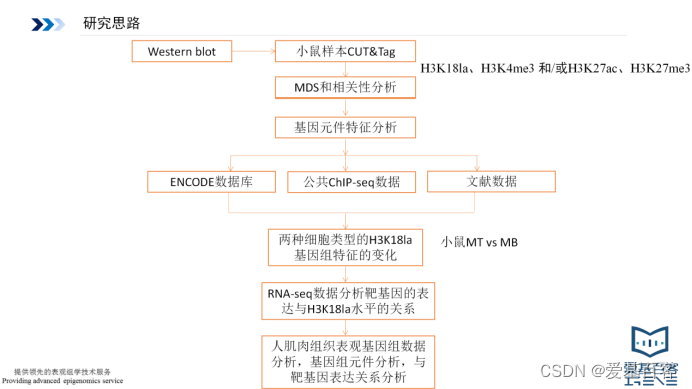
研究结果
1. 组蛋白乳酸化存在于多种代谢状态的组织中
为了探索H3K18la的作用,作者调查了一系列体内和体外样本的H3K18la全基因组分布。因为组蛋白乳酸化与糖酵解活性和乳酸水平有关,作者选择了发育阶段和有丝分裂活性不同的样品,并且这些样品的代谢范围很广,细胞内乳酸水平不同。
所选择的细胞和组织有:mESC-ser(Primed小鼠胚胎干细胞)、mESC-2i(Naïve小鼠胚胎干细胞)、MB(成肌细胞)、MT(肌管细胞)、GAS(小鼠肌肉样本,腓肠肌)、ADIPO(脂肪组织)、BMDM(骨髓源性巨噬细胞)和PIM(缺血后巨噬细胞)。Western blotting结果显示H3K18乳酸化存在所有细胞和组织中(图1A)。
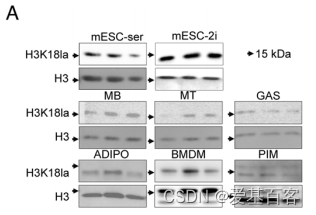
图1A 各个样本的Western blotting结果
2. H3K18la标记活性启动子
为了研究H3K18a的功能,作者构建了H3K18la、H3K4me3和/或H3K27ac(活跃翻译后修饰标记)和H3K27me3(抑制翻译后修饰标记)的CUT&Tag测序文库,用来表征它们的基因组定位。首先,作者在全局范围比较了这些数据集。采用了多维尺度分析(MDS分析),基于标记类型(活跃vs抑制),根据样品来源以及特异的翻译后修饰,这些样品聚集成簇(图1B)。MB、MT和GAS活性标记以及BMDM和PIM数据集聚集在一起(图1B)。对于每种细胞类型,H3K18la样本聚集得更接近H3K27ac,而不是H3K4me3样本。这强调了H3K18la的基因组分布具有组织特异性,类似于来自同一组织类型的H3K27ac。值得注意的是,H3K18la ADIPO样本聚集在接近肌肉样本,尽管它们有明显的代谢率差异。尽管mESC-2i和mESC-ser或MB和MT之间的代谢状态存在差异,但它们的H3K18la图谱也基于其起源聚集在一起。这些结果表明,发育特性对H3K18la基因组分布具有重要意义。
另外,作者使用“SEACR peak caller”来定义组蛋白翻译后修饰(hPTMs)。定量H3K18la peaks的相关性分析结果表明mESC H3K18la特征与分化的细胞类型差异最大(图1C)。与赵英明课题组此前报道的结果一致,作者发现许多H3K18la峰定位于转录起始位点(TSS)附近,并与基因启动子或内含子重叠(图1D)。mESC和ADIPO的H3K18a 峰落在启动子区域的占比最高(~40%)(图1D)。ADIPO、GAS、PIM、MB和MT的H3K18la峰分布在TSS下游略有偏移,相应的H3K4me3/H3K27ac活性标记也是如此,但(抑制性)H3K27me3峰则没有。
