








期刊:Frontiers in Plant Science
影响因子:6.627
发表日期:2022.6.30
单位:省部共建青稞和牦牛种质资源与遗传改良国家重点实验室
关键信息:白粉病(白粉菌),代谢组学,转录组,蛋白质组,茉莉酸,H3K27me3
2022年6月30日西藏自治区农牧科学团队在Frontiers in PlantScience上发表一篇名为“Resistance to Powdery Mildew in Qingke Involves the Accumulation of Aromatic Phenolamides Through Jasmonate-Mediated Activation of Defense-Related Genes”的研究论文。作者通过利用ChIP-seq、RNA-seq和RT-PCR技术,对感病材料(G72)和抗性材料(K69)进行研究,通过整合转录组和蛋白质组以及代谢组数据分析,得出以下结论:茉莉酸信号是通过去除H3K27me3以及激活防御相关基因的表达控制芳香族PAs的积累,进而增强青稞的免疫抗性。爱基百客为该研究提供ChIP-seq的技术支持。

研究背景
大麦(Hordeum vulgare L.)是10000年前在自然选择和人工选择下最早驯化和栽培的作物之一。西藏无壳大麦(Hordeum vulgare L.var.nudum),在汉语中也称为“裸大麦”或“青稞”,是西藏地区的主要主食。它还广泛用作牲畜饲料和生产啤酒、医药和保健产品的原料。目前研究主要集中在鉴定与白粉病相关的抗性基因,但缺少在青稞中对 PM 抗性代谢调控网络的机制的探索研究。因此,全面了解青稞抗白粉病的代谢调控网络,对探索青稞抗白粉病的潜在机制至关重要。
研究思路
从青稞核心种质资源中鉴定出两个栽培青稞品种,一份是抗性材料(K69),另一份是感病材料(G72)。均在西藏农牧科学研究院种植。将2份幼苗(20日龄)接种Blumeria graminis (DC.) f. sp. hordei (Bgh),置于25℃以及光照时间为14小时/10小时的温室里生长,分别在6、36、72和168 hpi下监测了植株的表型和症状。同时收集叶片组织,立即冷冻在液氮中并在−80℃保存,每个处理进行三次生物学重复试验。

研究结果
1. 生理指标的测定
通过对两种青稞幼苗接种Blumeria graminis(DC.) f. sp. hordei (Bgh)后不同生长期的表型观察,结果显示(图1),随着处理时间的增加,叶片的逐渐出现病变,并迅速扩大。其中,G72植株在36hpi时叶片出现小病变,这些病变迅速扩大,并在72 hpi时大量分布在叶片上,最终持续的PM感染导致病斑在168 hpi覆盖整个幼苗(图1A)。而K69植株上的病变受到很大限制,病变直到168 hpi才出现在叶片上 (图1B)。以上结果表明,K69 株系比 G72株系更能抵抗 PM 感染。

图1. G72株系和K69株系接种后的疾病发展过程
2. 茉莉酸信号在白粉病感染早期的诱导作用
前期研究表明,植物激素信号转导主要参与PM感染的早期反应(6和36 hpi)。为了确定哪些植物激素在 PM 处理后的早期植物-病原体相互作用阶段中起重要作用,使用GC-MS检测方法测定了两个青稞感染PM期间的JA-Ile(茉莉酸-异亮氨酸)、JA(茉莉酸)、水杨酸(SA)和脱落酸(ABA)的含量变化。在用PM处理的两个品种之间发现了不同的植物激素积累模式(图2),与G72相比,在K69系中观察到JA-Ile和JA后期显著下降,其值与G72相似或低于G72(图2A,B)。在PM感染后,与G72相比,JA相关基因的相对表达量在K69中明显上调。因此,较长时间的JA-Ile积累可能是G72感染真菌后的延迟反应。
此外,SA和ABA在不同的植物-病原体相互作用中发挥拮抗作用[1],同时也是促进叶片衰老和细胞死亡的关键信号分子[2]。在本研究中,PM感染显著诱导G72中SA和ABA水平(图2C,D)。以上结果表明,青稞对PM胁迫的抗性与JA信号的快速诱导密切相关。因此,诱导的时间可能对激活对这种感染的防御至关重要。
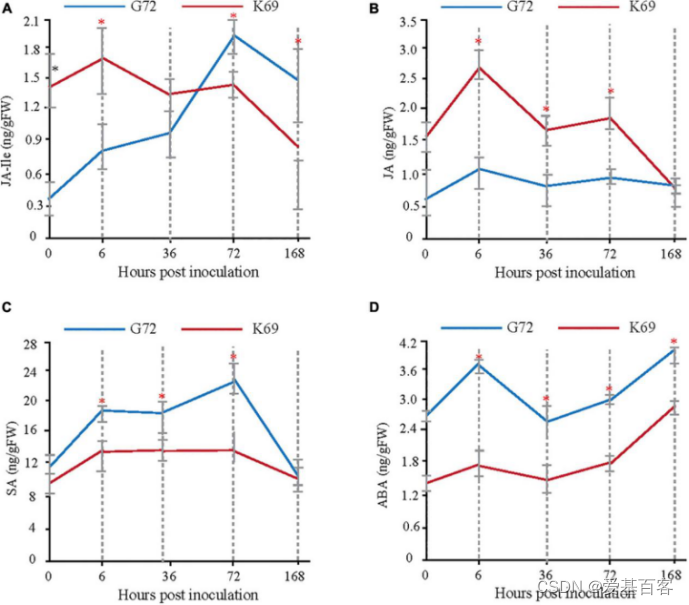
图2. K69与G72系白粉病侵染期间植物激素水平的测定与比较
3. PM诱导产生的芳香族酚酰胺是青稞抗白粉病的主要原因
在本研究中,作者发现与G72相比,在PM感染下K69中多胺(PAs)的前体,包括L-苯丙氨酸、肉桂酸、L-酪氨酸、L-精氨酸和对香豆酸显著降低(36-168 hpi)(图3A-E)。然而,与PAs前体的积累模式相比,在 K69中发现 PAs水平显著升高。其中,脂肪族 PAs表现出结构性积累,而芳香族 PAs则在青稞中对 PM 的抗性表现出诱导积累(图3F)。以上结果表明,青稞可能通过诱导芳香族PAs积累来抵御PM胁迫。

图3. K69与G72系白粉病侵染期间与PAs及其前体相关的表达水平
4. 芳香酚酰胺生物合成的N-酰基转移酶基因
通过转录组分析发现,K69和G72青稞中与PA生物合成相关的基因和代谢物被差异调控,如图所示(图4),与G72相比,与胺、氢肉桂酸CoA和PA生物合成相关的苯丙氨酸解氨酶(PAL)、对香豆酸3羟化酶(C3H)、4-香豆酸:辅酶a连接酶(4CL)、N-羟基肉桂酸/苯甲酰转移酶(HCT)、色胺羟基肉桂酸转移酶(THT)、色胺苯甲酰转移酶(TBT)和腐胺羟基肉桂酸转移酶(PHT)基因在K69中表达上调,并通过qRT-PCR分析验证这9个基因的表达水平,与转录组数据一致。

图4. K69与G72系白粉病侵染期间PAs生物合成途径的差异基因表达和差异代谢物的积累模式
酚酰胺是由BAHD N-酰基转移酶乙酰化的次级代谢产物[3]。为了研究编码BAHD N-酰基转移酶的基因在PM感染过程中与PAs合成有关,作者使用RNA-seq中的FPKM值分析了在两个青稞品种中所有注释的BAHD N-酰基转移酶的表达变化,有趣的是,有9个N-酰基转移酶基因(HvPHT3、HOUSG2299900、HOUSG0308900、HvACT、HvTHT、HOUSG4111500、HvTBT1、HvTBT2和HOUSG3724300)在K69中表达显著增加,然而在G72中没有显著变化(图5A)。系统发育分析显示,HvPHT3、HvACT、HvTHT、HvTBT1和HvTBT2编码的蛋白属于不同的BAHD N-酰基转移酶,已知这些酶在体外催化PAs的N-酰基化,例如腐胺、酪胺和色胺(图5B)[3,4]。综上所述,这5个基因被鉴定为参与PAs生物合成的候选基因。
为了验证这5个候选基因的功能,作者在大肠杆菌BL21菌株中的重组蛋白进行了体外酶活性测定和本氏烟草体内瞬时表达的体外酶促试验,分析发现HvACT和HvPHT3对腐胺、丁胺等脂肪胺具有较强的酰基活性(图5C-F和表1)。相比之下,HvTBT1和HvTBT2主要表现出对芳香胺(如酪胺、色胺和血清素)的强活性,而对脂肪胺(腐胺)的弱活性(表1)。以上这些结果表明,HvTBT1和HvTBT2两个基因参与了青稞芳香族PAs的生物合成。此外,为了确定负责芳香族PAs生物合成的两个N-酰基转移酶基因(HvTBT1和HvTBT2)是否位于青稞分化区(计算两两组之间的固定指数(Fst)),作者首先将这两个基因的序列与此前青稞群体分化研究中使用的青稞参考基因组进行比对,发现HvTBT1和HvTBT2分别位于4号染色体的55.68和473.55 Mb。然后,通过染色体定位与报道的群体分化区域的比对,将这两个基因准确定位在青稞群体分化中[4]。以上结果表明:芳香族聚氨基甲酸乙酯的合成途径是经过自然选择或人工驯化的。