基因组特征尺寸校正富集分析显示H3K18la信号在CpG岛(CGI)启动子中明显富集,而在非CGI启动子中没有富集(图1E)。为了研究启动子能否被不同组合的活性组蛋白翻译后修饰标记,作者将H3K4me3、H3K27ac和/或H3K18la 峰标记的启动子重叠(图1F)。作者发现,这些“活性”启动子中的绝大多数要么由H3K4me3+H3K27ac+H3K18la标记(37-50%),要么由H3K4me3+H3K27ac标记(19-21%),要么由H3K4me3单独标记(15-24%)。因此,只有大约一半的H3K4me3标记的启动子也被H3K18la标记。
为了研究H3K18la标记的活性启动子与非H3K18la标记的活性启动子之间是否存在功能差异,作者还观察了基因表达、基因本体(GO)富集及其相关基因的TF结合位点富集。综合这些结果,作者发现H3K18la富集于一个活性基因启动子(主要是CGI)的子集中,而H3K18la启动子水平与基因表达和公认的活性标记H3K4me3和H3K27ac呈正相关。
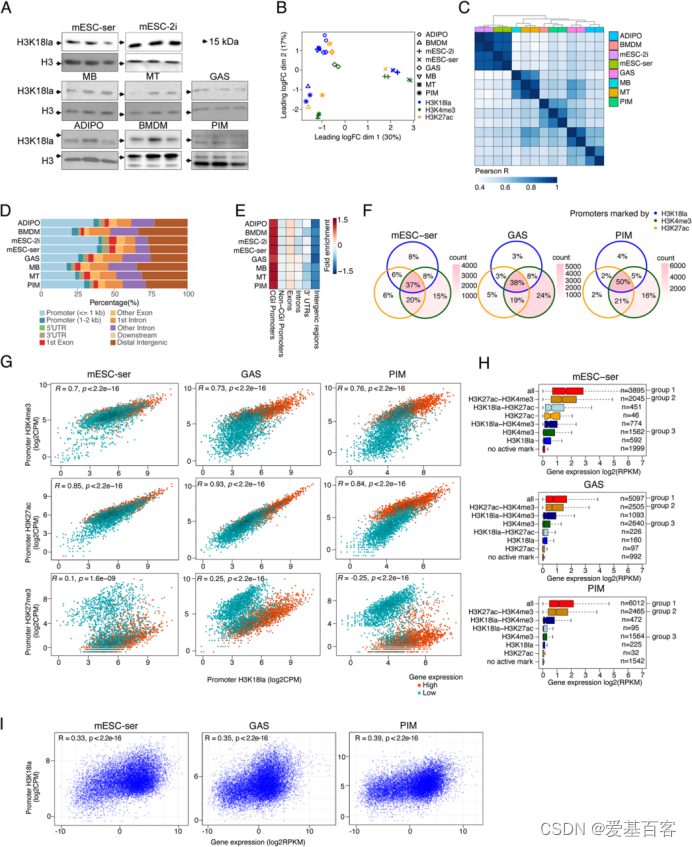
图1 H3K18la标记活性的CGI启动子
3. H3K18la标记活性启动子
在上面的研究中,作者发现H3K18la 峰在TSS附近明显富集,但是也有一部分落到了TSS的远端(>2kb),并且它与启动子不重叠,而是定位于内含子和基因间区域。此外,与其他组蛋白修饰的全基因组定位比较分析中发现H3K18la与H3K27ac相似,而H3K27ac既可以标记启动子,也标记增强子。于是,作者他们推测H3K18la是否能标记增强子。
该研究主要从三个方面去验证这一推测。第一,构建了无监督的 ChromHMM分析,然后计算了ChromHMM状态富集在数据库ENCODE的情况。第二,将公共ChIP-seq数据集与该研究的数据集进行比较分析,发现CUT&Tag的数据与它们匹配良好。H3K18la数据与公共组织特异的ChIP-seq数据重叠支持了H3K18la标记组织特异的活性增强子。第三,作者分析了文献报道的公认增强子信息,计算出这些增强子中的哪一部分与H3K18la峰重叠。在7个被调查的组织中,有5个(未被发表的MB和ADIPO增强子),超过60%的已发表的组织特异性增强子被研究的组织对应的H3K18la峰覆盖。