图5.K69与G72系白粉病侵染期间PAs生物合成途径相关基因的功能表征
表1.HvACT、HvPHT3、HvTBT1和HvTBT的动力学参数。

5. 茉莉酸甲酯诱导的去甲基化及促进酚酰胺生物合成
为了研究青稞中PA生物合成途径的调控,作者利用13种PAs作为诱饵,根据皮尔逊相关系数(PCC)筛选数据高度相关的基因和蛋白质。根据组学相关数据表明,7个与PAs相关基因(JMJ705-like和6个生物合成基因)和1个蛋白(茉莉酸盐诱导蛋白)与这些PAs密切相关(PCC>0.6)。此外,通过计算上述鉴定出的基因/代谢物/蛋白的PCC值,构建了相关网络(图6A)。有趣的是,共表达网络分析发现了JMJ705-like蛋白,据报道该蛋白参与了茉莉酸甲酯诱导的去除H3K27me3和基因表达的激活[5],茉莉酸酯诱导的蛋白与青稞的PA生物合成呈正相关(图6A)。此外,在PM处理后,JMJ705-like蛋白和茉莉酸诱导的蛋白显示出与4个N-酰基转移酶基因非常相似的表达模式(图5A)。这些结果表明了茉莉酸介导的JMJ705可能是通过激活4个负责PA合成的N-酰基转移酶基因参与PA的生物合成。
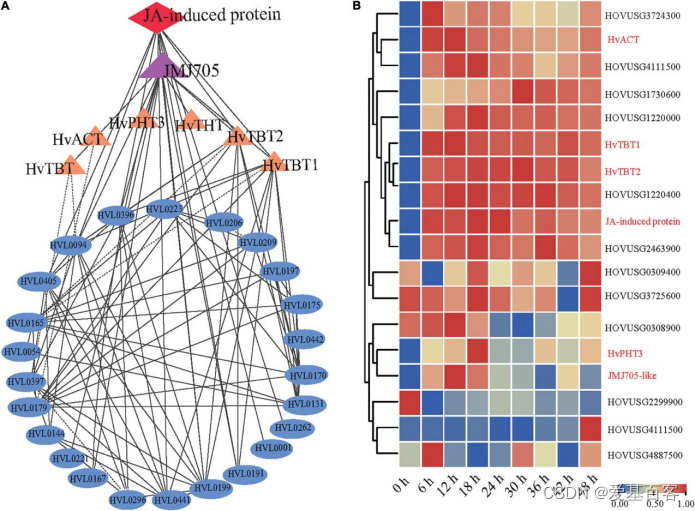
图6. JMJ705-like和JA诱导的蛋白与PAs的生物合成相关
为了进一步研究PAs积累对PM侵染反应的调节,作者使用MeJA处理了K69品种15d幼苗的0、6、12、18、24、30、36和48h。qRT-PCR分析表明,主要参与芳香PA生物合成的HvTBT1和HvTBT2的相对表达在MeJA处理期间,增加了约70.1-103.8倍(图6B)。同时,主要参与脂肪族PAs生物合成的HvACT和HvPHT3的表达水平在处理株系中仅比对照株系高2.0-8.0倍(图7A)。以上数据表明,MeJA处理对芳香族PAs控制基因的表达有较大的影响。此外,通过MeJA处理,也明显上调JMJ705的表达水平(图7A)。先前的研究表明,JMJ70-like与组蛋白修饰密切相关,例如在水稻中组蛋白H3的第27个氨基酸上三甲基化(H3k27me3)[6]。紧接着,作者对G72的MeJA、PM和乙醇(模拟)处理进行了ChIP-seq和ChIP-qPCR检测分析。结果表明,在MeJA处理下,以上4个基因的H3K27me3水平均显著降低(图7B,C)。在PM感染下也检测到了类似结果(图7B,C)。

图7. MEJA介导(或PM处理)对4种n-酰基转移酶的调控分析
这些结果表明,与PA生物合成相关基因的表达水平受JMJ705-like介导的组蛋白修饰去除的调控,并且该过程与MeJA信号通路响应PM胁迫密切相关。这一结果表明,JMJ705基因可能参与了MeJA介导的植物免疫抗性,其保守机制可能在禾草科广泛存在(图8)。
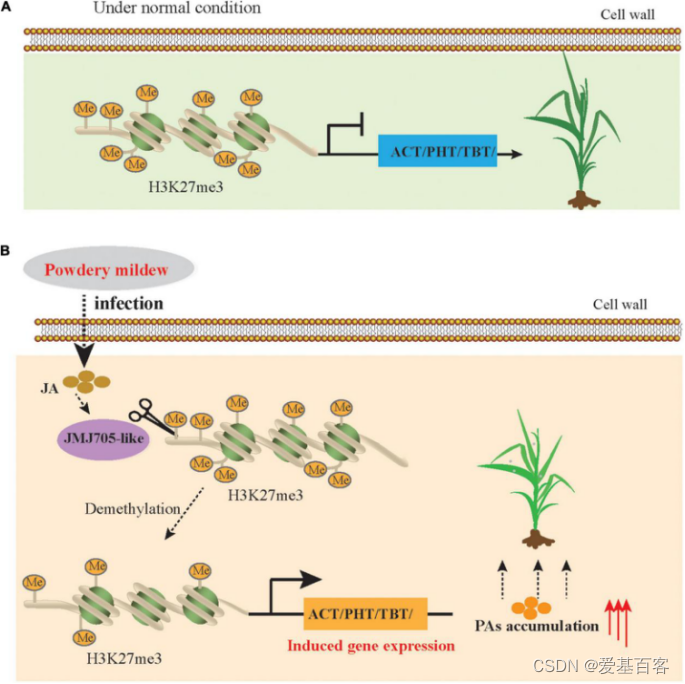
图8. JMJ705-like介导BAHD n-酰基转移酶表达改善白粉病抗性的简化模型
研究结论
在本研究中,作者通过对抗性(K69)和敏感(G72)青稞品种资源进行比较分析,利用转录组、代谢组和蛋白质组数据整合分析,筛选出与青稞抗性相关的潜在基因、蛋白和代谢物。通过数据整合分析,构建了一个共同表达网络,发现MeJA和PM诱导的组蛋白赖氨酸脱甲基酶JMJ705基因与PAs生物合成具有高度相关性。最后通过ChIP-seq和ChIP-qPCR进一步分析表明MeJA信号是通过去除H3K27me3和激活防御相关基因的表达控制芳香族PAs的积累以增强植物的免疫抗性。
这项工作不仅揭示了与芳香族PAs积累相关的PM胁迫抗性的调控机制,而且还提供了一种富含PA的青稞资源。这些发现为促进青稞的分子设计育种和可持续食品奠定了基础。
参考文献:
[1]Moeder W , Ung H , Mosher S , et al. SA-ABA antagonism in defense responses[J]. Plant signaling & behavior, 2010, 5(10):1231-1233.
[2]Zhu H, Chen L, Xing W, et al. Phytohormones-induced senescence efficiently promotes the transport of cadmium from roots into shoots of plants: a novel strategy for strengthening of phytoremediation[J]. Journal of hazardous materials, 2020, 388: 122080.
[3]Beck M, Zhou J, Faulkner C, et al. Spatio-temporal cellular dynamics of the Arabidopsis flagellin receptor reveal activation status-dependent endosomal sorting[J]. The Plant Cell, 2012, 24(10): 4205-4219.
[4]Zeng X, Yuan H, Dong X, et al. Genome-wide dissection of co-selected UV-B responsive pathways in the UV-B adaptation of qingke[J]. Molecular Plant, 2020, 13(1): 112-127.
[5]Virdi K S, Wamboldt Y, Kundariya H, et al. MSH1 is a plant organellar DNA binding and thylakoid protein under precise spatial regulation to alter development[J]. Molecular Plant, 2016, 9(2): 245-260.
[6]Li T, Chen X, Zhong X, et al. Jumonji C domain protein JMJ705-mediated removal of histone H3 lysine 27 trimethylation is involved in defense-related gene activation in rice[J]. The Plant Cell, 2013, 25(11): 4725-4736.















 3217
3217











 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