另外,作者发现大量假定的dELS(远端增强子)仅由H3K18la峰标记,而没有由H3K27ac峰(或H3K4me3)标记,这表明在dELS中H3K18la还有额外的特异性作用。随后,作者分析H3K18la 标记的增强子是否与靶基因高表达有关。相关性分析发现远端增强子上H3K18la水平和靶基因表达的相关性弱,但正相关且显著的。
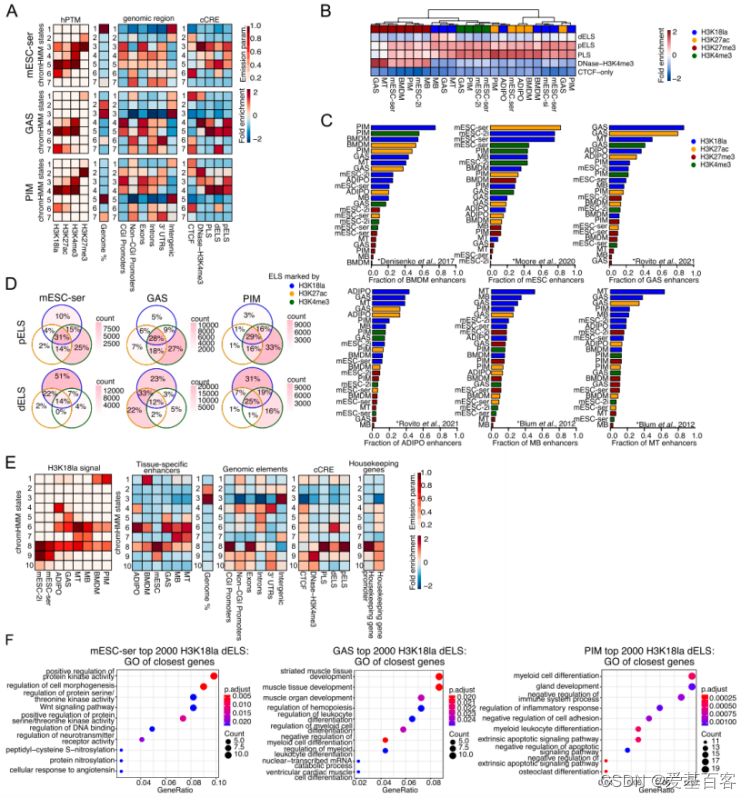
图2 H3K18la标记组织特异的活性增强子
4. H3K18la的动态变化影响转录适应
然后,作者重点研究了密切相关的细胞类型之间H3K18la的功能相关变化,并比较了MT与MB和mESC-ser与mESC-2i。作者观察了MT和MB两种细胞基因组特征的H3K18la的变化。两种细胞状态间,很多启动子失去或者获得H3K18la的修饰;这些变化与相关基因的上下调呈正相关。以Myh1基因为例,Myh1基因的表达在MT中通过Myhas的激活和高乳酸化来调控。
在增强子方面,与作者之前的观察一致,增强子(dELS)乳酸化比任何其他基因组区域的H3K18la的变化更动态。而dELS中H3K18la的变化也与最近基因的基因表达的变化呈正相关。对MT vs MB或mESC-ser vs mESC-2i中,H3K18la变化显著的最接近dELS的基因进行GO分析, 结果发现GO term与各自的细胞类型有关。对MB- 和MT-特异增强子的进一步细节分析,支持作者的推测:启动子和增强子的定量乳酸化变化可以再现,甚至可能促进细胞状态的转变。总的来说,作者的数据表明,乳酸可能是一种细胞状态的过渡,通过基因启动子和增强子的高乳酸化来刺激代谢产物。
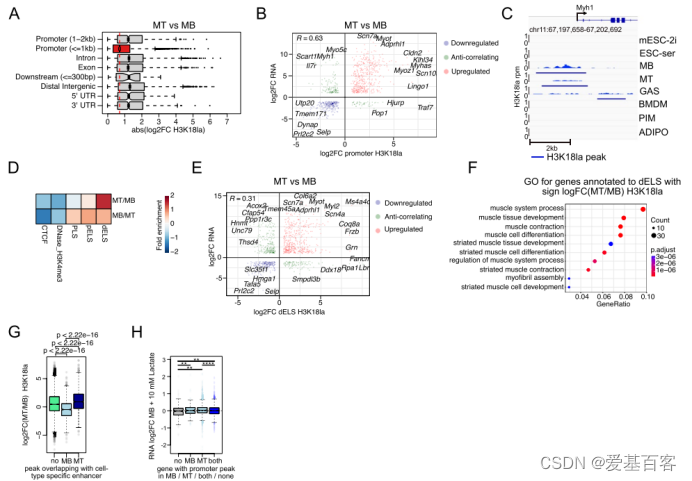
图3 H3K18la影响转录适应的动态变化
5. H3K18la在人肌肉中是保守的,且标记活性启动子和增强子
作者为了确认他们的发现是否在人组织中保守,分析了两个人受试者股外侧肌表观基因组,并评估了H3K18la、H3K27ac、H3K4me3、H3K27me3和H3K9me3。根据MDS分析,H3K18la的特征与H3K27ac和H3K4me3的相关性最好(图4A)。H3K18la的peaks在基因组特征的分布与小鼠肌肉样本的结果非常相似,H3K18la 在启动子、内含子和基因间区富集(图4B),并且相当一部分peaks位于距离TSS>10kb区域。H3K18la峰在CGI启动子中强烈富集,而在非CGI启动子中则较少富集(图4C)。大部分H3K18la标记的启动子也被H3K27ac和H3K4me3标记(图4D)。作者发现启动子上的H3K18la水平与来自同一肌肉的公共基因表达数据的相关性相当弱(尽管是正相关),尤其在与H3K4me3或H3K27ac比较时。
然后,对那些启动子H3K18la水平最高的前2000个基因进行GO分析,结果显示H3K18la标记了对肌肉生物学重要的基因的启动子,基因启动子上有活性组蛋白标记(H3K27ac和/或H3K4me3)而没有H3K18la的在肌肉生物学项目上没有富集。此外,远端增强子H3K18la水平与基因表达呈正相关,但总体上较弱。
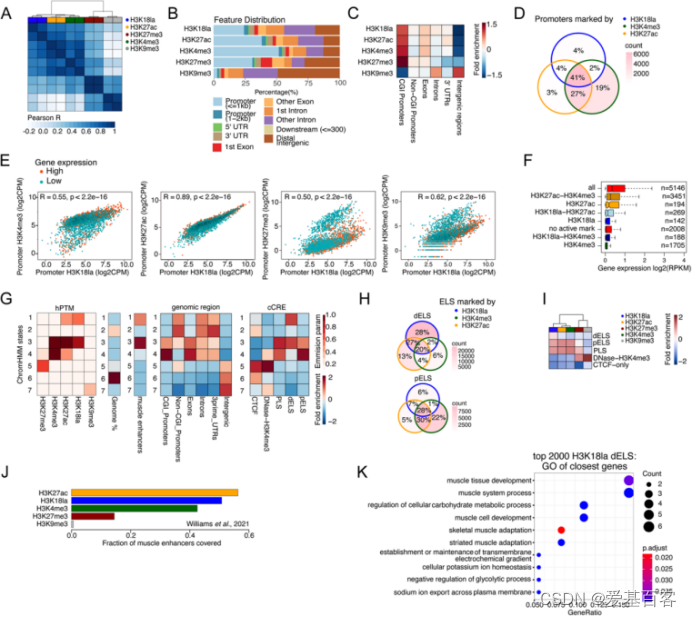
图4人肌肉中的H3K18la标记着活性启动子和增强子
研究结论
该研究生成了小鼠和人组织的H3K18la分布的全基因组数据集,其中这些组织覆盖到许多代谢状态。监督(与公共数据重叠)和无监督(ChromHMM)分析反映H3K18la除了活性启动子,还标记组织特异的活性增强子。研究中还有一个兴奋的发现,活性CGI启动子上的H3Kla可能主要标记启动子嵌入的增强子序列,使得H3K18la成为与H3K27ac部分不同的组蛋白翻译后修饰的增强子标记。
文献链接:doi: 10.1186/s13059-022-02775-y
文献资料
1. Zhang, D., Tang, Z., Huang, H. et al. Metabolic regulation of gene expression by histone lactylation. Nature 574, 575–580 (2019). https://doi.org/10.1038/s41586-019-1678-1
2. Li, X., Yang, Y., Zhang, B. et al.Lactate metabolism in human health and disease. Sig Transduct Target Ther 7, 372 (2022).
◆ CUT&Tag产品介绍 ◆
CUT&Tag是蛋白质和DNA互作新兴的实验方法,该方法是通过蛋白特异性抗体引导Protein A-Tn5酶在目标蛋白结合的DNA位置进行切割并且在序列两端加测序接头,经过PCR扩增后形成可以用于高通量测序的文库,随后对文库进行测序分析从而解析与目标蛋白结合的DNA片段。
有组蛋白乳酸化修饰研究需求的老师可以和我们咨询沟通,爱基百客提供方案设计、样本制备、测序和分析一站式服务哦~

















 312
312

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









